| Y. Yogo, corresponding author. e-mail: yogo-tky@umin.ac.jp phone: +81-3-5800-8662; fax: +81-3-5800-8917 Published online 29 June 2004 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase. 03115 |
The Mamanwa (or Mamanua) people of Northeast Mindanao in the Philippines belong to the hunter-gatherer Negritos of Southeast Asia and the western Pacific, and are presumed to be among the oldest indigenous peoples in the region (Omoto, 1984). Direct ancestors of the present-day Mamanwa have been postulated to be either the Proto-Malay population of late Pleistocene Sundaland (Omoto, 1984), or the Negritos from Borneo, Sumatra, and Malaya who came via the still remaining land bridges some 30,000–25,000 years ago (Lagassé, 2001; Burton, 2003). The Indonesians and Malays followed in successive waves, and their descendants now account for the majority of modern Filipinos. As each new wave of migration entered the Philippines, the earlier peoples were either driven into the hinterlands, or assimilated.
More recent history, for instance, describes that for the whole duration of Spanish rule in the 1500s, indigenous populations in the Philippines, including the Mamanwa, endeavored to avoid colonization by resettling in more inaccessible and mountainous regions and practiced slash-and-burn farming along with hunting and foraging (Burton, 2003). In fact, pockets of the Philippine Negritos remain to this day in remote areas mostly along the Sierra Madre mountain range that extends along the entire eastern side of Luzon island in the northern Philippines (Headland, 2002). The Mamanwa is the only Negrito group in the southern Philippines, geographically separated from the other Negrito groups in Luzon and the central Philippines. Presently, there are about 1500 Mamanwas confined to the provinces of Agusan and Surigao in Northeast Mindanao.
To obtain information about the origins of the Mamanwa, this study attempted to elucidate JC virus (JCV) genotypes in a Mamanwa population in the province of Surigao del Norte in Northeast Mindanao, the Philippines. JCV is ubiquitous in the human population (Padgett and Walker, 1973), usually being transmitted from parents to children during cohabitation (Kunitake et al., 1995; Kato et al., 1997; Suzuki et al., 2002; Zheng et al., 2004). All JCV strains in the world constitute a single serotype (Major, 2001), but can be classified into more than ten major genotypes, with each occupying a unique geographical domain (Yogo et al., 2004). Distribution patterns of JCV genotypes have been found to be compatible with human migrations (Yogo et al., 2004). It was recently suggested that like modern humans, JCV originated in Africa (Pavesi, 2003). JCV genotyping analysis has helped gain new insights into the origins of ethnic groups worldwide (Yogo et al., 2004).
We recently investigated the JCV genotype profiles of modern urban Philippine populations, the Tagalogs of Luzon island, Cebuanos of Cebu island, and residents of Pamalican island in Palawan (Sugimoto et al., 1997; Miranda et al., 2003; Takasaka et al., 2004). Stoner and his colleagues reported JCV genotype profiles in Oceanic populations (Ryschkewitsch et al., 2000; Jobes et al., 2001; Yanagihara et al., 2002). Takasaka et al. (2004) elucidated the relationships among JCV genotypes in the Philippines and the western Pacific. Here, we report the JCV genotype profile in a Philippine Negrito population, the Mamanwa. We discuss the present findings in the context of human expansion in the Pacific as well as in relation to current knowledge about the genetics of the Mamanwa and other populations.
Urine samples were collected with informed consent from 49 unrelated Mamanwas, aged 40 years or older, residing in Mat-i, Sison and Surigao city of Surigao del Norte province in Northeast Mindanao, the Philippines. About 40 ml of urine samples were collected in 50 ml plastic tubes that contained 0.5 ml of 0.5 M EDTA, pH 8.0, and sent to Department of Urology, Faculty of Medicine, The University of Tokyo, where DNA was extracted as described previously (Kitamura et al., 1990).
