| E. Maschenko, corresponding author. e-mail: evmash@paleo.ru phone: +7-095-116-49-71; fax: +7-095-339-06-22 Published online 10 December 2004 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.04S015 |
The early history of the order Primates from the eastern part of Eurasia reflects the restructuring of the mammalian faunas of Eurasia and North America that occurred at the Paleocene–Eocene boundary at about 57 Ma. The appearance of the order Primates in Asia is interrelated with the advent of certain groups of American mammals in Asia. The most ancient finds of primates from Asia are dated to the Early Eocene (Tsagan Khushu, Mongolia in the southern Gobi) (Dashzeveg and McKenna, 1977; Gingerich et al., 1991; Maschenko, 2002).
Primates became a frequent component of the mammalian faunas of the Late Miocene of Russia and adjacent areas (Late Turolian, MN 11–13, about 8 Ma). In terms of zoogeography, the southern part of eastern Europe, theTranscaucasus, and Central Asia were part of the Mediterranean province. The taxonomic composition of primates of this area during this time span does not differ much from that of typical Mediterranean faunas, except in the presence of some endemic species. Primates of this area are represented by one genus (and species) of Hominoidea, Dryopithecus, and by one genus and two? species of the subfamily Colobinae (Table 1). During the Late Miocene, the apes apparently became extinct. Later, in the Early Pliocene and Pleistocene, Central Asia was most probably linked zoogeographically with Africa. Such a link would explain the distribution of some cercopithecid genera of African affinity.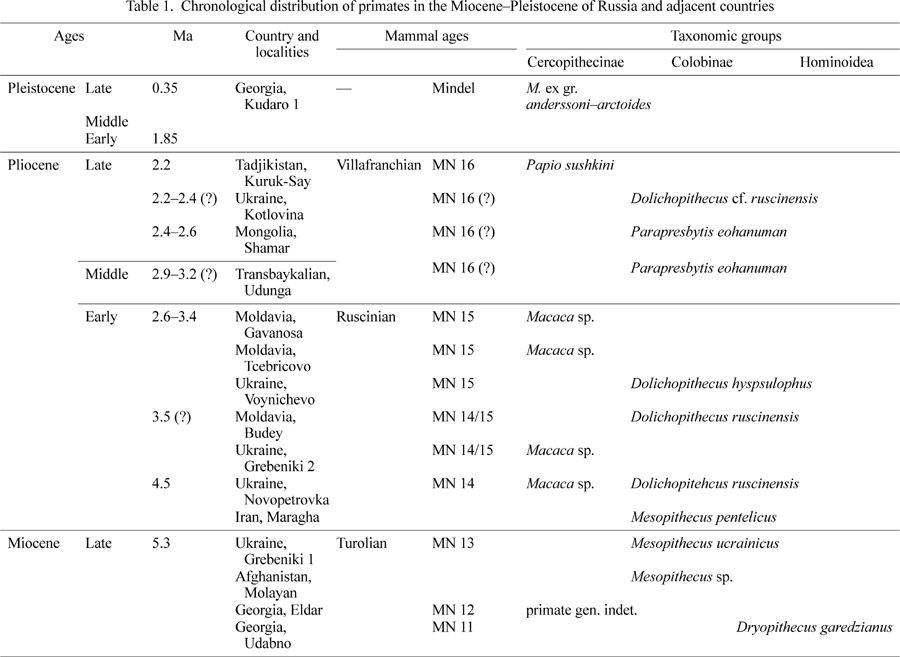
Abbreviations in the text and figures are as follows: PIN, Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; GIN, Geological Institute, Russian Academy of Sciences, Moscow, Russia; GIN U, Geological Institute Siberian Branch, Russian Academy of Sciences, Ulan-Ude, Russia; ZIN, Zoological Institute, Russian Academy of Sciences, St Petersburg, Russia.
Family Omomyidae Trouessart, 1879
Altanius Dashzeveg and McKenna, 1977
Altanius orlovi Dashzeveg and McKenna, 1977
This is the most ancient and the only Asian representative of the family Omomyidae. Altanius orlovi is one of the smallest and most specialized representatives of the order Primates. Morphologically it is close to North American anaptomorphines. It is distinguished by a reduced P4 talonid. Metaconid is well expressed in the P4. The M1 trigonid is open, and the paraconid is cone-shaped and not reduced. The M3 has a large paraconid displaced lingually and an elongated talonid.
Additional studies of A. orlovi revealed two forms (species?) with similar dental morphology that differ only in size. The status of the larger form of Altanius sp. is unclear. The larger form has a 30% deeper mandibular corpus than the smaller one. Differences between the two forms in postcanine tooth size are insignificant. A more precise evaluation may be possible when more data are available on variability and sexual dimorphism of the species (Maschenko, 2001, 2002).
Distribution. Late Paleocene(?)–Early Eocene, Tsagan Khushu locality, Gashatan-Bumbanian, southern Mongolia, Central Asia.
In 1908, Professor V.D. Laskarev reported finds of Oreopithecus sp. from Kalfa, Moldavia (Late Turolian) (Laskarev, 1908) (Figure 1). The material, on which the communication was based, was later lost. However, given that the tribe Oreopithecini is endemic to central Italy, the report may by unreliable. The taxonomy of primates from the Kalfa locality remains uncertain. So, by the beginning of the Ruscinian, no hominoid representatives are known from the geographic area considered in this review, whereas Cercopithecidae became more abundant. Several finds from southern Russia were known from the beginning of the 20th century (Homenko, 1915). At the end of the Miocene, cercopithecids became a common component of the Hipparion faunas of Moldavia, Ukraine, Georgia, northern Iran and Afghanistan. Out of the five known Miocene primate localities, Dryopithecidae is present in one locality and Colobinae in four localities (Gremiatskiy, 1958, 1962; Burchak-Abramovich, 1967, 1973; Maschenko, 1989).
