| Greg C. Nelson, corresponding author. e-mail: gcnelson@uoregon.edu phone: +1-541-688-2906; fax: +1-541-346-0668 Published online 29 June 2005 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.040710 |
Remote Oceania was the last major geographic area occupied by Homo sapiens as it grew into the worldwide species we know today. This colonization, which occurred within the last 4500 years or so, has left numerous tantalizing clues as to its origins, direction, and timing. That pottery-using, Austronesian-speaking peoples probably expanded out of Taiwan, southward through the Philippines and into Near Oceania (where the distinctive Lapita complex developed) and thence into Remote Oceania, is supported on both archaeological and linguistic grounds (Spriggs, 1995, 1997; Kirch, 1997).
Lapita peoples first appeared in the Bismark archipelago around 3500 BP, moved rapidly south and east beginning about 3200 BP, and reached islands in western Polynesia within about 300 years (Kirch, 2000). However, occupations of Palau and the Marianas were contemporaneous with Lapita expansion into the rest of Remote Oceania. The earliest acceptable dates in western Micronesia cluster around 3500 BP (Spoehr, 1957; Butler, 1994) and provide an avenue for deciphering the settlement patterns of linguistically similar, yet vastly separated Oceanic peoples.
Archaeological and linguistic evidence seems to suggest that the colonization of islands in western Micronesia predates the appearance of the Lapita complex. Pottery from both Palau and the Marianas is similar to early red-slipped ware in the northern Philippines, and both the Palaun and Chomorro languages belong to the western Malayo-Polynesian subgroup of Austronesian, a more ancient branch of the language than the Oceanic branch associated with Lapita (Kirch, 2000). Paleoenvironmental evidence from Guam (Athens and Ward, 1995) and Palau (Ward et al., 1998, cited in Kirch, 2000) may suggest an even earlier human presence, perhaps as early as ca. 4000 BP. Although not definitive, these clues emphasize the importance of investigating prehistoric habitation of Palau, since it could record the earliest human occupation of Remote Oceania. As the westernmost island group in Micronesia, Palau is the closest to insular Southeast Asia. Thus, it is a logical location for pre-Lapita Austronesian speakers to have occupied on their migration southward from Taiwan as it could be reached from Guam in the north or from the Philippines to the west.
Despite proxy evidence of a second millennium BC settlement of Palau, the earliest acceptable dates hover between 3000 and 3300 BP. Unfortunately there is little skeletal evidence of these early inhabitants that would help answer questions about their biological makeup, pathologies, and mortuary behaviors. Genetic studies (Lum and Cann, 1998, 2000) have only cursorily been corroborated with human remains (Pietrusewsky, 1985, 1996; Kirch et al., 1989; Hanson and Pietrusewsky, 1997). This lack of a large sample representing inhabitants from an early population remains a major roadblock in deciphering the history of population movements into and within the Pacific. Recent discovery of a large, early, cemetery in Palau in western Micronesia has the potential to fill many gaps in our knowledge of the earliest peoples of this region.
The Chelechol ra Orrak site (B:IR-1:23) is located on Orrak Island just off the southeast tip of the volcanic island of Babeldaob, the largest in the Palauan archipelago (Figure 1). It is one of several sites explored as part of the Palau Stone Money Project, a joint undertaking by one of the authors (S.M.F) and the Palau Bureau of Arts and Culture (BAC, formerly the Division of Cultural Affairs) involved in investigating the manufacture and exchange of Yapese stone money disks (Fitzpatrick, 2001, 2003a). The island is limestone in composition and connected to Babeldaob by a prehistoric causeway constructed of coral rubble now covered in mangrove vegetation.
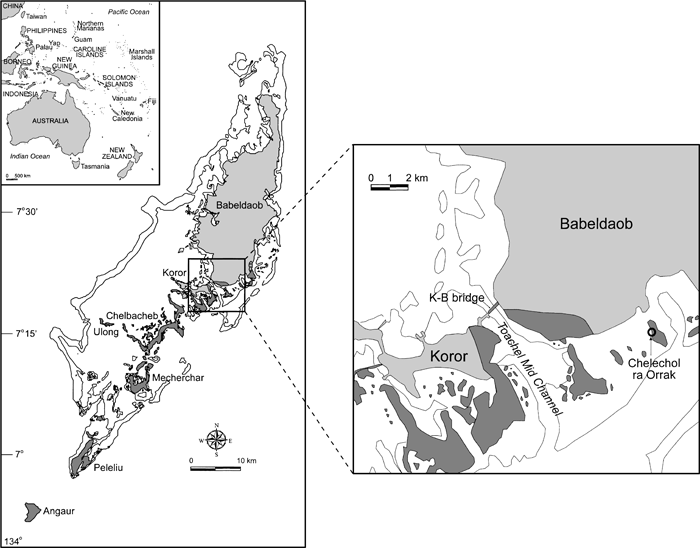 View Details | Figure 1. Map of the main Palauan archipelago (darker shading indicates limestone rock). |
Excavations at Orrak in August 2000 uncovered human skeletal remains at depths generally greater than 60–70 cm. Radiocarbon dates from human bone, charcoal, and shell returned acceptable ages ranging from ca. 2000 to 3000 calBP, making Orrak the earliest human burial site in the Pacific outside of Melanesia (Fitzpatrick, 2002a, 2003b). Although isolated human remains associated with Lapita occupations (2500–3500 BP) have been reported (Pietrusewsky, 1985; Kirch et al., 1989) these are fragmentary and few in number. In Micronesia, outside of large precontact cemeteries on Guam generally associated with the pre-Latte and Latte periods dating to the last 2000 years (Douglas et al., 1997; Hanson and Butler, 1997), skeletal remains of early inhabitants are few. The Chelechol ra Orrak site, with approximately 25 individuals represented, is the only large cemetery site dating in the 2000–3000 BP range found in Micronesia.
