| Collected Papers in Honor of Professor Emeritus Banri Endo: Commemoration of His Seventieth Birthday M. Pietrusewsky, corresponding author. e-mail: mikep@hawaii.edu phone: +1-808-956-6653; fax: +1-808-956-9541 Published online 13 July 2004 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.03107 |
A recent multivariate analysis of measurements recorded in male crania (Pietrusewsky, 1999) reached a number of conclusions regarding the historical-biological relationships of the recent and earlier inhabitants of the Japanese archipelago, including the Ryukyu Islands. Among those conclusions was the demonstration of a connection among the Ryukyu Island cranial series and those representing the Yayoi, Kofun, and Kamakura periods of Japan—groups that, in turn, were well differentiated from modern Japanese, Ainu, and Jomon cranial series. The results also reiterated a relatively close connection between the Jomon and Ainu cranial series, a connection that has been demonstrated in numerous studies (e.g. Howells, 1966; Turner, 1976; K. Hanihara, 1985, 1998; Mizoguchi, 1986; Brace et al., 1989; Ossenberg, 1992; Yamaguchi, 1992; Ishida, 1996; Omoto et al., 1996; Matsumura, 2001; Dodo and Kawakubo, 2002). The absence of evidence for a close affinity between any of the Ryukyu Island cranial series and the prehistoric Jomon and modern Ainu cranial series has been supported by other work in physical anthropology (Pietrusewsky, 1996; Doi et al., 1997; Omoto and Saitou, 1997; Dodo et al. 1998).
In this new analysis, multivariate statistical procedures were applied to female crania representing virtually the identical groups from the Ryukyu Islands and the main islands of Japan investigated in Pietrusewsky (1999). The results of this female analysis are compared with the results of the earlier male analysis as well as with other studies that have examined biological relationships of the earlier and later inhabitants of the Japanese Archipelago. The approach used is model free, and measures of biological distance are used to investigate patterns of craniometric variation for assessing biological relationships and population history regardless of exact causes (Pietrusewsky, 2000).
Sixteen cranial series, totaling 428 female adult specimens, representing the Ryukyu Islands and the main islands of Japan, are included in this study (Table 1, Figure 1). The Ryukyu Islands cranial series includes Tokunoshima and Yoro Islands in the Amami Group; Okinawa, Gyokusendo Cave, and Kume Island in the Okinawa Group; and Sakish-ima Islands in the southern Ryukyus. The specimens represent modern or near-modern inhabitants of the Ryukyu Islands, and were collected at least a half century ago from traditional mortuary contexts (caves and/or rockshelters). All data were recorded by the author on two separate occasions in 1990 and 1997, respectively. Drs Hiroto Takamiya, Naomi Doi, and Mark Hudson assisted with the data collection on Okinawa Island in 1997. In addition to the cranial series from the Ryukyu Islands, female crania representing prehistoric, historic, and more modern Japanese (and Ainu) are included in these analyses. The sex and age of each specimen were determined by the author using standard criteria (e.g. Krogman and Iscan, 1986; Buikstra and Ubelaker, 1994).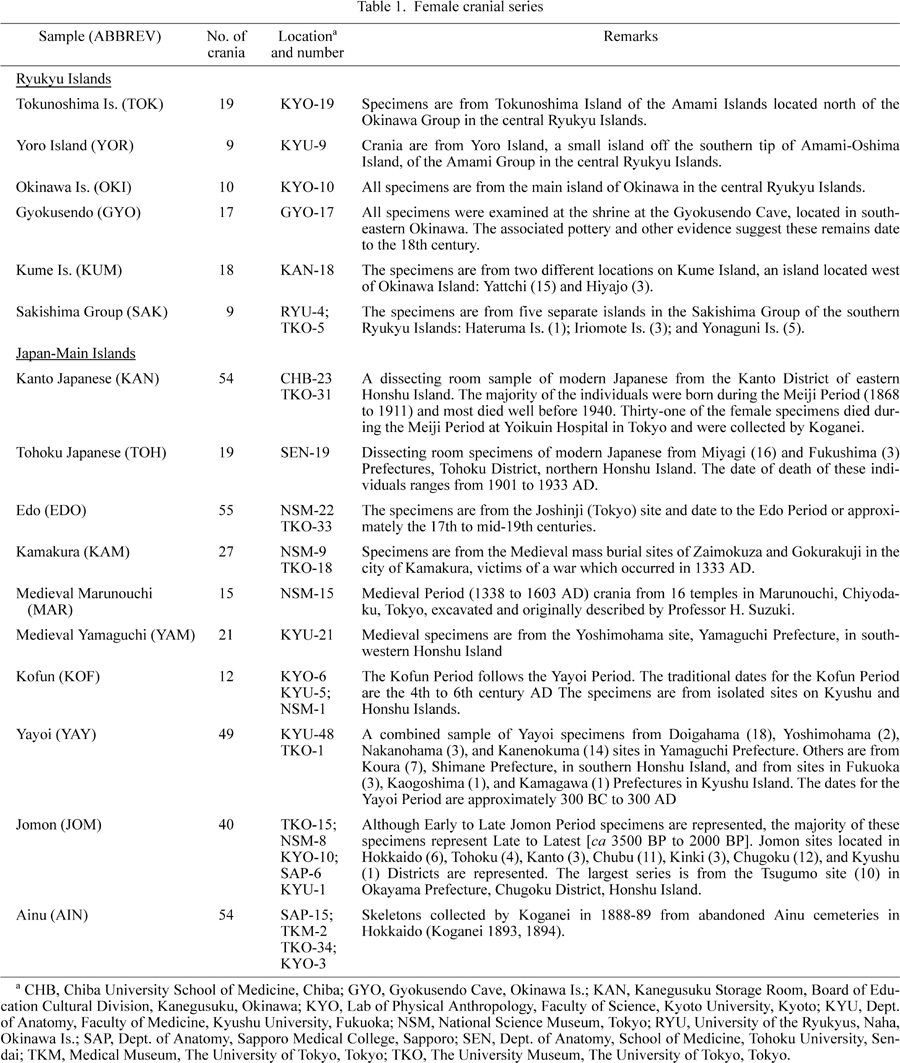