The 610 bp IG region was amplified by polymerase chain reaction (PCR) using primers P3 and P4 (Miranda et al., 2003). The IG region of the viral genome encompasses the 3′-terminal regions of both the T-antigen and VPl genes, and was established as a region of the JCV genome that contains abundant type-determining sites (Ault and Stoner, 1992). The reaction was carried out for 50 cycles with PWO DNA polymerase (Roche Diagnostics, Tokyo, Japan) or ProofStart DNA polymerase (QIAGEN GmbH, Hilden, Germany). The amplified fragments were cloned into the vector pBluescript II SK (+) (Stratagene, La Jolla, USA) (Miranda et al., 2003), and purified recombinant plasmids were sequenced with an autosequencer (ABI PRISM 373S DNA Analyzer, Applied Biosystems, Foster City, USA).
A neighbor-joining (NJ) phylogenetic tree (Saitou and Nei, 1987) was constructed using the CLUSTAL W program (Thompson et al., 1994). Divergences were estimated with the two-parameter method (Kimura, 1980). The phylogenetic tree was visualized using TREEVIEW (Page, 1996). To assess the confidence of branching patterns of the NJ trees, 1000 bootstrap replicates were performed (Felsenstein, 1985).
To test for genotypic differentiation between the populations, a log-likelihood (G)-based exact test was performed using Genepop version 3.1c at http://www.biomed.curtin. edu.au/genepop (Raymond and Rousset, 1995). The principle of this test is the same as the probability test (or Fisher exact test). The null hypothesis (H0) tested was: ‘the genotypic distribution is identical across populations’. The Markov chain parameters used were: dememorization: 1000; batches: 100; iterations per batch: 1000.
Using a PCR that amplifies the IG region, we detected JCV DNA from 11 (22%) of the 49 urine samples collected from Mamanwas of Surigao province. This detection rate is lower than rates reported previously for the same age group (40 years or older) in many other populations (Kitamura et al., 1994; Agostini et al., 1996; Chang et al., 1999; Saruwatari et al., 2002a, b; Miranda et al., 2003). Nevertheless, lower detection rates for JCV DNA were also reported for some ethnic groups in Central Africa (Chima et al., 1998) and Northeast Siberia and the Arctic regions (Sugimoto et al., 2002).
We cloned IG regions amplified from the Mamanwa urine samples, and sequenced representative clones for each urine sample. We obtained single sequences from most urine samples, but detected two independent sequences from one urine sample (no. 29) (Table 1). From the 12 IG sequences (including both sequences from no. 29), together with reference sequences in Asia and the western Pacific (Table 2), a phylogenetic tree was constructed using the NJ method (Saitou and Nei, 1987). According to the resultant phylogenetic tree (Figure 1), half of the Mamanwa sequences were found in the B3-b/2E cluster and half in the SC-f/7A cluster. As two systems have been developed to designate JCV genotypes, we describe here a genotype named B3-b (Miranda et al., 2003) or Type 2E (Jobes et al., 2001) as B3-b/2E and a genotype named SC-f (Saruwatari et al., 2002b) or Type 7A (Agostini et al., 2001) as SC-f/7A. Although the bootstrap probabilities for these clusters were 50% or lower, representative isolates (shown by asterisks in Figure 1) formed distinct clusters with higher bootstrap probabilities in a phylogenetic analysis based on complete sequences (Takasaka et al., 2004). In addition, the genotypes of JCV based on the phylogenetic analysis were consistent with genotype-specific variations of the noncoding control region (Guo et al., 1996; Chang et al., 1999; Ryschkewitsch et al., 2000; and data not shown).