The Chelechol ra Orrak (B:IR-1:23) site was originally identified as a Yapese stone money quarry by Blaiyok (1993) and consists of a large rockshelter infused with several smaller caves and crevices located at the head of a small tidal inlet on the western margin of Orrak island. At present, extensive vegetation and coral rock features built by Yapese Islanders for quarrying stone money (Fitzpatrick, 2002b) cover the inlet and the beach immediately outside the rockshelter and obscure the site from view. The beach, extending in a northwest to southeast direction from the inlet, is one of three main beaches on the island and is 2–8 m wide depending on the tide level. Archaeological investigation at Orrak initially concentrated on collecting data on Yapese stone money production and focused in the center portion of the rockshelter about 10 m from an unfinished stone money disk.
Four test units were opened (two are 1.0×1.0 m; two are 1.0×0.5 m) (Figure 2); three were excavated to 90 cm below surface or more and all contained human remains. Soils in the upper 50 cm were typically a mixture of calcareous sand and silty loam intermixed with limestone and coral rock, shellfish, fish bone, and a plethora of artifacts. Soils below 50 cm were mostly calcareous sand deposits with spotty loam inclusions with a decrease in faunal remains and artifacts. Sediments were water screened through 1/8-inch mesh to ensure good recovery of smaller cultural remains. An extensive faunal assemblage was collected representing elasmobranchs, Crustacea, turtle, sea urchin, small amounts of mammal and bird bone, nearly a hundred different shellfish taxa, and over 20 teleost fishes (Fitzpatrick and Kataoka, 2005). Artifacts found included glass beads, pearl shell (Pinctada margaretifera) scrapers/graters (Fitzpatrick and Boyle, 2002), Tridacna sp. adzes, a stone adze, shell ornaments (e.g. Trochus sp. rings, Conus sp. shell beads, disks, and pendants), pottery, drilled turtle and fish bone, and a bone needle.
 View Details | Figure 2. Site map of Chelechol ra Orrak with inset of Orrak Island. Test Units 1–4 (shaded) were excavated in year 2000. Test Units E2S1, E2S2, and E3S1 were excavated in 2002 to a depth of approximately 50 cm. |
To determine the antiquity of burials and subsequent deposits, samples of bone, charcoal, and shell were submitted to two different labs for AMS radiocarbon dating—the National Ocean Sciences Atomic Mass Spectrometer (NOSAMS) Facility at Woods Hole in Massachusetts and the University of Arizona AMS facility. All samples were calibrated using CALIB 4.3 after Stuiver and Reimer (1993; see Fitzpatrick, 2003b for detailed discussion of the dates).
Specimens for dating were collected from each test unit and nearly every stratigraphic layer down to depths of up to 110 cmbs. A majority of the dates (12 of 19) were from Test Unit 1 with ages ranging from the historic period in the upper layers to around 3000 calBP in the lower burial deposits. Six other dates from Test Units 2–4 ranged from 2340 to 3840 calBP. These dates indicate that burial activity at Chelechol ra Orrak took place from approximately 1800 to 3000 calBP, and possibly earlier, similar to what was found in Test Unit 1. All of the dates suggest that early burial activity was not restricted to a small area, but was fairly widespread throughout the rockshelter. It should be noted that the earliest dates (pre-3000 BP) are all from human bone and it is unclear whether diagenic effects have impacted preservation or if these dates are in fact reliable and simply represent older interments that have become mixed with later deposits through bioturbation or subsequent burial activity (Fitzpatrick, 2003b). Nonetheless, the radiocarbon suite suggests the antiquity of burials at the site extend back at least 3000 years and is supported by multiple dates on different sample material.
Upon completion of fieldwork in September 2000, permission was received from the BAC to transport the recovered human skeletal remains to the University of Oregon for study. Inventory of the specimens began in December 2000 and continued through January 2001. Reconstruction and analysis then proceeded through summer 2001. Additional material was received in September 2001 and integrated into the collection.