 View Details | Figure 1. Map of Japan showing the geographical locations of the six Ryukyu and ten female Japanese cranial series used in this analysis. |
The data consist of 29 standard craniometric landmark measurements, the largest number of variables common to all series compared. The measurements, described in Table 3 and Table 4, include linear dimensions of the cranial vault and face. Because the multivariate statistical procedures used in the present study require complete sets of data, missing measurements were replaced through stepwise regression analysis using the computer program PAM (Dixon and Brown, 1979). Since complete or nearly complete specimens were initially selected, this procedure was used sparingly.
Two multivariate statistical procedures, multiple stepwise discriminant function analysis (canonical analysis) and Mahalanobis’ generalized distance (D2), were applied to 29 cranial measurements.
The computer program BMDP-7M (Dixon and Brown, 1979) was used to perform the multiple stepwise discriminant function analysis. This program selects the original measurements to compute the linear classification functions in a stepwise manner such that, at each step, the measurement that adds the most to the separation of the groups is entered into the discriminant function in advance of the others. The first few canonical variates account for the majority of the variation among groups. In this analysis, the group means are plotted on the first three canonical variates, or functions, for interpreting intergroup relationships.
Mahalanobis’ generalized distance (Mahalanobis, 1936; Mahalanobis et al., 1949) was applied to the same data analyzed by discriminant function analysis. Generalized distance analysis provides a single quantitative measure of dissimilarity (distance) between individual groups using many variables while accounting for intercorrelations between the variables.
The significance of the distances was determined using the methods described in Rao (1952, p. 245) and Buranarugsa and Leach (1993, p. 17). Non-significance of D2 generally indicates that the differences are too small for detection of group differences and/or that the sample sizes are too small. The SAHN clustering method in the NTSYS-pc computer software program (Rohlf, 1993), which uses the average linkage within-group clustering algorithm (Sneath and Sokal, 1973), was used to construct the dendrograms using Mahalanobis’ distances. This algorithm combines clusters such that the average distance among all cases in the resulting cluster is as small as possible. The distance between two clusters is taken to be the average distance among all possible pairs of cases between the two clusters.