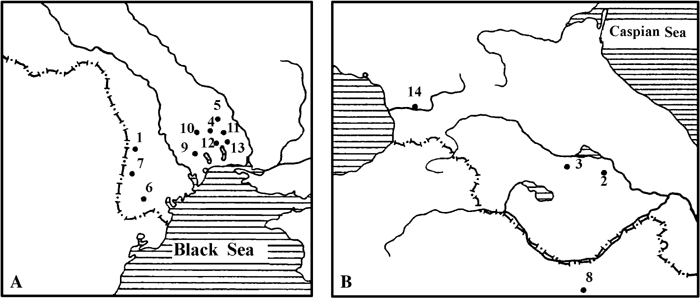 View Details | Figure 1. Major primate localities. (A) Map of the southern part of eastern Europe. (B) Map of the Transcaucasus area. 1, Kalfa, Moldavia; 2, Eldar, Georgia; 3, Udabno, Georgia; 4, Grebeniki 1 and 2, Ukraine; 5, Petroverovka (Novopetrovka), Ukraine; 6, Budey, Moldavia; 7, Gavanosa, Moldavia; 8, Maragha, Iran; 9, Zotven, Ukraine; 10, Novopetrovka, Ukraine; 11, Voynichevo, Ukraine; 12, Kuchurgan (Voynichevo), Ukraine; 13, Kotlovina, Ukraine; 14, Kudaro 1, Georgia. |
Order Primates
Gen. indet.?
The list of mammals of the Hipparion fauna from the Eldar locality, eastern Georgia, contains an entry “the skull of monkey.” No systematic position was determined for this specimen, and no description is known (Gremiatskiy, 1957; Burchak-Abramovich, 1967). All Late Miocene mammalian materials from this locality were collected before 1917 and lost in 1918.
Distribution. Late Miocene (Upper Sarmatian, MN 12?), Eldar locality, eastern Georgia (Figure 1B).
Superfamily Hominoidea Gray, 1825
Dryopithecus Lartet, 1856
Dryopithecus (Udabnopithecus) garedzianus (Burchak-Abramovich and Gabashvili, 1945)
This taxon is represented by a right P4 and M1 in a small upper jaw fragment. The fossil is housed in the State Museum of Georgia, Tbilisi. This is a small-sized Dryopithecus (Figure 2), the only representative of the genus from the geographical area considered in the present review. It indicates a possible link with Dryopithecus of eastern Europe. This demonstrates that links between the Mediterranean and Asia were maintained through the Transcaucasus, hence the zoogeographic significance of this specimen.
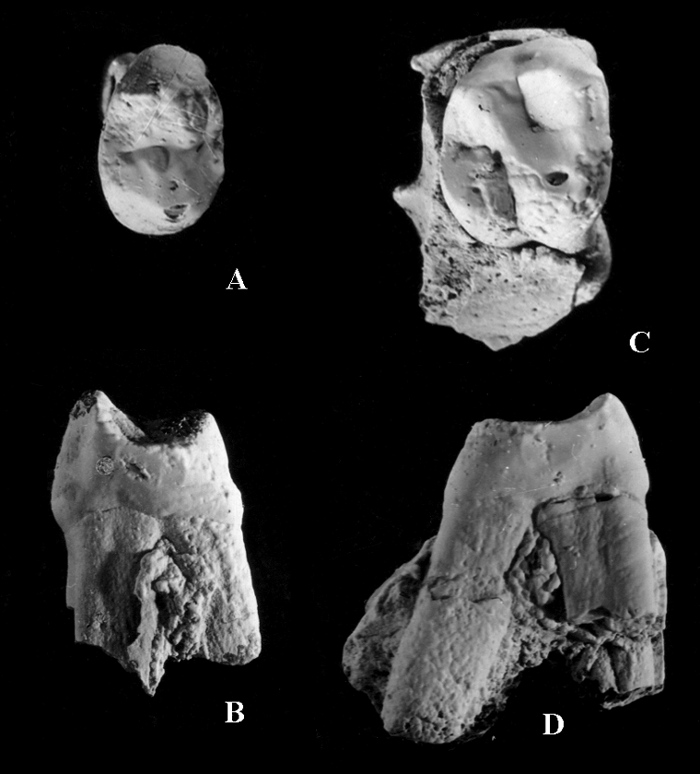 View Details | Figure 2. Right upper P4 and M1 of Dryopithecus garedzianus from the Udabno locality, Georgia, Late Miocene (from Burchak-Abramovich and Gabashvili, 1945). (A) P4 occlusal view; (B) P4 mesial view; (C) M1 occlusal view; (D) M1 mesial view. |
Both teeth are little worn. Tooth dimensions are: P4 length, 6.9 mm; P4 width, 10.1 mm; M1 length, 9.0 mm; M1 width, 10.3 mm. The P4 crown is bicuspid and has three roots. The protocone is the largest cusp. The mesiodistal crown diameter is greater lingually than buccally. A cingulum is present on the lingual side of the crown. The M1 crown is four-cusped and has three roots. The paracone and metacone are larger than the protocone and hypocone. The cingulum is weak and present at the buccal side of the crown (Burchak-Abramovich and Gabashvili, 1945; Gabunia et al., 2001).
Distribution. Late Miocene (Upper Maeotian, MN 11–12?), Udabno locality, Sagaredgi district, eastern Georgia (Figure 1B).
Subfamily Colobinae Blyth, 1875
Mesopithecus Wagner, 1839
Mesopithecus ucrainicus Gremiatskiy, 1953
This taxon is represented by PIN 1997-4, a left maxillary fragment with P4–M3 (Gremiatskiy, 1957, 1961; Maschenko, 1989) (Figure 3). It is a small-sized species of Mesopithecus. The largest tooth is the M2. The M3 does not have a markedly reduced distal part of the crown, although its mesial transverse diameter is 25% larger than the distal transverse diameter. Relief of the molar crowns is moderately high. The P4 is bicuspid, and the cingulum is well developed on the distolingual surface of the crown. The length of the P4–M3 tooth row is 25.1 mm.