Of the four test units excavated during the 2000 field season, Test Units 1 and 4 produced the vast majority of human skeletal remains. These remains are apparently from both primary and secondary burial contexts, although the skeletal element profiles of the units are quite different. Located at the back of the cave near the wall, the skeletal assemblage of TU-1 (see Figure 2) could best be described as a mix of unassociated fragments representing many individuals plus two sets of articulated legs in the lowest levels. In contrast, remains from Test Unit 4 (2 m distant to the southwest) consist of five disarticulated, though more complete, adult individuals. Test Units 2 and 3 have a few isolated remains each. Preservation of the bone spans the continuum from excellent to very poor and fragile. In general, material from inside the drip line (Test Units 1 and 2) is in better condition, perhaps due to greater protection from storm surges, tidal action, and rainfall. Determination of minimum number of individuals (MNI) was made through analysis and correlation of dental and skeletal element count, dental wear, skeletal and dental age, and element size. Age was estimated through standard assessment practices including dental eruption and wear, long bone growth, epiphyseal union, and cranial suture closure (Ubelaker, 1989; White, 1991). Although sexing was difficult due to the fragmentary nature of the remains and the lack of complete innominates, several sciatic notches are preserved (all from Test Unit 4) and this, combined with cranial robusticity and long bone size and robusticity, allowed an estimate of the number of males and females represented.
Skeletal metrics were recorded for all complete and reconstructed elements available for measurement. Long bone lengths were recorded with an osteometric board. Long bone shaft and articular surface measurements and the length of metacarpals and metatarsals, were recorded with a digital Mitutoyo sliding caliper calibrated to 0.01 mm. Measurement protocols follow those outlined in Buikstra and Ubelaker (1994) and Bass (1995).
At the request of the Palau BAC, individual elements have not been numbered and are identified only by unit and level. Table 1 reports the MNI and the estimated breakdown by age and sex.
 |
The fragmentary nature of the majority of the remains makes it difficult to build a profile for the Orrak sample. Metrics for the few elements complete enough to allow some measurements are presented in Table 2, Table 3, Table 4, Table 5, Table 6, and Table 7. As it can be seen, the available measurable specimens consist only of two crania and 16 postcranial elements. Unfortunately, the incompleteness of the crania precludes comparative craniometrics being implemented at this time.





Description of the skeletal biology of the Orrak sample is by excavation unit. Only Test Units 3 and 4 are contiguous but the number and quality of the skeletal elements recovered from each is quite different, warranting separate treatment. The information presented here includes a detailed inventory of skeletal material present and anatomical description of important elements.
Human skeletal remains found in TU-1 represent an MNI of 13 ranging in age from prenatal to adult. There are approximately 350 identifiable (as human) bone fragments attributed to this unit, and these can be separated into two basic groupings. The first, comprising the majority of the bone, consists of unassociated fragments representing most skeletal elements with the largest being the distal two-thirds of a humerus. The second comprises portions of two sets of legs found in anatomical position at the bottom of the unit, one each in Layers 8 and 9, which probably represent portions of two internments (Figure 3). The legs from Layer 9 were collected while those from Layer 8 were carefully reburied.
 View Details | Figure 3. Test Unit 1 facing east. Note the two sets of articulated legs in the corners. |
Among the fragments belonging to the first group are elements representing five juveniles; prenatal, neonatal, 2–3 years old, 5 years old, and 10 years old, although the majority of skeletal elements recovered from TU-1 belong to adult individuals. The MNI of eight adults is based on the presence of at least two individuals in Layer 7 and three individuals in each of Layers 8 and 9 (isolated teeth from Layers 1 and 6 were not considered in MNI tabulation) broken down in the following manner. Layer 7, MNI is based on the differential staining of teeth, as a result of betel nut (Areca catechu) chewing (Fitzpatrick et al., 2003); Layer 8, cranial, long bone, and carpal elements each indicate a minimum of three individuals; Layer 9, articulated legs plus postcranial remains of two other individuals.
The human skeletal remains recovered from TU-2 consist of 14 postcranial fragments and three teeth. Overall, the preservation is moderate with the only complete element being a permanent upper left third molar. An MNI of two is based on the identification of one child (right dc) and at least one adult (left M3, fragmentary upper C). There is no duplication of elements within the postcranial assemblage.
Four teeth and 22 skeletal fragments including 11 adult cranial fragments, three juvenile cranial fragments, the body of a hyoid, one complete scaphoid, and the body of one thoracic vertebra (T11 or T12) were recovered from TU-3. By element count the MNI is two—one adult and one juvenile. However, although there is no duplication among the teeth (right I1, left M2, right M2, and right M3) the upper molars are from different individuals as there is no distal interproximal wear facet on the right M2 while there is a mesial facet on the right M3. This last indicates the presence of at least two adults and an MNI of three for the unit.
The human skeletal remains from TU-4 consist of cranial and postcranial elements representing at least five adults and two juveniles including portions of the lower legs of one prenatal individual. Preservation is poor overall and the adult postcranial bones in particular are fragile and fragmentary. In this unit three skulls were found grouped together and not in anatomical association with any of the postcranial elements. For TU-4, the adult MNI was calculated on the basis of these skulls and the four female and one male innominate recovered. When discovered, two of the crania, Skulls 2 and 4, were largely complete (Figure 4). However, as waterlogged bone is very soft, pliable, and highly susceptible to breakage, their condition soon deteriorated. The fragments were carefully packaged for transport but were in need of extensive reconstruction after reaching the lab. During inventory and reconstruction it was found that elements labeled in the field as belonging to two individuals, Skulls 1 and 3, in fact represented only one.
