| Noboru Adachi, corresponding author. e-mail: nadachi@mail.tains.tohoku.ac.jp phone: +81-22-717-8026; fax: +81-22-717-8030 Published online 13 September 2005 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.041228 |
The relationship between individuals from a double burial is important for reconstructing prehistoric funerary behavior because of the possibility that such individuals were consanguineous or had a strong social relationship. Heretofore, kinship analyses of ancient skeletons have been performed based on morphological studies (e.g. Spence, 1974; Sjøvold, 1975; Doi et al., 1985, 1986; Alt et al., 1997). Above all, tooth size proportions were statistically confirmed to reflect the hereditary background of both parents through studies based on consanguineous modern Japanese (Doi et al., 1986), and similarity analysis using tooth size proportions has been applied to specific cases to infer kinship of prehistoric Jomon and protohistoric Kofun people (Hanihara et al., 1983; Doi et al., 1986; Tanaka, 1995; Matsumura and Nishimoto, 1996).
Although morphological methods are useful, they cannot provide decisive information about kinship. DNA analysis, an established method in forensic kinship determination, has been applied successfully to ancient skeletal specimens (e.g. Kurosaki et al., 1993; Oota et al., 1995; Delefosse and Hänni, 1997; Shinoda and Kanai, 1999). However, most ancient DNA studies are limited to mitochondrial DNA (mtDNA), because its survival in ancient skeletons is more likely than nuclear DNA due to its high copy numbers in a cell. Therefore, on most ancient skeletons, only maternal kinship can be estimated by DNA analysis.
To overcome this disadvantage, similarity analysis of tooth size proportions, which reflects the genetic effect of the paternal line as well as the maternal line, was combined with mtDNA analysis in recent studies, and this combination analysis was confirmed to be useful to infer the relationship between individuals unearthed from archaeological sites (Shinoda et al., 1998; Corruccini et al., 2002; Adachi et al., 2003).
In the present study, we examined the relationship between two adult female skeletons excavated from the double burial at the Usu-Moshiri site, Hokkaido, Japan.
The Usu-Moshiri site is located on a small island in Usu Bay in Date City, southwestern Hokkaido, Japan (Figure 1). Excavations at the site were carried out from 1985 to 1989 under the direction of one of the authors (Y.D.). Twenty-one pit graves of the Final Jomon to Epi-Jomon periods were found, and the skeletal remains of more than 50 individuals associated with many grave goods were unearthed (Ohshima et al., 1989, 1990).
 View Details | Figure 1. Map of the Usu-Moshiri site, located in the southwestern area of Hokkaido, Japan. |
Two double burials were found at the site: two juvenile individuals excavated in 1985, and two adults unearthed in 1988. The adult skeletons were well preserved, and were named 16A and 16B, respectively. These skeletons were discovered in a grave located between rocks covered with soil. The skeletons were buried in the fetal position, with the head of specimen 16A pointing west, and that of specimen 16B pointing east. Of the two double burials found from the Usu-Moshiri site, the juvenile skeletons were buried in a squatting position and their head directions were unclear. Therefore, the apparently opposing head directions were observed only in specimens 16A and 16B. Both skeletons wore shell bracelets, which might be Oh-tsutanoha (Patella optima) and Benkei-gai (Glycymeris albolineata), on their left forearms (Figure 2). Based on morphological evidence (Table 1), the age of these specimens was estimated at late thirties to early forties for 16A and late twenties to early thirties for 16B. Both specimens were sexed as female. As these two skeletons were situated side by side and maintained anatomical articulations, they were considered to have been buried at the same time. Judging from the latest style of Jomon pottery (Ohora A) found from the grave, the date of burial was estimated at about 2500 years ago, corresponding to the Final Jomon era in Hokkaido (Ohshima et al., 1989).
The preservation state of the skulls and permanent teeth in specimens 16A and 16B is shown in Figure 3 and Figure 4.
 View Details | Figure 2. The burial of skeletal specimens 16A and 16B. |

 View Details | Figure 3. Skulls of skeletal specimens 16A and 16B. |
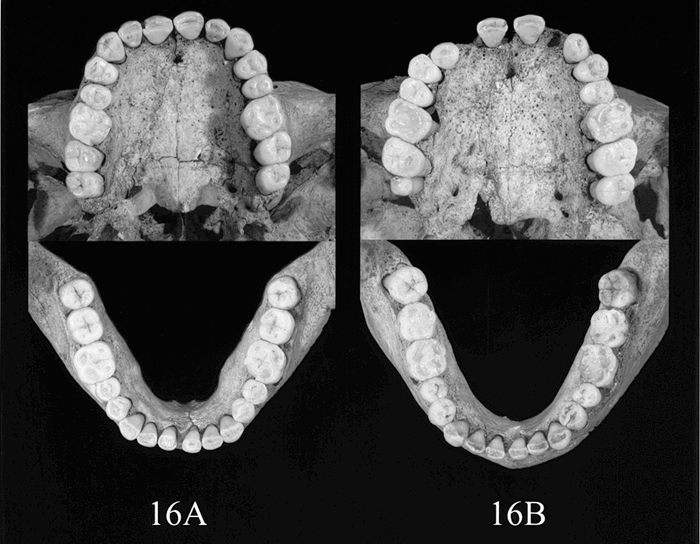 View Details | Figure 4. Dentitions of skeletal specimens 16A and 16B. In 16B, the upper right second incisor was intentionally extracted before death and the upper left second incisor was lost after the photograph shown in Figure 3 was taken. |
Measurement
Mesiodistal and buccolingual diameters of the permanent tooth crowns of each skeleton were measured to the nearest 0.01 mm with a digital sliding caliper (Mitutoyo NTD 10-10C) according to the method of Fujita (1949). The teeth examined and their sizes are listed in Table 2. In specimen 16B, antemortem extraction of the upper right second incisor was observed and the upper left second incisor was lost after reconstructing the skull. Therefore the upper second incisor was not used to estimate the similarity of tooth size proportions.