Two separate applications of multivariate statistical procedures to 29 cranial measurements are reported. The first analysis compares six female Ryukyu cranial series and the second analysis compares 16 female Ryukyu and Japanese cranial series. The means and standard deviations for the 29 cranial measurements recorded in 16 female Ryukyu and Japanese cranial series are presented in Table 2.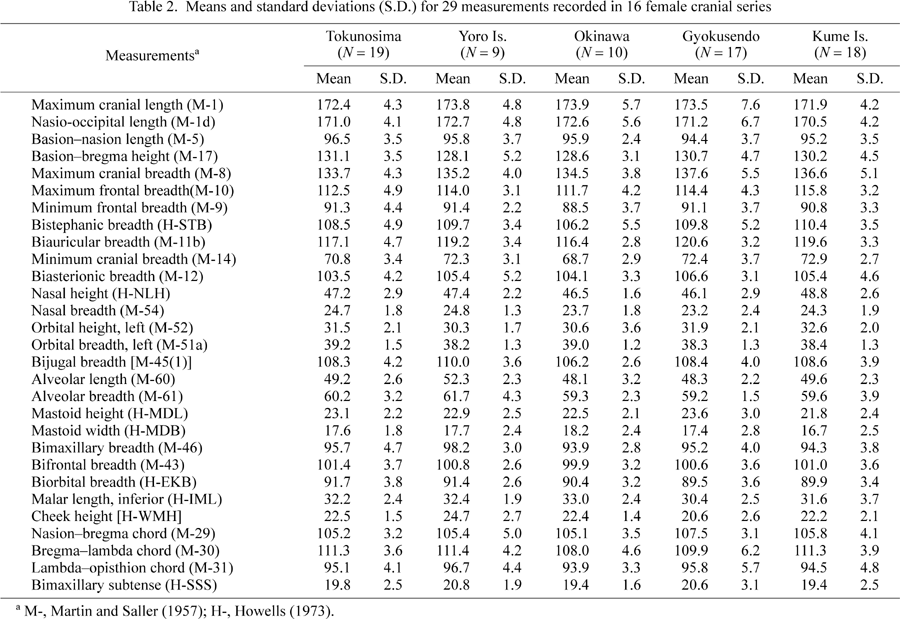


Stepwise discriminant function analysis
A summary of the 29 measurements, ranked according to the F-values received in the final step of discriminant function analysis, is given in Table 3. Among the measurements contributing most to the discrimination are cheek height, minimum cranial breadth, orbital breadth, biauricular breadth, and bimaxillary breadth.
Eigenvalues, the percentage of total dispersion, cumulative percentage of dispersion, and level of significance for the first five canonical variates are presented in Table 4. The first three canonical variates account for 80.9% of the total variation produced. Only the first eigenvalue is significant at the 1% level.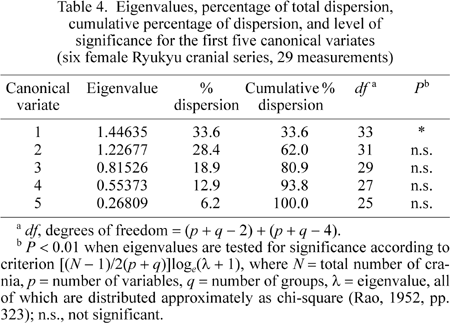
Canonical coefficients for 29 measurements for the first three canonical variates (Table 5) indicate that the most important variables in producing group separation in the first canonical variate are orbital breadth, nasio-occipital length, maximum cranial length, and maximum frontal breadth. Orbital breadth, bifrontal breadth, bijugal breadth, and biorbital breadth are primarily responsible for the discrimination produced in the second canonical variate. Cheek height, orbital breadth, alveolar length, and maximum frontal breadth are primarily responsible for group separation produced in the third canonical variate.
A summary of the group classification results, based on posterior probabilities (Table 6), indicates that Yoro, Gyokusendo, Kume, and Sakishima achieve the best classification results, with over 80% of the cases in these cranial series being correctly classified to their original groups. The poorest classification results are those for the Okinawa and Tokunoshima cranial series.
The jackknifed classification results (Table 7) are uniformly poor for all cranial series. Yoro, Gyokusendo, and Sakishima have the best classification results. Tokunoshima achieves the poorest classification results, with only 2/19 (10.5%) of its cases being correctly classified. These results reiterate the relative cranial morphological homogeneity of the female Ryukyu Island cranial series.
A plot of the group means on the first two canonical variates (Figure 2) reveals two major clusters, one which includes Tokunoshima and Okinawa, and the other which contains Gyokusendo, Kume, and Sakishima. Yoro occupies an isolated position in this diagram. A plot of the means for these same cranial series on the first three canonical variates (Figure 2), emphasizes these associations.