 View Details | Figure 4. Test Unit 4 with Skull 4 in situ. |
Skull 1 is a fragmentary calotte consisting of the right and left parietals with an attached fragment of the left zygomatic. Upon reconstruction, Skull 2 (Figure 5) presents most of the calvaria minus the foramen magnum (occipital condyles present but detached) and the bodies of the ethmoid and sphenoid. The parietals are complete except for an approximately 1 cm2 area on the left at the squamosal suture and a similarly sized gap in the central portion of the right. On the frontal, the area around glabella is broken away exposing the sinuses, approximately one-fourth of the outer table has spalled off, and several small pieces are missing throughout the main body of the bone. Portions of both greater wings of the sphenoid remain attached to the temporals, each of which is missing the superior portions of the squama. The mastoids are relatively small and the neurocranium lightly built, possibly indicating that Skull 2 is that of a female.
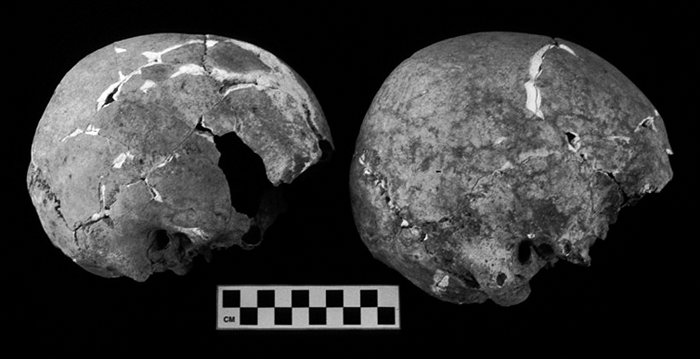 View Details | Figure 5. Skull 2 (left) and Skull 4 (right) reconstructed. Right norma lateralis. |
The most complete of the skulls is Skull 4 (Figure 5). The only elements absent are the ethmoid, body of the sphenoid, lacrimals, vomer, and conchae. Most of the maxilla is present but is too fragmentary and fragile to reconstruct. Though several fragments are missing we were able to partially reconstruct the mandible. The frontal is broken in the area of glabella (glabella is present but detached) exposing the sinuses. Although the zygomatics are present, the fragmentary nature of the anterior portion of the frontal precludes reconstruction of the orbits. All teeth except for the upper third molars are present and in good condition. That all teeth had erupted, but are in a relatively unworn condition, indicates that this individual was in his early 20s at death. Due to the general robusticity of the skull, the relatively large size of the mastoid processes, and the robusticity of the mandible, the sex is estimated to be male. The majority of the remaining cranial bones are fragile but complete except for the mastoid and petrous portion areas of the left temporal. In the posterior region around the area of the left occipitomastoid and superior into the lambdoidal suture there is very slight plastic deformation. No pathology was observed. However, a large intrasutural, or Inca, bone is located along the lambdoidal suture and comprises the superior third of the occipital bone.
Other human skeletal remains from TU-4 include six isolated teeth, five adult and one child, representing an MNI of three. Postcranial bones include fragments of all elements, although no whole bones remain except for patellae, carpals, tarsals, metacarpals, metatarsals, and phalanges.
Considering the overall bone preservation at Orrak it is surprising how many specimens derive from juveniles. Forty-three skeletal elements or fragments can be attributed to individuals aged approximately 10 years or less. Remarkably, three of those represent prenatal or neonatal individuals. One very small right tibia, minus epiphyses, from TU-1 is 44.2 mm long, which correlates to an in utero age of 7–8 months (Ubelaker, 1989). Also deriving from TU-1 is a neonate represented by four elements: a distal half of a right femur, a humeral shaft fragment, a small rib fragment, and a piece of frontal that retains a portion of the right orbit and the fronto-malar suture. The third comes from TU-4 and is represented by the right and left tibial diaphyses, a fibula diaphysis (unsided) missing the distal end, one diaphysis of a metatarsal or metacarpal, and five small unidentified fragments (Figure 6). The length of the right tibia is 46.96 mm, placing the in utero age at about 8 months (Ubelaker, 1989).