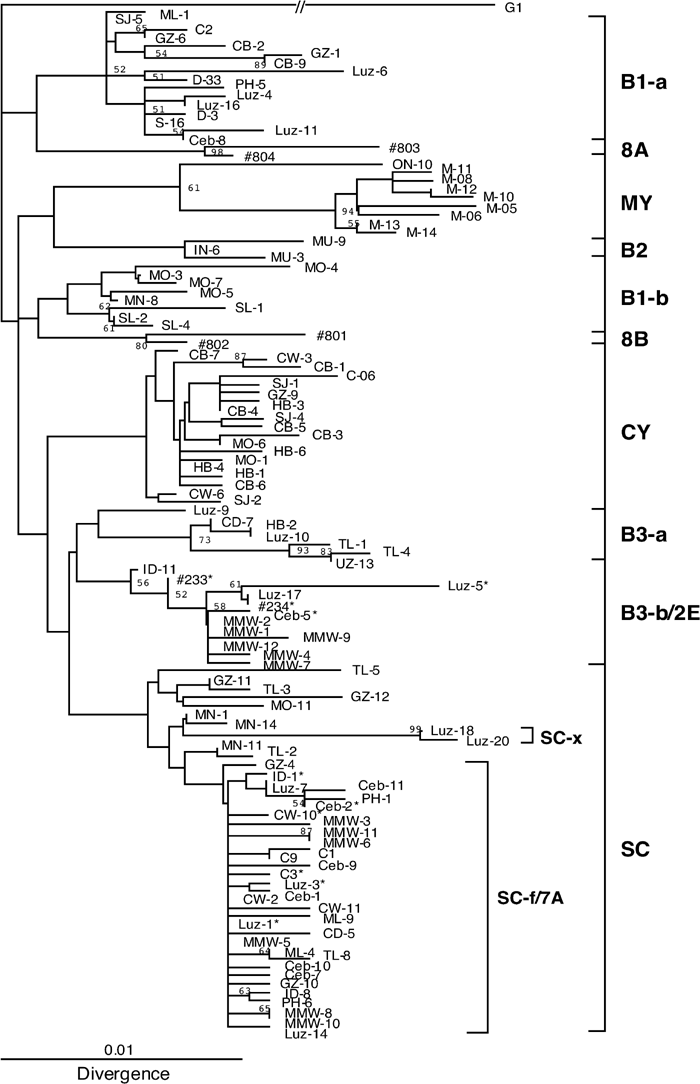 View Details | Figure 1. Phylogenetic tree used to classify the JCV isolates detected into distinctive genotypes. IG sequences in Mamanwas together with those detected previously in Asia and Oceania were used to construct an NJ phylogenetic tree using CLUSTAL W (Thompson et al., 1994). The phylogenetic tree was visualized using TREEVIEW (Page, 1996). The tree was rooted using a European isolate (G1) as the outgroup. Genotypes are indicated at the right of the tree. Asterisks identify isolates (within the B3-b/2E and SC-f/7A genotypes) previously analyzed using the whole genome approach (Takasaka et al., 2004). Origins of isolates are shown in Tables 1 and 2. The numbers at nodes give bootstrap confidence level (%) obtained for 1000 replicates (only values ≥50% are shown for major nodes). |
Table 3 shows the JCV genotype frequencies in three populations in the Philippines (Mamanwa, Tagalog, and Cebuano). The data for the Tagalog and Cebuano were from our previous report (Miranda et al., 2003). SC-f/7A and B3-b/2E each accounted for 50% of all genotypes detected in the Mamanwa. This was in contrast with the profiles for the other Philippine populations which carried up to five genotypes, with B3-b/2E showing only a low frequency. The test for genotype differentiation showed that the JCV genotypic distribution in the Mamanwa was significantly different from that in the Tagalog and Cebuano populations (Table 4).

Results of the present study show that the JCV genotype profile of the Mamanwa is significantly different from the profiles of two other Philippine populations, the Tagalogs of Luzon island and the Cebuanos of Cebu island, in which up to five genotypes (B1-a, B3-a, B3-b/2E, SC-f/7A, and SC-x) were detected and B3-b/2E had only a low frequency (Miranda et al., 2003). The JCV genotype patterns in the Tagalogs and the Cebuanos were somewhat consistent with the traditional view about the formation of the modern Philippine population (see Introduction), although it also suggested some previously undescribed migrations to the Philippines, recent as well as ancient (Miranda et al., 2003). In contrast, the JCV genotype profile of the Mamanwa suggests that this tribe was formed by two major events of colonization, one involving carriers of B3-b/2E and the other, carriers of SC-f/7A.