 View Details | Figure 3. Fragment of left maxilla of Mesopithecus ucrainicus from the Grebeniki 1 locality, Ukraine, Late Turolian. PIN 1997-4: (A) occlusal view; (B) lingual view. |
Distribution. Late Miocene (Late Turolian/Upper Maeotian, MN 13?), Grebeniki 1 locality, Velikomichailovskiy district, Odessa region, Ukraine (Figure 1A).
Subfamily Colobinae Blyth, 1875
Mesopithecus Wagner, 1839
Mesopithecus pentelicus Wagner, 1839
This taxon is represented by a male mandibular corpus with complete tooth row and missing ascending ramus (Heintz et al., 1981). This is the only find of Colobinae from the Miocene of Central Asia (collection number MOL 001, housed in the Paris Natural History Museum).
Distribution. Late Miocene (MN 12?), Molayan locality, central Afghanistan, Kabul province, Central Asia.
Mesopithecus sp. (Mesopithecus pentelicus?)
A complete male mandibular corpus, isolated teeth, lower jaw fragments, and limb bones of different individuals of Mesopithecus are known from Maragha in northern Iran. The mammalian fauna of Maragha is similar to the Pikermi fauna (de Mecquenem, 1925; Bernor et al., 1996). The male mandible is housed in the Natural History Museum, Paris, collection number MRG 1905-10; the location of the other specimens is unknown.
Distribution. Late Miocene (MN 12?), Maragha locality (Figure 1B), northern Iran (Iranian Azerbaijan), 50 km from the southeast coast of Lake Urmia.
From the Pliocene (Ruscinian) and Pleistocene of Russia (Transbaikalian province), northern Mongolia, Ukraine, Moldavia, and Tadjikistan, representatives of Cercopithecidae are known, including two genera and four? species of Cercopithecinae, and two genera and three species of Colobinae. From the territories of Russia and the adjacent countries, 11 localities of this age are known that have produced remains of cercopithecid monkeys. The chronological distribution of Cercopithecidae from these localities spans the end of the Early Pliocene (MN 14, 4.5 Ma) to the Middle Pleistocene (0.35 Ma) (Table 1). In Moldavia and Ukraine, cercopithecids make up part of the Moldavian (Ruscinian) and Chaprovian (Early Villafranchian) mammalian assemblages. One cercopithecid genus present in the Late Pliocene of Tadjikistan forms part of an Asian analogy of the Late Chaprovian (Middle Villafranchian) mammalian assemblage. Cercopithecidae from western Transbaikalian Russia were part of the Early–Middle Villafranchian mammal assemblages. The one and only representative of Cercopithecinae is known from the Middle Pleistocene of Georgia; genus Macaca, forming a part of the Caucasian analogy of the Cromerian (warm stage of Mindel or Interglacial IV).
In the Late Ruscinian of southern Moldavia and Ukraine, both subfamilies of Cercopithecidae coexisted: Colobinae (one genus and two species: Dolichopithecus ruscinensis and D. hypsilophus) and Cercopithecinae (one genus: Macaca, perhaps including two species). Cercopithecinae and Colobinae are known together from at least two localities dated to the latest Ruscinian of Ukraine: Novopetrovka and Budey (Figure 1A, Table 1). Beginning in the second half of the Late Pliocene (from MN 16), the subfamily Cercopithecinae is present only in the southern part of eastern Europe and in Tadjikistan. Stratigraphic distribution of the genus Dolichopithecus in southern Ukraine was most probably limited to between 2.2 and 2.4 Ma. From the Late Ruscinian(?)–Early Villafranchian of Ukraine, D. hypsilophus, a species smaller than D. ruscinensis, is known.
Subfamily Colobinae Blyth, 1875
Dolichopithecus Deperet, 1889
Dolichopithecus ruscinensis Deperet, 1889
This taxon is represented in the Early Pliocene by isolated lower M2 and P4, and a fragmentary right mandible (part of the mandibular corpus and base of the processus coronoides of a juvenile individual). The dp4 is preserved in the latter specimen from Novopetrovka, the permanent M1 of which is not yet erupted (Figure 4) (Alexeeva, 1964). Measurements of the specimen are: crown length of dp4, 9.4 mm; mandibular corpus depth at M1, 17 mm, corpus width at M1 18 mm. The specimen is housed in the Zoological Institute, Academy of Sciences of Ukraine, Kiev.
 View Details | Figure 4. Fragment of the mandible of Dolichopithecus ruscinensis from the Novopetrovka locality, Ukraine, Early Pliocene (from Alexeeva, 1964). (A) Buccal view; (B) occlusal view. |
A left M2 (M1?), GIN 541-133 from Kotlovina (Figure 5), is attributed to Dolichopithecus ruscinensis as the latest example of the genus in southeastern Europe. The associated mammalian fauna is Chaprovian (Early Villafranchian) (Alexeeva, 1977). The specimen was found by I.M. Gromov in 1964, and is housed in the Geological Institute, Russian Academy of Sciences, Moscow.