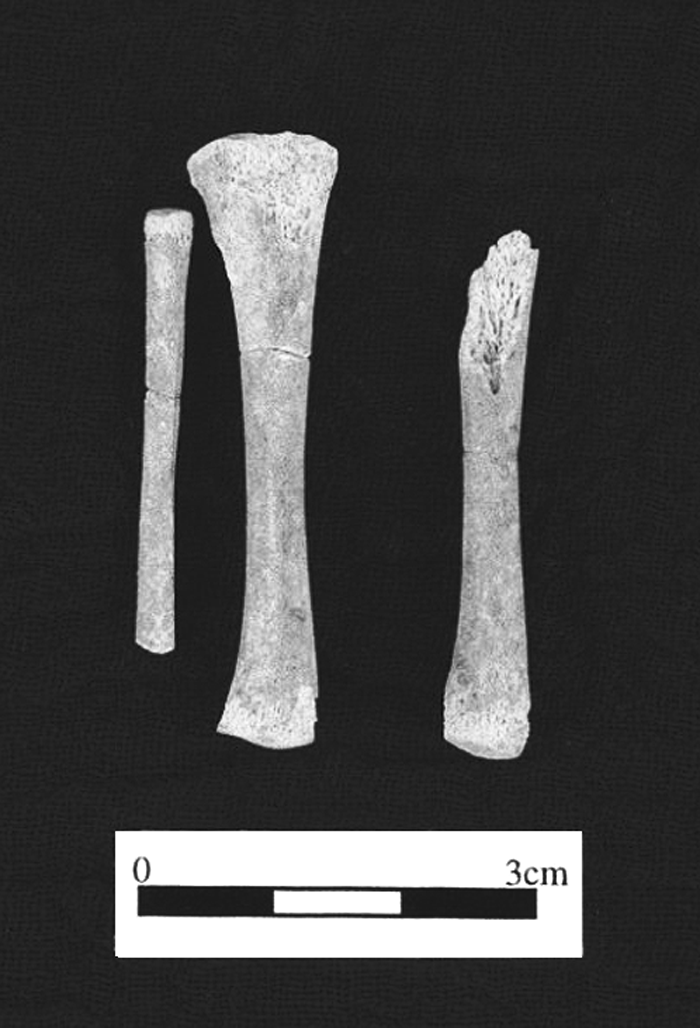 View Details | Figure 6. Right tibia and fibula and left tibia from prenatal individual from TU-4. |
The remainder of the juvenile remains from TU-1 include a 2–4 year old, a 5+ year old, and one of approximately 10 years of age. The 2–4 year old is represented by five fragments, most notably a portion of the left maxilla. Anteriorly, the maxilla is broken through the socket of left di1 and the crypt of left I1 and posteriorly just posterior of the socket of left dm2. In addition, the labial portion of the sockets for the left dc, left dm1, and left dm2 are broken away. The crypt for the left P3 is empty. The left dm1 fits in the partial socket and left I1 fits in the crypt. The left upper C is encrypted as is the left I2. Portions of the clivus and left nasal process are also retained.
Twelve fragments can be attributed to the 5+ year old including four cranial fragments, seven rib fragments, and a well preserved humeral diaphysis. Elements ascribed to an individual of approximately 10 years include a left mandibular corpus, three cranial fragments, a radius fragment, and a carpal phalange missing the proximal epiphyseal plate. The mandibular corpus extends from the mesial socket wall of the right C through the midpoint of the crypt for the left M2. The left dm2 and left M1 remain in place with the left M1 fully occluded. An isolated right I2 which fits in the right I2 socket is very slightly worn. The left P3 is visible in its crypt although the form of the socket for the overlying ldm1 indicates that this tooth was probably still present.
The final two juveniles are represented by teeth only. One right dc from TU-2 has a complete root with no indication that resorption had begun. An left di1 from TU-4 also has a complete root with wear just exposing the dentine.
Individuals aged 10 years or less comprise 36 percent (9/25) of the sample from Orrak. This compares with rates of 25.7 percent (39/152) for Apurguan, Guam (Douglas et al., 1997) and 37 percent (10/27) from Rota (Hanson, 1990). These seem to be high frequencies given that children are generally underrepresented in skeletal series due to factors such as poor preservation of their small, fragile bones, and differential burial practices. Considering the overall poor condition of the remains found at Orrak, it does seem anomalous that there are so many children represented, particularly neonatal and prenatal individuals. Although they could belong to very small full-term newborns, the tibia from TU-1 and the tibiae from TU-4 are so small that they probably represent premature deliveries or the death of pregnant females. At 44.2 mm long, the tibia from TU-1 is nearly 30 mm shorter than the 71.6 mm average cited by Ubelaker (1989) for tibial lengths of individuals aged newborn to six months. Using standards developed by Fazekas and Kosa (1978, cited in Ubelaker, 1989) the TU-1 tibia translates to a fetal length of 36.9 cm and a fetal age of approximately seven months. The right tibia from TU-4 (49.6 mm) indicates a fetus of 40.8 cm in length and a fetal age of eight months. Even considering that the research of Fazekas and Kosa (1978) is not based on a Pacific Island population and that each population has its own characteristic growth and development rates, these specimens, particularly that from TU-1, are so small and developmentally immature that it is doubtful they were full-term births.
The TU-1 fetus is not associated with other skeletal material complete enough to determine its burial situation. However, the fetus from TU-4 is found among a group of at least four adult females. Based on the stage of iliac crest fusion, one of these females was in her late teens at the time of death, and two of the others appear to be of childbearing age. Therefore, it is possible that the fetus is associated with one of these females. Since we do not know the circumstances of the deaths and burial of these individuals an association is unlikely unless DNA can be retrieved and comparisons made. Unfortunately the condition of the bone makes this improbable. Further excavation planned for Chelechol ra Orrak will hopefully help answer many of these questions.