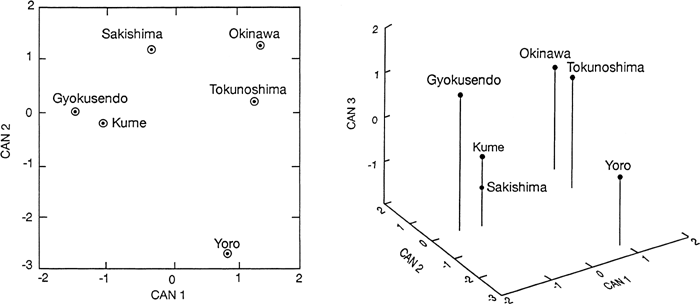 View Details | Figure 2. Plot of six female Ryukyu group means on the first two canonical variates (left) and on the first three canonical variates (right), which result from discriminant function analysis. |
Mahalanobis’ generalized distance
Mahalanobis’ distances and tests of significance using 29 measurements and six female Ryukyu cranial series (Table 8) indicate that none of the distances are significant at the 5% level. The smallest distances are those between Tokunoshima and Okinawa (3.907), Kume and Gyokusendo (5.697), and Kume and Tokunoshima (7.600), respectively. The largest distances involve the Yoro series. The dendrogram (Figure 3) created when the UPGMA clustering algorithm is applied to these distances reveals that Okinawa and Tokunoshima occupy a single cluster as do Gyokusendo and Kume. Sakishima and Yoro occupy isolated positions in this diagram.
 View Details | Figure 3. Diagram of relationship based on a cluster analysis (UPGMA) of Mahalanobis’ generalized distances for six female Ryukyu cranial series. |
In this next analysis, stepwise discriminant function analysis and Mahalanobis’ generalized distance are applied to 29 measurements recorded in 16 female Ryukyu and Japanese cranial series.
Stepwise discriminant function analysis
A summary of the 29 measurements, ranked according to the F-values received in the final step of discriminant function analysis (Table 9) indicates that the first 17 measurements contribute most to the discrimination produced. In this analysis, bijugal breadth, cheek height, basion-nasion length, nasal height, and biauricular breadth are ranked the highest in overall ability to discriminat. 
Eigenvalues for the first 15 canonical variates (Table 10), in this analysis, indicate that the first three canonical variates account for 56.2% of the total variation. All eigenvalues in this table are significant at the 1% level.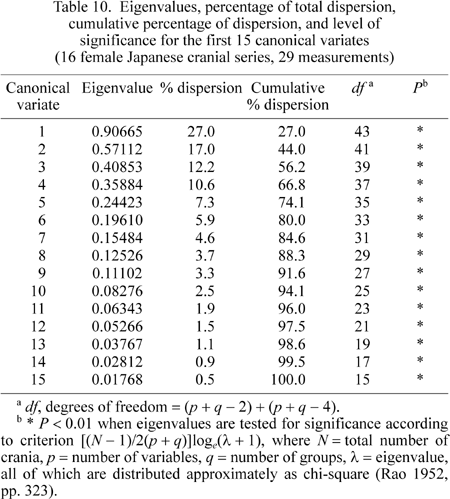
Canonical coefficients for 29 measurements for the first three canonical variates are given in Table 11. Orbital breadth, bifrontal breadth, bijugal breadth, nasal height, and naso-occipital length (those variables with the highest coefficients regardless of sign) are the most important variables in producing group separation in the first canonical variate. Orbital height, cheek height, bijugal breadth, and minimum cranial breadth are most responsible for group separation produced in the second canonical variate. Bimaxillary breadth, malar length, biorbital breadth, and orbital height are primarily responsible for the discrimination produced in the third canonical variate.
A summary of the group classification results, based on posterior probabilities (Table 12), indicate that Yoro, Gyokusendo, Okinawa, and Jomon achieve the best classification results, with over 70% of the cases being correctly classified to their own groups. The poorest classification results are those for Kanto, Tohoku, Yayoi, and Edo, with less than 40% of these cases being classified to their original group. Two of the Kume cases are reclassified as Kofun and two of the Sakishima specimens are classified as Yayoi.