The question then arises as to which colonization occurred first. Before discussing this issue, we will summarize recent findings about the JCV genotypes in Oceania. Seven oceanic JCV genotypes (B1-a, B3-a, B3-b/2E, SC-f/7A, SC-x, 8A, and 8B) have been described (Ryschkewitsch et al., 2000; Jobes et al., 2001; Yanagihara et al., 2002; Miranda et al., 2003; Takasaka et al., 2004). All of these genotypes belong to the Type-B supercluster from which most Asian genotypes were generated (Takasaka et al., 2004). The Oceanic genotypes were classified into three categories (herein designated groups 1–3) on the basis of their geographic distribution patterns (Takasaka et al., 2004). Group 1 includes three genotypes (SC-f/7A, B1-a, and B3-a) distributed on the Asian continent as well as islands near the continent; group 2 includes two genotypes (B3-b/2E and SC-x) distributed in islands near the Asian continent as well as in the western Pacific; and group 3 includes two genotypes (8A and 8B) distributed only in the western Pacific. It was suggested that groups 2 and 3, which are rarely detected on the Asian continent, represent ancient human migration, while group 1, which is detected both in the Pacific and on the Asian continent, represents recent human migration (Takasaka et al., 2004). We detected B3-b/2E and SC-f/7A equally in the Mamanwa. According to the classification above, B3-b/2E and SC-f/7A are classified as group 2 and group 1 genotypes, respectively. Thus, the occurrence of B3-b/2E in the Mamanwa may suggest ancient colonization, while the occurrence of SC-f/7A may represent recent colonization.
Ryschkewitsch et al. (2000) recently reported the JCV genotype profile of the indigenous tribe (the Chamorro) in Guam. Of the 20 isolates examined, nine (45%) were classified as B3-b/2E, and eight (40%) as SC-f/7A (the others were European types probably imported recently). This genotype profile suggests that Chamorros have a population history similar to that assumed for the Mamanwa, i.e. an earlier colonization by people carrying B3-b/2E followed by later migrants carrying SC-f/7A. It is tempting to postulate that other indigenous tribes in Southeast Asian and Oceanic islands near the Asian continent may have analogous population histories. If our hypothesis is correct, then the genetic differences previously observed between the Mamanwa and other Philippine and Asian populations (Matsumoto et al., 1979; Omoto, 1980, 1984; Horai et al., 1981; Omoto et al., 1981; Davila et al., 2002) may be explained by the difference in the ratios of the two ancestral populations carrying B3-b/2E or SC-f/7A.
Interestingly, a close correlation between the Mamanwa and the Chamorro was previously suggested by the genetic observation that a carbonic anhydrase-1 variant called CA1-3N frequently occurs in both the Mamanwa and the Chamorro (Omoto, 1980; Omoto et al., 1981). Furthermore, this variant shows a broad distribution in the western Pacific (Omoto, 1980; Omoto et al. 1981). The languages spoken by islanders in a wide area of the western Pacific, including the Mamanwas and Chamorros, belong to the Austronesian family (Ruhlen, 1987). All in all, it is likely that ancient Austronesians carried the CA1-3N variant as well as the B2-b/2E genotype of JCV and that as they spread in Oceania, the genetic as well as JCV markers expanded in the region. An analogous view was proposed by Yanagihara et al. (2002), who suggested that Austronesians carrying B3-b/2E migrated into Oceania 3500 years ago or more; the earliest migrants (ancestors of the modern Papuans and Melanesians) were thought to have brought genotypes 8A and 8B about 40,000 years ago (Yanagihara et al., 2002).
We are grateful to all urine donors. This study was supported in part by grants from the Ministry of Education, Culture, Sports, Science and Technology, Japan and from the Ministry of Health, Labour and Welfare, Japan.