Skeletal elements from TU-1 and -4 (no pathology was noted in the few fragments recovered from TU-2 and -3) exhibit examples of several pathological conditions and hint at some of the problems encountered by early inhabitants of Palau. Unfortunately, the fragmentary and disarticulated nature of the specimens from Orrak present a series of problems in interpreting the pathology found among the skeletal remains. Foremost is that in TU-4 no crania were found in association with more than a few cervical vertebrae so, although five adults appear to be present, no ‘individuals’, as the term is commonly construed, can be analyzed. However, within the postcranial remains a few associations between elements are possible since some bones, particularly hands, were found in anatomical association. Among the associated elements are several that exhibit the same extreme level of bony degeneration and appear to belong to a single osteoporotic individual. Common among skeletal pathologies found at Orrak is degenerative joint disease (DJD) with examples from both TU-1 and TU-4. In addition, single examples of porotic hyperostosis and periostitis are noted as well as the one ‘individual’ who exhibits significant evidence of osteoporosis.
Two examples of DJD are observed in specimens from TU-1, Layer 8. First is a right patella with fairly extensive articular surface remodeling including porosity and destruction of the outer table and osteophytic growth along the inferior and lateral margins. The second example consists of a fifth lumbar vertebra (L5) with osteophytic lipping on the superior left lateral border (right side border broken away) and similar though slighter involvement of the inferior margin.
Also from TU-1 are the skeletal elements that comprise the lower legs and feet collected from Layer 9; these exhibit bony changes consistent with the early stages of DJD on most articular surfaces. The condyles of both femora, the tibial plateaux, the articular surface of the left patella (no right patella), and the distal fibulae all present areas of eroded outer table and porosity. There is minor lipping along the superior and medial margins of the articular surface of the patella and along the medial margin of the medial articular facet of the right tibia. In addition, the fovea capitis of the left femur exhibits moderate enlargement anteroposteriorly with slight ridging along the margin. Both tali (Figure 7) exhibit early osteophytic growth around the margins of the posterior calcaneal articular surface while the articular surfaces themselves have areas of eroded outer table characterized by porosity and possible eburnation. These features are mirrored on the calcanei. The right foot is slightly more involved as the anterior talar-calcaneal articulations show the very earliest stages of pathology while those of the left do not. Although in other studied prehistoric populations from North America (Bridges, 1991; Hemphill, 1999) and the Marianas (Douglas et al., 1997; Pietrusewsky et al., 1997) the ankles and feet exhibit relatively low rates for DJD, traversing the jagged limestone of Palau’s Rock Islands or other difficult terrain would create added stress on this joint complex and could explain the bilateral manifestation in a relatively young individual.
 View Details | Figure 7. Right (a) and left (b) talus and calcaneus from TU-1, individual 1 (the legs on the right of Figure 3). Note degenerative changes to the talar-calcaneal articulation. |
From TU-4 DJD is evident on two skeletal elements. A proximal left ulna exhibits slight degenerative changes impacting the inferior margin of the radial notch. A bony growth extends 2–3 mm inferiorly of the inferior margin and there is a small bony growth crossing the guiding ridge between the medial and lateral portions of the trochlear notch. A poorly preserved right patella may have early stages of DJD on the articular facets but preservation is such that it is difficult to determine the extent or even if it exists at all.
Among the five individuals from TU-4, one appears to exhibit extensive pathology throughout the skeleton that fits with a diagnosis of idiopathic osteoporosis (Aufderheide and Rorriguez-Martin, 1998). Although all individuals are quite fragmentary, one pelvis, one lumbar series, one patella, one distal ulna, and one set of hands and wrists show extensive degenerative changes.
The pelvis comprises fragments of a right innominate of a female that shows definite signs of osteoporosis as it is very lightly constructed with widely separated trabeculae. A right patella exhibits extensive destruction of the lateral articular facet with eburnation and a bony osteophyte extending inferiorly from the point. There is a circular growth of bone on the medial articular surface near midline approximately 5 mm in diameter and rising 0.5–1 mm above the surface.
The extensive involvement of the hands and wrists extends to nearly all elements of both right and left sides, with the right being the most severely affected. All carpal bones present (missing only the left lunate, capitate, and trapezoid and the right pisiform) exhibit extensive lipping around the articular surfaces, remodeling (particularly evident on triquetrals, trapazoid, and lunate), and deformation. Eburnation and outer table destruction occurs on the distal articular surface of the right hamate where the fourth and fifth metacarpals articulate. The carpal–metacarpal articulations of the other metacarpals exhibit moderate to severe involvement. In general, this extends to the joints of the fingers as lipping and bony remodeling are evident on the proximal articular facets of proximal, intermediate, and distal phalanges. The proximal first phalanges, particularly that of the right, show extensive remodeling, lipping, and eburnation. In addition a distal right ulna has DJD associated changes to the articular surface. The styloid process, though broken posteriorly, appears altered and there is evidence of eburnation on the articular surface.
The lumbar series (Figure 8a) has severe degenerative changes affecting all elements (L1–S1). In addition to extensive lipping throughout, the bone of all centra is light with horizontal structures diminished considerably. There is a compression fracture of the body of L5 with a corresponding spondylolytic separation through the pars interarticularis of the arch of L4 (Figure 8b).