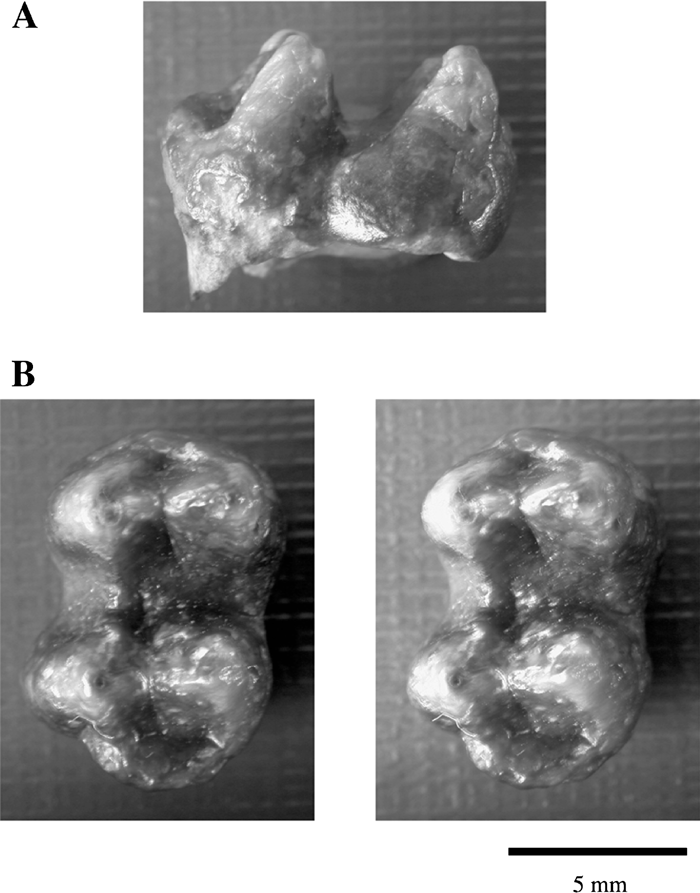 View Details | Figure 5. Left M2 of Dolichopithecus ruscinensis from the Kotlovina locality, Ukraine, Late Villafranchian. GIN 541-133: (A) buccal view; (B) occlusal view (stereo pair). |
Distribution. Early Pliocene, Ruscinian (MN 14–15?), Novopetrovka locality, Velikomichailovskiy district, Odessa region, Ukraine (Figure 1A). Late Pliocene, Late Villafranchian (MN 16?), the Kotlovina locality, Odessa region, southern Ukraine (Figure 1A).
Dolichopithecus hypsilophus (Gremiatskiy, 1958)
This taxon is represented by two lower jaw fragments from a locality near Voynichevo village (Kuchurgan River valley) (Gremiatskiy, 1958, 1961; Maschenko, 1989, 1991a). The specimens are housed in the Paleontological Institute, Russian Academy of Sciences: PIN 355-7 is a left mandibular corpus preserving fragmentary P3–M3; PIN 355-9 is a right mandibular corpus preserving the symphyseal part, lower canine, and P4–M3 (Figure 6). This is a middle-sized colobine species, smaller than D. ruscinensis. The length of the P3–M3 row is 47 mm. The very narrow symphysis is flattened. The incisors are aligned in a straight row and, judging by alveolar size, were very small. In the male D. hypsilophus, the canine is smaller than in the male of D. ruscinensis. Molar relief is not as deep as it is in D. ruscinensis. It does not differ significantly from D. ruscinensis in the structure of the C–P4, the morphology of the symphyseal part and the relatively small lower incisors. The main cusps of the molars are not as inclined mesially as they are in D. ruscinensis.
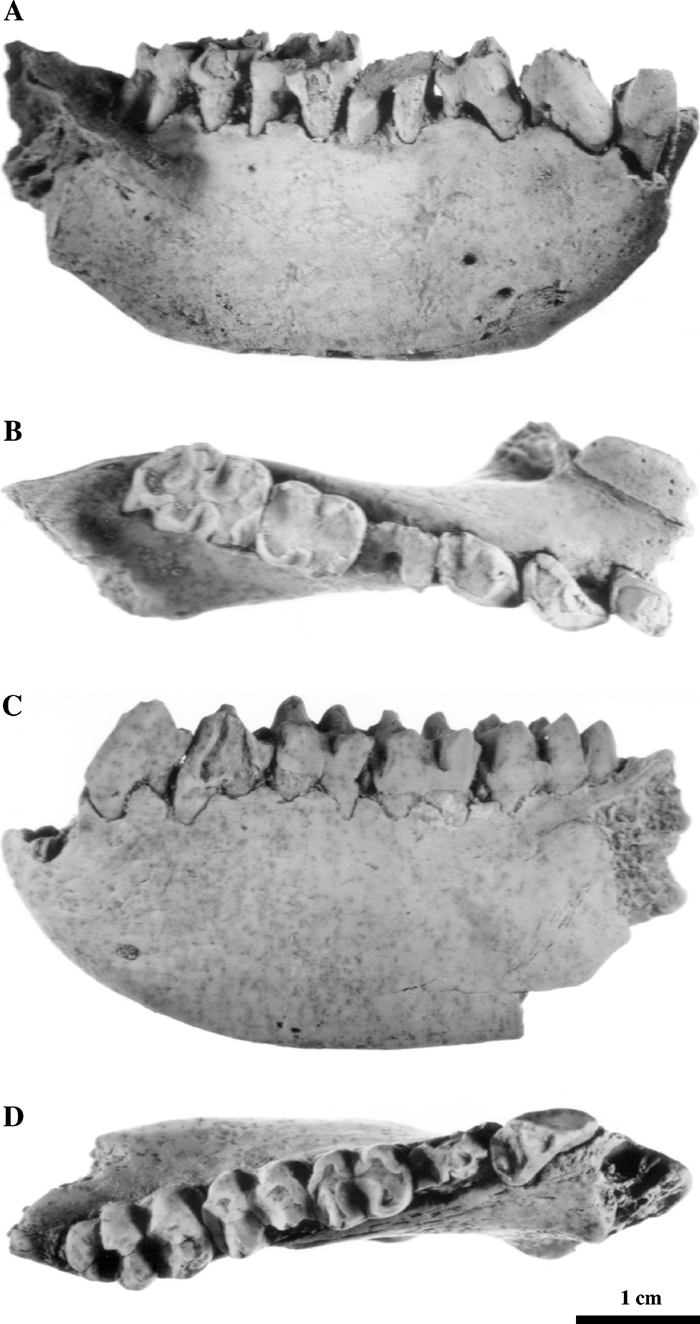 View Details | Figure 6. Mandibles of Dolichopithecus hypsilophus from the Voynichevo locality, Ukraine, the latest Ruscinian–Early Villafranchian(?) (MN 15). PIN 355-9: (A) buccal view; (B) occlusal view; PIN 355-7: (C) buccal view; (D) occlusal view. |
Distribution. Early Pliocene, latest Ruscinian–Early Villafranchian(?) (MN 15), the Voynichevo locality, Kuchurgan river, Odessa region, southern Ukraine (Figure 1A).