Each tooth was measured twice and the average was recorded. The average of both sides was used as the representative value of that tooth. However, the mesiodistal diameter of the lower right second incisor and buccolingual diameter of the upper right first molar of specimen 16A and buccolingual diameters of the lower left first incisor, lower left first premolar and lower left first molar of specimen 16B could not be measured due to damage. In these cases, the value of the tooth on the side measured was used as the representative value.
Estimation of the similarity of tooth size proportions
Based on the tooth crown measurements, Q-mode correlation coefficients (Sokal and Sneath, 1963) were calculated between specimens 16A and 16B. Before calculation, each crown diameter was standardized by using the means and standard deviations of female Jomon data reported by Matsumura (1995). The combinations of tooth crown diameters used for calculation of the Q-mode correlation coefficients are given in Table 3. These combinations were selected in such a way that the risk of accidental resemblance was less than 20% according to the criteria of Doi et al. (1986).
Intensive precautionary measures against modern DNA contamination were taken as described elsewhere (Adachi et al., 2003). DNA was extracted from the upper left third molar of specimen 16A and a piece of the left femur of specimen 16B according to the method of Adachi et al. (2004).
A segment of HVR 1 [np 16209 to 16402 (16209–16402), relative to the revised Cambridge reference sequence (Andrews et al., 1999)], which is known to be highly polymorphic and proved to be effective for ancient DNA analysis (e.g. Horai et al., 1989; Shinoda and Kanai, 1999), was amplified and sequenced as described elsewhere (Adachi et al., 2004).
In all combinations of the tooth crown diameters examined, the Q-mode correlations were consistently low (Table 3). Doi et al. (1986) regarded a Q-mode correlation coefficient of 0.5 as the criterion of kinship. However, in the present study, the value of the Q-mode correlation coefficient was negative in 13 out of 20 combinations of tooth crown diameters. The highest value of Q-mode correlation coefficient (0.451) was obtained by using six variables, and there was a tendency for an increase in the number of variables to lead to a decrease in the value of the Q-mode correlation coefficient (Table 3).
Even if the value of the Q-mode correlation coefficient is high, there is a considerable risk of stochastic resemblance when the number of variables is small. Generally, the risk of stochastic resemblance can be reduced by increasing the number of variables. This means that the increase in the number of variables raises the precision of similarity analysis. In consequence, the tendency observed in the present study indicates that the teeth of the two skeletons examined were morphologically dissimilar. Therefore, from a morphological viewpoint, the possibility that specimens 16A and 16B were relatives is low.
The mtDNA sequences of specimens 16A and 16B could be determined unambiguously. Compared to the revised Cambridge reference sequence, a C-to-T substitution at position 16223, a T-to-C substitution at position 16325 and a T-to-C substitution at position 16362 were observed in specimen 16A, and a C-to-T substitution at position 16223 and an A-to-G substitution at position 16247 were observed in specimen 16B. Basically, among maternal relatives, the mtDNA sequence is identical. On the other hand, as shown above, the mtDNA sequences of the two skeletons were apparently different. Therefore, maternal kinship of the specimens 16A and 16B is denied with a high level of certainty.
Lack of paternal kinship can be confirmed indisputably if STR and/or VNTR loci in chromosomal DNA can be analyzed, and these loci have been successfully analyzed in some studies of ancient DNA (e.g. Kurosaki et al. 1993; Hummel et al. 1999). Unfortunately, STR and VNTR analyses were not successful in the present study. Nevertheless, judging from both morphological and genetic evidence, specimens 16A and 16B are more likely to have been unrelated than consanguineous.
From the archaeological viewpoint, a difference in style of antemortem tooth extraction is estimated to reflect a difference in social and/or personal status, including genealogy (e.g. Harunari, 1982; Yamada, 2003). The fact that antemortem tooth extraction was observed only in specimen 16B is considered to strengthen this hypothesis. Moreover, the opposing head directions of specimens 16A and 16B might reflect the lack of kinship between them.
On the other hand, both skeletons wore shell bracelets on their left forearms, and both were buried in the fetal position. These common findings may indicate some relationship other than kinship between specimens 16A and 16B that resulted in these individuals being buried together.
The results of the present study suggest that the Jomon people, at least in Hokkaido, had the custom of burying genealogically different people side by side at the same time in the Final Jomon era. To clarify how genealogical differences are related to difference in burial style, or to examine if this type of double burial exists outside Hokkaido, skeletons discovered from other double burials should be analyzed morphologically, genetically and archaeologically.
The authors would like to thank Dr Yasuhiro Yamada of Shimane University, Japan for advice and help in writing this paper. A Grant-in-Aid for Young Scientists (No. 15770157) from the Japanese Ministry of Education, Science, Sports and Culture sponsored this study.
|