 View Details | Figure 8. (a) Lumbar vetebrae 1–5 and 1st sacral vertebra (left to right). Note degenerative changes and porosity. (b) Spondylolysis of 4th lumbar vertebra. |
Whether the degenerative changes of the hands, wrists, and other joints results from afflictions such as rheumatoid arthritis or generalized DJD, separate from the osteoporosis, is difficult to determine. The bone loss of the spine, pelvis, and distal ulna and the compression fracture of a vertebral body are classic indicators of osteoporosis. However, as outlined by Aufderheide and Rodriguez-Martin (1998), the gross involvement of the hands and wrists is not generally characteristic of osteoporosis. The bilateral nature of hand/wrist involvement, the extreme degenerative changes, and that the hands are typically the first and most common joints to be affected suggest a diagnosis of rheumatoid arthritis. Alternately, activity induced DJD cannot be ruled out since the wrists are equally, or more severely, affected and wrist involvement is not characteristic of rheumatoid arthritis.
The more advanced DJD found in the lumbar series and the hands and wrists from TU-4 are among a suite of pathologies apparently affecting one individual. The best available comparative material, deriving from pre-Latte (1–1000 AD) and Latte (1000–1521 AD) period sites in the Marianas (Douglas et al., 1997; Pietrusewsky et al., 1997), indicates that the advanced stages of DJD and osteoporosis seen in this individual are relatively rare occurrences. Douglas et al. (1997) report rates of moderate to advanced DJD of the carpals and metacarpals in 5.2 and 2.9 percent of individuals, respectively, and osteoporosis of lumbar centra at 5.3 percent of vertebrae from Apurguan, Guam. Spondylolysis, in this case of the 4th lumbar vertebra, generally occurs in low frequencies in most populations (Merbs, 1989) but has been found at the relatively high rate of 21 percent of individuals from the Hyatt site on Guam by Ariazza (1997). Although generally attributed to physiological stress of the lower back, this case of L4 spondylolysis may very well be consequence of the compression fracture of L5 which, in turn, was most probably related to the severe osteoporosis seen in this individual.
Cranial pathology from elements recovered from TU-4 consists of apparent porotic hyperostosis on the parietal bones of Skull 2 (Figure 9). The parietal regions of this specimen exhibit possible porotic hyperostosis as manifested by pin-prick sized porosity over a wide area of the posterior region of both parietals and the superior portion of the occipital. In addition, inflation of portions of the parietals extends posterolaterally from near the mid-point of the sagittal suture. The ridges created by this inflation produce a porosity-free triangular depression superior to lambda.
 View Details | Figure 9. Porotic hyperostosis of parietals on Skull 2. |
Porotic hyperostosis has been linked with iron deficiency anemia, particularly in childhood (Stuart-Macadam, 1992). The specimen from Orrak is a good example of a lesion in the process of healing (Auferheide and Rodriguez-Martin, 1998) as it presents small pin-prick sized porosity of the outer table while retaining the characteristic inflated diplöe. In their evaluation of early Chamorro pathology, Douglas et al. (1997) note that porotic hyperostosis is uncommon, but then report that 12 crania exhibit the lesion. Although they are not clear as to the number of crania examined, this works out to approximately 25 percent (12/46) if the number of crania they examined in their analysis of cribra orbitalia is used here. Whether common or uncommon, the appearance of porotic hyperostosis in an individual at Orrak does indicate the presence of the anemia in this mortuary sample.
A right fibular shaft from TU-1 Layer 8 exhibits periosteal reactive bone growth along the interosseous crest. As the interosseous crest is the attachment point for the interosseous ligament that holds the tibia and fibula together it is often a stress point where chronic fatigue can result in the ligament attachment pulling away from the bone resulting in reactive bone growth along the crest.
Periostotis is commonly indicative of treponemal infection (Ortner and Putschar, 1985). In the Marianas it is present in prehistoric skeletal remains and is generally attributed to yaws (Pietrusewsky et al., 1997). However, in Palau, the skeletal material from Ngermereues Ridge, though fragmentary, has no periostitis reported (Reith and Liston, 2001). Although treponemal infection cannot be ruled out, no elements within the Orrak material show the classic signs of systemic infection (Ortner and Putschar, 1985). The slight manifestation of the periostitis in this case matches a scenario in which physical stress involving the interosseous ligament is the likely cause.
Developing a dental pathology profile from material recovered from Chelechol ra Orrak is difficult because of the low numbers of specimens. Although 71 teeth are available for study (67 permanent and 4 deciduous), 30 belong to one individual (Skull 4) and the rest are either isolated or associated with one to three other teeth in a fragmentary mandible or maxilla. Therefore, no reporting of frequencies by number of individuals affected is possible and doing so by tooth frequency is skewed by the high number of teeth originating from one individual.