Jackknifed classification results (Table 13) reveal a much higher percentage of misclassifications. Kofun, Kanto, Gyokusendo, Sakishima, and Okinawa register some of the poorest classification results, while Ainu and Jomon achieve the best classification results. Several of the cases originally assigned to Okinawa, Yoro, and Sakishima are reclassified as Yayoi, Kamakura, and Kofun.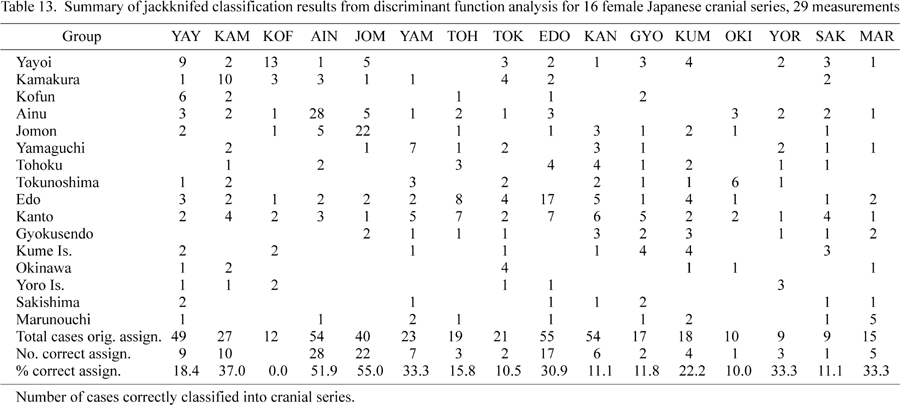
A plot of the group means on the first two canonical variates (Figure 4) reveals three distinct groupings. Two medieval (Yamaguchi and Marunouchi) and three near modern Japanese series (Kanto, Tohoku, and Edo) form one of these clusters. Kofun, Yayoi, and Kamakura and six Ryukyu series form another grouping. The most isolated cluster is the one occupied by Jomon and Ainu. The plot of the means for these same groups on the first three canonical variates (Figure 4) positions the six Ryukyu series between the near modern Japanese series and a group consisting of Yayoi, Kofun, and Kamakura. Kamakura is closest, in this representation, to Yoro, one of the Ryukyu series. The Jomon and Ainu cranial series again are relegated to isolated positions in the plot.
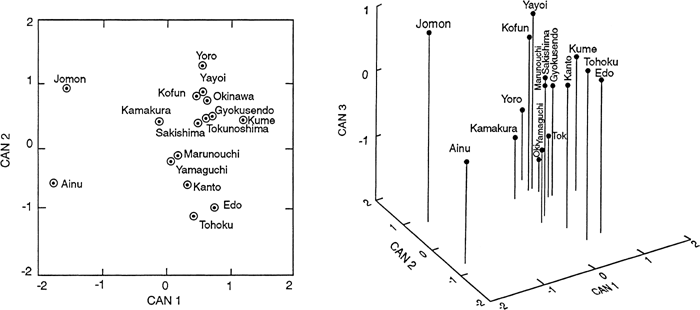 View Details | Figure 4. Plot of 16 female Japanese group means on the first two canonical variates (left) and on the first three canonical variates (right) which result from discriminant function analysis |
Mahalanobis’ generalized distance
Mahalanobis’ distances and tests of significance using 29 measurements for 16 female Ryukyu and Japanese cranial series are presented in Table 14. Applying the UPGMA clustering algorithm to the distances in Table 14 results in the dendrogram presented Figure 5 Three modern and near-modern Japanese series (Tohoku, Edo, and Kanto) and the medieval series from Yamaguchi prefecture form a separate cluster in this diagram, which then attracts a cluster that includes Kamakura, Kofun, and Yayoi. The Ryukyu Island series and the Marunouchi medieval series are grouped with these latter. The Jomon–Ainu cluster is the last to join this association.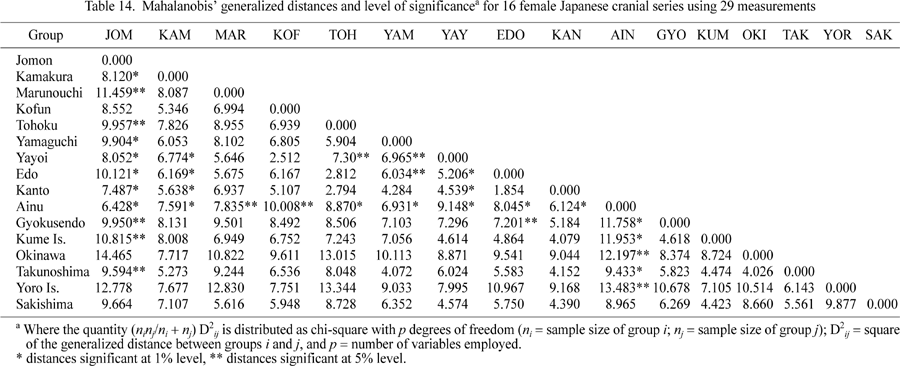
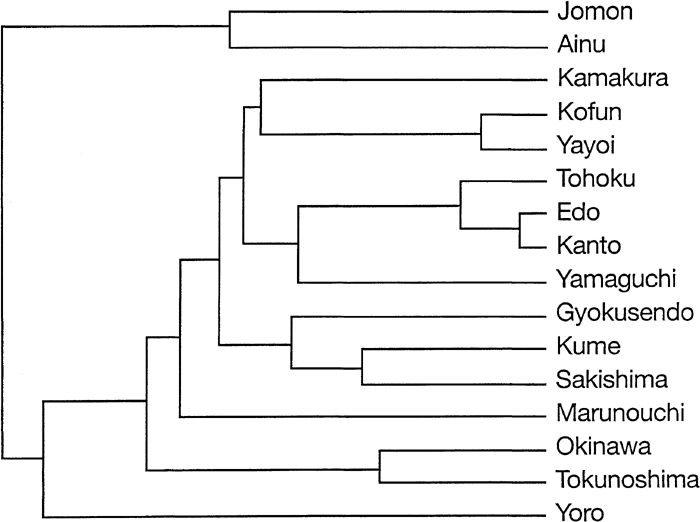 View Details | Figure 5. Diagram of relationship based on a cluster analysis (UPGMA) of Mahalanobis’ generalized distances for 16 female Japanese cranial series. |
Comparisons of six female Ryukyu cranial series in the present analysis demonstrate close similarities with the relationships reported in an analysis based on male craniometric data (Pietrusewsky, 1999). As was the case in the male analysis, overall, there is a general lack of marked differentiation among the female Ryukyu cranial series, which suggests relative craniometric homogeneity for this region. A similar conclusion was reached by Dodo et al. (1998) using cranial nonmetric traits recorded in Ryukyu crania. The patterning that is found generally reflects geography and other isolating mechanisms such as genetic drift.