Dolichopithecus sp.
A fragmentary humerus (Figure 7) and a lower jaw fragment with P3–M3 lacking the ascending ramus from Budey are attributed to Dolicopithecus. Both specimens are housed in the Zoological Institute, Academy of Sciences of Moldavia, Kishinev (Alexeeva, 1964; David and Obade, 1995). Measurements of the humerus are: preserved length (lacking proximal and distal epiphyses), 182 mm; preserved distal width, 35 mm; minimal diaphyseal width, 16 mm.
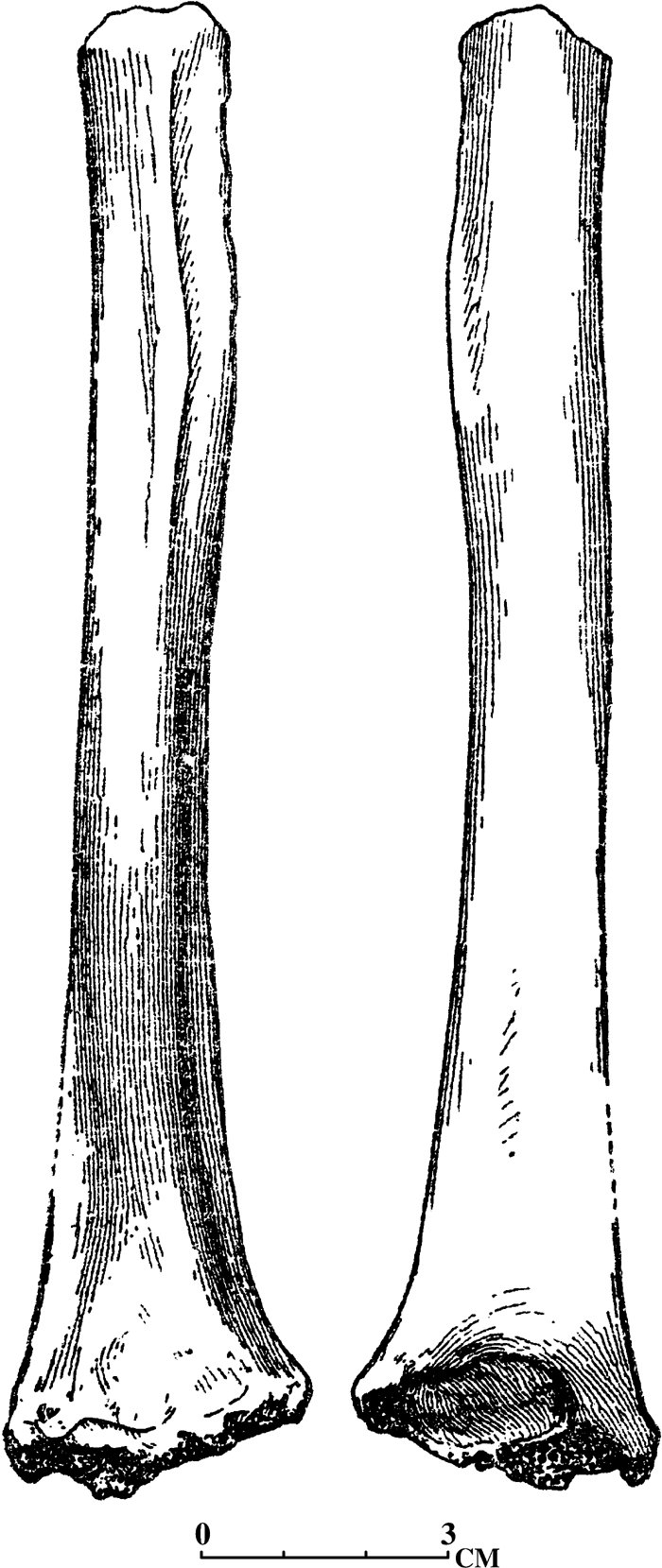 View Details | Figure 7. Fragment of left humerus Dolichopithecus ruscinensis (from Alexeeva, 1964) from the Budey locality, Moldavia, Early Pliocene. (A) Anterior view; (B) posterior view. |
Distribution. Early Pliocene, Late Ruscinian (MN 14–15), the Budey locality, right bank of the Bolshaia Salcha river, Porat district, southern Moldavia (Figure 1A).
Subfamily Cercopithecinae Gray, 1821
Macaca Lacepede, 1799
Macaca sp.
Macaca is represented by an isolated left upper M1 (GIN 1047/93) (Figure 8) (Tesakov and Maschenko, 1992) from the Grebeniki 2 locality, the first reliably identified find of the genus Macaca from the Pliocene of Ukraine. The locality is attributed to the end of the Early Ruscinian to the beginning the Late Ruscinian (MN 14/15). The main cusps of the molar are bunodont. There is an additional styloid cusp in the middle of the buccal surface. Mesial and distal transverse diameters of the crown are equal. The crown relief is similar to the cercopithecine condition, as well as the shape of the main cusps and their positions. In features such as the size of the mesial and distal flanks, shape and depth of the median-lingual and median-buccal notches and grooves, the Grebeniki 2 monkey may be assigned to the genus Macaca. The mesial transverse diameter is 6.5 mm; distal transverse diameter 6.5 mm; mesiodistal diameter 6.9 mm; length of lingual root 9.5 mm; buccal height of crown 4.8 mm.