The only pathologies noted are dental caries and enamel hypoplasia of both linear and pit forms. A total of one deciduous and five permanent teeth exhibit caries. In all cases the decay is localized interproximally or along the cemento-enamel junction (CEJ) and is generally confined to the anterior teeth (exceptions being a left M3 and left dm2 from TU-1, Layer 8). The individual represented by Skull 4 has small to moderate sized carious lesions on three teeth, left I1, left I2, and left lower C. The remaining tooth with caries is an isolated right I1 from TU-4, Layer 10. Although the low incidence and severity of caries may be explained by the small sample size and the fact that Skull 4 represents a relatively young person, it is also possible that the apparent ubiquitous chewing of betel (Areca catechu) among the people sampled here had some impact (Fitzpatrick et al., 2003). Hanson and Butler (1997, and references therein) cite numerous factors concomitant with betel chewing that have a cariostatic effect including: the fibrous nature of betel that aids in cleansing of teeth, the high pH oral environment created by the use of slaked lime that inhibits the formation of cariogenic acids, and increased saliva flow that enhances flushing of the mouth. Specifically, the individual represented by Skull 4 has light betel staining and only interproximal caries confined to the anterior teeth, a likely scenario in the case of betel chewing.
Two teeth exhibit both pit and linear enamel hypoplasia (LEH). A right upper C from TU-1, Layer 6 has one LEH line and several pits on the labial surface and a left upper C from TU-1, Layer 7 has two light LEH bands lingually and small pits on the mesial and distal margins of the crown. The other two examples consist of a right M2 from TU-3, Layer 10 with one LEH line near the CEJ and an isolated left upper C from TU-4, Layer 10 with numerous small pits across the labial surface.
The cemetery site at Chelechol ra Orrak is in the early stages of excavation and analysis and has the potential to become an important component in the search to understand the movements of early peoples into and within Micronesia. Spanning 3000 years, the site contains some of the earliest dates from the archipelago and represents the earliest large-scale burial site currently known in Micronesia or Polynesia. Radiocarbon dating indicates the site was used as a cemetery for about 1000 years near the beginnings of known human occupation of Palau. The suite of radiocarbon ages from the burials indicates that when fully excavated, this site, and perhaps others on Orrak Island that were identified in a 2002 survey, should produce an excellent mortuary sample and opportunities to learn more about the health of early Palauan populations.
Although the fragmentary nature of the skeletal material from Chelechol ra Orrak precludes a thorough analysis at this time, some insight into demographic structure is possible. Unlike many mortuary samples this one appears to have good representation of all ages and both sexes. The retrieval of elements belonging to several very young individuals is important because infants and the very young are frequently underrepresented because of the fragility of their skeletons and the often differential burial patterns involving children. What has been recovered so far indicates that this cemetery records a death assemblage that, as closely as possible (Wood et al., 1992), is a reasonable facsimile of the demographic profile of the living population.
Considering the incompleteness of the skeletal remains from Orrak, a reasonable pathology profile can be developed that gives some indication of the stresses encountered by the early inhabitants of Palau. Examples of dental pathology, osteoporosis, spondylolysis, degenerative joint disease (DJD), periostitis, and porotic hyperostosis that are found within the sample provide insight into the lifeways of Pacific Islanders within the last 2000–3000 years.
The pathologies visible in the individuals from Orrak are of the type generally expected for prehistoric populations. Osteoporosis and DJD, as expressed in the feet from TU-1 and the lumbar series and hands and wrists from TU-4, are classic examples of the degenerative changes the human skeleton can undergo during life (Jurmain, 1980; Merbs, 1983; Hemphill, 1999). The appearance of healing lesions of porotic hyperostosis in one individual indicates that anemia was present, as least to some degree. A single case of periostitis appears to be activity induced and not due to treponemal infection although, since yaws appears to be present in Guam (Pietrusewsky et al., 1997), more skeletal material is needed before conclusions can be drawn concerning etiology. Dental pathologies such as caries and enamel hypoplasia are expected in prehistoric populations such as that sampled at Chelechol ra Orrak although, for both, rates appear relatively low. For caries this can probably be explained by the cariostatic effects of betel chewing, whereas small sample size is likely a confounding factor for determining hypoplasia incidence.
As more of the cemetery is excavated, insight into burial patterns and the true extent of the cemetery’s size will be illuminated. Fostering a better understanding of interment distribution will further elucidate our understanding of Palauan mortuary behavior and contribute to our efforts at examining biological and cultural changes in prehistoric Pacific island populations. A larger sample size, including more complete individuals, will expand our ability to understand the skeletal biology, paleopathology, and paleodemography of this prehistoric population sample. In addition, a more comprehensive skeletal sample will hopefully allow us to conduct stable isotope and DNA analyses so that we may better comprehend the diet and genetic affinities of the early Palauans whose remains are interred at Chelechol ra Orrak.
This research was supported by grants and fellowships to Fitzpatrick from the National Science Foundation (SBR-0001531), the Sasakawa Peace Foundation (Micronesian Archaeological Survey with William S. Ayres and a Sasakawa International Trade and Development Fellowship), and the Center for Asian-Pacific Studies at the University of Oregon. Funding for AMS radiocarbon dates was enhanced by NSF sponsorship and cooperative agreements with NOSAMS (OCE-9807266) and the University of Arizona AMS Facility. We would like to thank the staff from the Palau Bureau of Arts and Culture for their help in coordinating logistical aspects of this project and continuing support over the years. The comments of John R. Lukacs on previous drafts were extremely helpful and the comments and suggestions of AS editor Dr Tsunehiko Hanihara and anonymous reviewers were also greatly appreciated. This project could not have been completed without help from students at the University of Oregon who worked on various phases of the project in the field and laboratory.
|