As was found in the analysis of male craniometric data (Pietrusewsky, 1999), the female cranial series from Gyokusendo Cave on Okinawa Island and Kume Island, an island in the Okinawa group, are closely related. Likewise, cranial series representing Okinawa and Tokunoshima (an island in the Amami group north of Okinawa), demonstrate a morphological closeness using both male and female data, and the cranial series representing the southernmost Ryukyu Islands (Sakishima) was found to be related to the cranial series from the Okinawa and Amami groups. Finally, the most isolated cranial series in both the female and male analyses is the one representing Yoro Island, a small island located off the southern tip of Amami-Oshima Island in the Amami group. The geographical isolation of this island may help to explain this result.
Turning to the results of the second analysis presented in this study, again there is broad agreement between the female results and those based on male data (Pietrusewsky, 1999). In both studies, the near-modern Japanese cranial series, Tohoku, Edo, and Kanto, are closely related. Likewise, the Kamakura, Kofun, and Yayoi cranial series form a close association that is isolated from the more modern cranial series. The marked separation of the modern Japanese cranial series since Yayoi times and the Jomon–Ainu series agrees with the general consensus among biological anthropologists that a major influx of new people arrived in the Japanese archipelago (via the Korean peninsula) beginning with the Yayoi (ca. 400 BC to 300 AD) and Kofun (4th to 7th century AD) periods. This view has been popularized as the ‘dual structure model’ by Kazuro Hanihara (1991).
As was demonstrated in the male analyses (Pietrusewsky, 1999), the female Ryukyu Island cranial series are also more closely related to the Kamakura, Kofun, and Yayoi cranial series than they are to the modern Japanese series. The medieval Marunouchi and Yamaguchi cranial series, while dispersed, have a tendency to associate with Ryukyu Island samples (see especially Figure 4). The separation of modern Japanese from the modern inhabitants of the Ryukyu Islands and their presumed ancestors (Yayoi, Kofun, and medieval series) may reflect changes attributable to the effects of modernization in skull form in the modern Japanese since the Yayoi period (Pietrusewsky, 1999). Likewise, the modern inhabitants of the Ryukyu Islands may have differentially retained features of skull form traceable to the Yayoi immigrants.