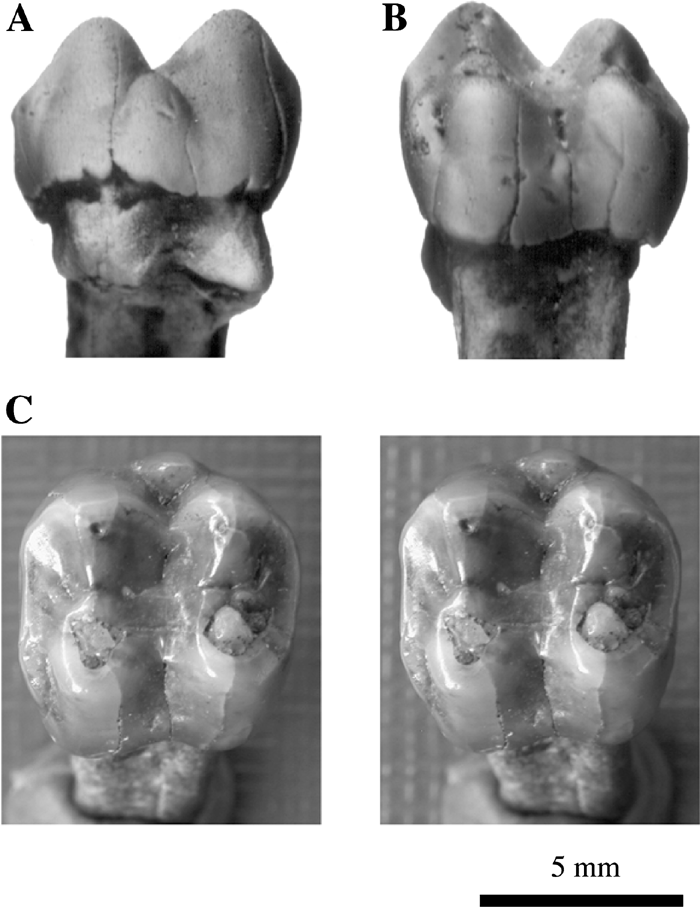 View Details | Figure 8. Isolated upper left M1 of Macaca sp. from the Grebeniki 2 locality, Ukraine, Late Ruscinian. GIN 1074/93: (A) buccal view; (B) lingual view; (C) occlusal view (stereo pair). |
Other specimens attributable to Macaca include a partial upper jaw with P3–M3 and base of the zygomatic process from the Gavanosa ravine, housed in the Zoological Institute, Academy of Sciences of Moldavia, Kishinev (David and Obade, 1995); and an isolated left male canine from Tcebricovo, Moldavia, housed in the Paleontological Institute, Russian Academy of Sciences, Moscow.
Distribution. Early Pliocene, Late Ruscinian (MN 14/15), the Grebeniki 2 locality, Velikomichailovskiy district, Odessa region, Ukraine (Figure 1A). Early Pliocene, Late Ruscinian (MN 14–15), the Gavanosa ravine locality, Bolshaia Salcha river, southern Moldavia (Figure 1A). Early Pliocene, Late Ruscinian (MN 15), the Tcebricovo locality, Kagul district, southern Moldavia (Figure 1A).
In Transbaikalian and Mongolia, from the boundary of the Early–Middle Pliocene, only Colobinae (Parapresbytis eohanuman) is present and Cercopithecinae is absent. The cercopithecines appear in China and Korea in the Middle Pleistocene. P. eohanuman is a representative of the ‘rhinopithecomorph’ colobine group, widely spread in Asia (Transbaikalian, Mongolia, China, Japan) from the end of the Middle Pliocene to the Early Pleistocene. It is a large-sized representative of Colobinae characterized by robust upper and lower incisors. Enamel on the labial surface of the incisors has well developed shovelling. The lingual surface of the incisors forms an enamel pocket with lateral enamel folds. The M3 has a non-reduced distal pair of main cusps. The dentition of P. eohanuman is rather close to that of modern Rhinopithecus roxellana and R. lantianensis from the Early Pleistocene of China.
Subfamily Colobinae Blyth, 1875
Parapresbytis Kalmykov and Maschenko, 1992
Parapresbytis eohanuman (Borissoglebskaya, 1981)
The colobine monkey from southern Transbaikalian (the Udunga locality, Buryat Republic) is the northernmost primate ever recorded in Eurasia. It is notably the only primate occurrence known from Siberia. Abundant material from the Udunga locality provided a basis for distinguishing a new colobine genus, Parapresbytis eohanuman (Kalmykov and Maschenko, 1992, 1995; Maschenko, 1994b). The specimens are housed in the Geological Institute, Siberian Branch of the Russian Academy of Sciences, Ulan-Ude. Specimens include GIN U 987/445, GIN U 987/493 (1), GIN U 987/878 (1), GIN U 987/878 (2), the upper jaws, and cranial fragment (Figure 9). Dating of the locality is disputable. Most probably, it is at the boundary of Late Ruscinian and Early Villafranchian (Kalmykov, 1992).
 View Details | Figure 9. Upper jaw and cranial fragments of Middle Pliocene Parapresbytis eohanuman from the Udunga locality, Transbaikalian, Russia. GIN U 987/445, right maxilla preserving P3–M3: (A) lingual view; (B) buccal view; (F) occlusal view. GIN U 987/878 (2), premaxilla preserving right I1 and left I1–2: (C) lingual view; (D) labial view. GIN U 987/878 (1), left premaxilla and maxilla preserving I1–P3: (E) lingual view; (G) labial view. GIN U 987/493 (1), cranial fragments: (H) dorsal view; (I) ventral view; (J) frontal view; (K) lateral view. |
The frontal part of the maxilla is broadened transversely and robust. The upper incisors are relatively large and inclined anteriorly. The labial enamel of the incisors forms vertical folds. The mesiodistal diameter of the incisors is less or equal to the buccolingual diameter. The mandibular incisors are inclined posteriorly. Lateral crests (folds) of enamel and enamel protuberance at the base of the lingual surface of the incisors form pockets. Parapresbytis differs from Dolichopithecus in the relatively minor development of crown relief of the molars, in C–P3 and C–P3 complex structures, and in the extremely wide and short lower facial part of the skull. The special adaptation of this Asian colobine is illustrated by the morphology of the incisors, canines, and premolars (robust and usually deeply worn in comparison with molars), and morphology of the frontal part of the maxilla. These features differentiate Parapresbytis from Rhinopithecus as well as from other Asian colobines.
Two lower jaws (PIN 3381-236, 3381-235) and fragments of the limb, a humerus, and an ulna (PIN 3381-210, 3381-211) from Shamar, northern Mongolia are also attributable to P. eohanuman (Figure 10). These specimens, found in 1975, were later described as Presbytis eohanuman Borissoglebskaya, 1981 (Borissoglebskaya, 1981). Re-examination of the Shamar material shows that the Transbaikalian colobine bears similarity with recent Rhinopithecus. Parapresbytis is larger in size than recent colobines. The characters of the trochlea of the humerus are indicative of an adaptation to arboreal life rather than to terrestriality (Figure 10). These specimens are housed in the Paleontological Institute, Russian Academy of Sciences, Moscow.
 View Details | Figure 10. Lower jaws and two limb bone fragments of Parapresbytis eohanuman from Shamar Locality, northern Mongolia (from Borissoglebskaya, 1981). PIN 3381–236, nearly complete mandibular corpus preserving right and left I1–M3: (A) occlusal view; (B) buccal view. PIN 3381-235, mandible: (C) occlusal view. PIN 3381–210, distal end of left humerus, and PIN 3381-211, right ulna: (D) humerus anterior view; (E) humerus posterior view. (F) ulna lateral view; (G) ulna anterior view. |
Distribution. Middle Pliocene, latest Ruscinian(?)–Early Villafranchian (MN 16?), the Udunga locality, southern Transbaikalian, Russia. Late Pliocene, Early Villafranchian (MN 16), the Shamar locality, northern Mongolia.
In the Late Pliocene–Early Pleistocene of Ukraine and Moldavia the genus Paradolichopithecus is not known. In the Late Pliocene of Central Asia (Tadjikistan, Kuruk-Say) a representative of the genus Papio is reliably identified. Papio (Paradolichopithecus) sushkini is a specialized representative of the genus. It is medium-sized and differs from other extinct and modern Papio in large bunodont molar teeth and thick enamel. The material from Tadjikistan testifies that Papio was not endemic to Africa, and that in the end of the Pliocene to the beginning of the Pleistocene the genus was wide-spread in Asia. The time of dispersal of Papio coincided or was almost synchronous with the adaptive radiation of the genus into areas of its basic distribution. Most likely, the Late Pliocene (Early–Middle Villafranchian) was a time of maximal distribution of the genus in the Old World.
Subfamily Cercopithecinae Gray, 1821
Papio Muller, 1773
Papio sushkini (Trofimov, 1977)
This taxon is represented by crania, lower jaws, and isolated teeth from Kuruk-Say, Tadjikistan. The specimens are housed in the Paleontological Institute, Russian Academy of Sciences, Moscow (Trofimov, 1977; Maschenko, 1989, 1991b, 1994a). Specimens include the holotype female skull, PIN 3120-523 (Figure 11A, B); a fragment of the maxilla of a subadult male, PIN 3120-524; the lower jaw of an adult male, PIN 3120-526 (Figure 11C, D); the lower jaw of an adult male, PIN 3120-525 (Figure 11E, F); a fragment of the skull of a subadult individual, PIN 3120-524; isolated P3 and M3, PIN 3120-707; isolated I1 and I2, PIN 3120-708, 709.
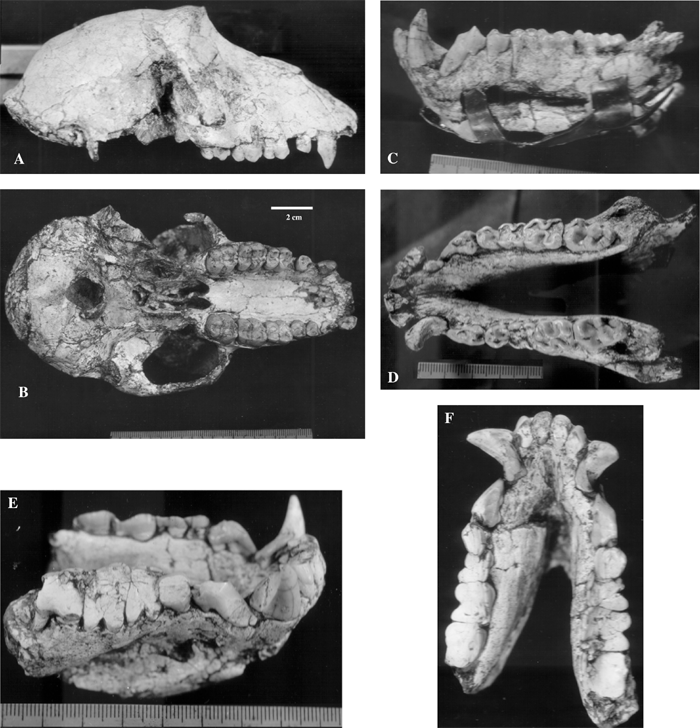 View Details | Figure 11. Skull and lower jaws of the Middle Pleistocene Asian baboon Papio sushkini from the Kuruk-Say locality, southern Tadjikistan. PIN 3120-523, female skull: (A) lateral view; (B) occlusal view. PIN 3120-525, male mandible: (C) lateral view; (D) occlusal view. PIN 3120-526, male mandible: (E) lateral view; (F) occlusal view (E and F are in the same scale). |
Papio sushkini is a medium-sized extinct specialized representative of Papio. It is similar to recent Papio in morphology of the infraorbital foramina, ratio of lengths of the facial and cranial parts of the skull, the short and robust alveolar process of the upper jaw, the large anteroposterior diameter of foramen magnum, the shape of the nasal bones, position of the fossa glandula lacrimalis (only on os lacrimale), shape of the cecal foramen, morphology of the CP3 complex, molar morphology, and relation of lengths of the premolar and molar tooth rows. Specialization of Papio sushkini in comparison with modern representatives of the genus is found in a greater robustness of molars relative to premolars, greater enamel thickness, greater length of the zygomatic arch relative to total length of the skull, and a more robust M3 in comparison with M2.