The Jomon and Ainu cranial series form a cluster well removed from all remaining series in both the male and female analyses. The results based on female data further indicate a close connection between Ainu and Jomon, a finding that reiterates a great many previous studies (e.g. Howells, 1966, 1986; Turner, 1976, 1990; Yamaguchi, 1982, 1992; K. Hanihara, 1985; Dodo, 1986; Mizoguchi, 1986; Brace et al., 1989; Matsumura, 1989, 2001; Kozintsev, 1990; Ossenberg, 1992; K. Hanihara et al., 1993; T. Hanihara, 1993; Ishida, 1996; Omoto et al., 1996; Pietrusewsky, 1996, 2000; Ishida and Kondo, 1999; Dodo and Kawakubo, 2002).
Neither the male or female results support an especially close affinity between the Ryukyu Island and the Ainu/Jomon cranial series, a conclusion supported by other work (e.g. Mouri, 1988; Pietrusewsky, 1994, 1996, 1997; Horai et al., 1996; Doi et al., 1997; Dodo et al., 1998). This conclusion stands in marked contrast to other studies that have suggested a Ryukyu–Ainu–Jomon connection (e.g. Baelz, 1911; Newman and Eng, 1947; K. Hanihara et al., 1974, 1986, 1991; Yamaguchi, 1982, 1992; T. Hanihara, 1989, 1991, 1992; Omoto, 1992; Matsumura, 1995; Omoto and Saitou, 1997).
Finally, the present results do not support the results reported by Brace and co-workers (e.g. Brace et al., 1989; Brace and Tracer, 1992), who claim that the Kamakura series may represent Ainu.
The results of this new multivariate craniometric study, based on female data, allow a number of tentative conclusions regarding the biological relationships of Ryukyu Islanders and their relationship to prehistoric and modern Japanese.
1. As was found in previous work that used male cranial series, the female Ryukyu cranial series are relatively homogeneous. The cranial series from Yoro Island, a small island in the Amami group, is the most differentiated of the Ryukyu Island series.
2. The majority of the Ryukyu cranial series are closest to the Yayoi, Kofun, and Kamakura cranial series, and are well differentiated from modern Japanese, Jomon, and Ainu cranial series. A similar finding using male data suggests that the immigrants from the north commencing during the Yayoi period, or from approximately 400 BC, influenced the inhabitants of the Ryukyu Islands.
3. Comparisons using cranial series from the Japanese archipelago, including the Ryukyu Islands, indicate that the Jomon and Ainu groups are the most differentiated groups, suggesting a separate origin for the Jomon and their presumed descendants, the Ainu.
4. There is no evidence for a close connection between Ryukyu and the Jomon/Ainu cranial series.
A biological similarity between the Ryukyu series and those representing Yayoi, Kofun, and the medieval series from Kamakura, demonstrated by both male and female craniometric data, warrants further investigation.
Except for the new female cranial series examined in 1997 on Okinawa Island, permission to examine the cranial series used in the present study has been acknowledged previously (Pietrusewsky, 1994, 1995, 1996, 1997, 1999; Pietrusewsky and Chang, 2003). I wish to especially thank Drs Hiroto Takamiya (Sapporo University), Naomi Doi (University of the Ryukyus), and Mark Hudson (formerly of the University of Tsukuba) who facilitated my visit to Okinawa Island in December of 1997, and who assisted me in recording data at Gyokusendo Cave, the Kanegusuku Storage Facility, Board of Education Cultural Division, and at the University of the Ryukyus. Ms Rona Ikehara-Quebral assisted in the analysis of the data and Ms Billie Ikeda, of the Center for Instructional Support of the University of Hawaii, produced the figures used in this paper. My thanks to Dr Michele Toomay Douglas for her helpful comments in preparing this paper. Research for the present paper was initiated while a Visiting Professor at the International Research Center for Japanese Studies in Kyoto in 1997–1998. I wish to acknowledge the kind support of the International Research Center for Japanese Studies and Professor Keiichi Omoto, who was my sponsor during my stay in Kyoto. Finally, a great deal of the data used in this paper was recorded while a Fulbright Scholar to Japan in 1990. During that stay, Professor Banri Endo was my sponsor. I owe a multitude of thanks to Professor Endo for all his kindness and assistance extended to me during my stay at the University Museum of The University of Tokyo.
|