Distribution. Late Pliocene, Late Villafranchian (MN 16), the Kuruk-Say locality, southern Tadjikistan, Central Asia.
Subfamily Cercopithecinae Gray, 1821
Macaca Lacepede, 1799
Macaca ex gr. anderssoni–arctoides
A macaque is known from the Middle Pleistocene Kudaro 1 locality (Georgia), represented by isolated teeth, astragalus (Figure 12), and a phalanx of finger 3 or 4. All macaque remains come from the lower part of cultural layer 5, which is formed by yellow-brown loam. Layer 5 contains stone artefacts typologically referred to as Acheulean. Layer 5c (the southern gallery of the Kudaro cave) was dated by the radiothermoluminescence method to 360 ka BP; and layer 5b was dated to 350 ka BP. Thus, layers 5b and 5c can be referred to the warm phase of the Mindel interglacial epoch (interstadial IV of the Cromerian complex) (Vereshchagin, 1959; Vereshchagin and Lubin, 1960; Delson, 1980; Vereshchagin and Baryshnikov, 1994). Molar morphology is closer to the Asian Macaca condition of the ‘sinica’ species group than to that of the European representatives of the genus Macaca (Maschenko and Baryshnikov, 2002). Structural features of the astragalar trochlea and the proportions and orientation of the head relative to the axis of the body show a great similarity mainly to terrestrial species. The macaque from Kudaro is larger than extant M. nemestrina. Among extinct species, it is smaller than only M. anderssoni from the Middle Pleistocene of China.
 View Details | Figure 12. Middle Pleistocene M. ex gr. anderssoni–arctoides from Kudaro 1 Cave, northern Osetia, Georgia. ZIN 34600. right M2: (A) lingual view; (B) buccal view; (C) occlusal view. ZIN 34605, astragalus: (D) dorsal view. |
An important common dental feature of Asian macaques and the form from Kudaro is the bottom of the mesial and distal foveae, which are level with each other while the bottom of the trigon basin is positioned lower. In some species from Asia and in M. sylvanus, the bottom of the mesial fovea is positioned higher than with the two other basins. Furthermore, in the macaque from Kudaro, and in extant M. nemestrina and M. fuscata and extinct M. anderssoni from China, the difference of the mesial and distal transverse diameters is relatively small, and the size difference between M2 and M3 is minimal. In these features, the Kudaro form shows a greater similarity to extant Asian macaques than to Pleistocene M. sylvanus. This testifies to its affinity with Asian Macaca species. Tooth morphology allows us to propose that the sinica group is closer to the Kudaro form than the other Asian groups; and within this group, the closest species are M. arctoides and M. anderssoni. Thus, the most likely identification of the macaque from Kudaro is M. ex gr. anderssoni–arctoides.
Distribution. Middle Pleistocene, Mindel interglacial epoch (interstadial IV of the Cromerian complex), the Kudaro 1 locality, northern Osetia, Georgia (Figure 1B).
In the Cenozoic, the eastern Eurasian area under consideration in the present paper was not the center of origin of a large number of primate taxonomic groups. The western area was influenced zoogeographically by the Mediterranean; the eastern area was in contact with southern Asia, and sometime in the Pliocene, with Africa. This explains the abundance of either specialized, or widely distributed, forms of primates found in the Eurasian faunas. The total number of known primate taxa is not great. From the Early Eocene to Pleistocene there are three families (Omomyidae, Hominidae, Cercopithecidae), seven genera (Altanius, Dryopithecus, Mesopithecus, Dolichopithecus, Papio, Parapresbytis, Macaca), and eleven species.
During the Late Miocene and Early Pliocene, owing to links between the Mediterranean and African provinces, representatives of both subfamilies of Cercopithecidae came to inhabit the geographic area reviewed in this paper. From the point of view of general modifications of mammalian fauna compositions, the appearance of Omomyidae is due to faunal exchange between Asia and North America. Other lower primates never existed in this area. In the end of the Miocene the extinction of representatives of Hominoidea coincides in time with the distribution of the Hipparion faunas. This may be due to global climatic change (establishment of seasonal climate) in the end of the Miocene and distribution of open landscapes (disappearance of rainforests). As in western Europe, at the western part of the area considered in this review, the diversity of Cercopithecidae increased by the end of the Miocene, overwhelming the diversity of Hominoidea. At the end of the Pliocene to the beginning of the Pleistocene, Cercopithecinae came to substitute Colobinae. From this time onwards, the latter occurred only in the eastern part of the area considered in this review, and later retreated to southern Asia. Extinction of the few representatives of the subfamily Cercopithecinae in this area during the Pleistocene is connected with the general fall of temperature in the northern hemisphere. During the Pleistocene, however, some species of Macaca existed in the southernmost areas, in a relatively more optimal environment. During the Late Pleistocene, primates became extinct, and in the Holocene they reappeared again in certain regions of Asia (e.g. M. arctoides in Afghanistan).
Further studies of primates from the Eurasian area considered here will probably yield new information on the systematic diversity of primates in the Neogene and cast new light on some of the peculiarities of evolution of the several groups. Most probably new forms of the family Cercopithecidae are to be discovered; and the diversity of Hominoidea may have been higher than the available data suggests.
The author is grateful to two anonymous reviewers for their valuable comments which greatly improved the paper and to editors Dr M. Takai and Dr M. Huffman for their careful work with the manuscript. The author’s research was partly supported by the President of Russian Federation for outstanding Scientific Schools #1840.2003.4. The author is grateful to Dr Alexander N. Kharitonov (Psychological Institute RAS) for help in translation of the manuscript from Russian into English.
|