| Christopher J. Norton, corresponding author. e-mail: k_s_g@hotmail.com, cjn@ivpp.ac.cn phone: +86-10-8836-9254; fax: +86-10-6833-7001 Published online 25 May 2007 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.060605 |
Taphonomic data can contribute to a deeper understanding of modern human behavior only if signatures of human activity can first be distinguished from those of natural processes (Gifford, 1981; Gifford-Gonzalez, 1991; Lyman, 1994a; Reitz and Wing, 1999; Norton et al., 2007a). Once it has been determined that the faunal collection has at least a partial archeological component, then and only then can interpretations derived from the analysis of that bone assemblage be utilized to address questions relating to the evolution of human behavior. Nevertheless, many Pleistocene faunal assemblages that were excavated and studied before taphonomic analyses became standard (beginning in the 1980s) were collected under the assumption that hominins accumulated all of the faunal remains. A number of important Late Pleistocene bone assemblages from Northeast (NE) Asia were collected before taphonomic approaches developed in North America, Europe, and Africa became available in NE Asia. The Hanaizumi site, located in Japan, exemplifies this point because it was excavated without the application of taphonomic methodologies and was originally interpreted to have been the direct result of human hunting activity without supporting taphonomic studies (e.g. Matsumoto et al., 1959a, b).
Hanaizumi is an open-air site located on the middle terrace of the Kinryu River, approximately 10 km south of Ichinoseki City, Iwate Prefecture, Japan (Figure 1). The landowner, Seiji Numakura, originally discovered the site in 1927 during digging to create a pool to drain excess water from his farmland. The discovery of fossilized animal bones caused the digging to be stopped. The potential of Hanaizumi as an archeological site was later brought to the attention of Morisuke Sasaki and Hiroshi Sone who directed the first excavation of the site in the late autumn of 1953. By 1988 fieldwork had been conducted at Hanaizumi 15 times, with floral and faunal remains found throughout the deposits (Figure 2). Analysis of these biological residues has facilitated paleoenvironmental reconstructions of Honshu, Japan, between 35000 and 10000 BP (Matsumoto et al., 1959b; Yasuda, 1978; Takeuti and Ozaki, 1987; Hanaizumi Site Excavation and Research Group, 1993).
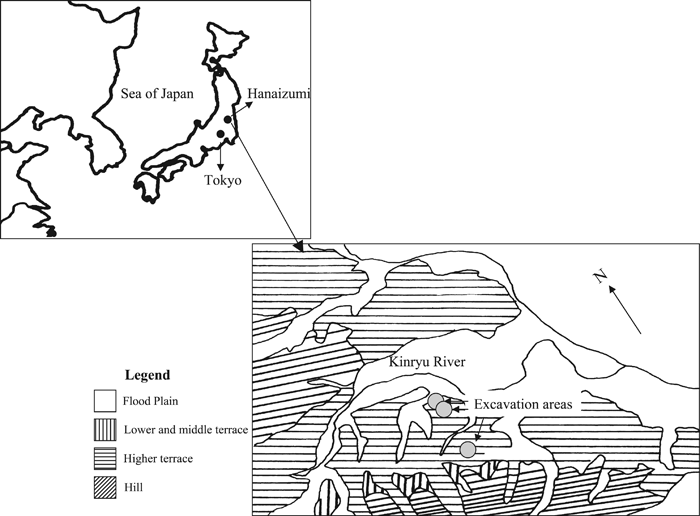 View Details | Figure 1. Location of Hanaizumi in Japan and along the Kinryu River. |
 View Details | Figure 2. General location of the Hanaizumi site along the Kinryu River. The general site area is currently approximately 200 m from the river. |
The Hanaizumi Formation lies above the Pliocene Ariga Formation and below Holocene deposits. Five primary geological units have been discerned at Hanaizumi: (1) Hill; (2) Higher Terrace; (3) Middle Terrace; (4) Lower Terrace; and (5) Flood Plain (see Figure 1). The Hill is comprised of sand, silt, tuff, and gravel sediments and has been dated to the Pliocene. The Higher Terrace ranges between 50 and 80 m above the riverbank and is composed primarily of gravel with a layer of volcanic ash directly above it. This level is about 1.5 m thick. The Middle Terrace, situated 30–50 m above the Kinryu River, is comprised of sand, silt, gravel, and peat and is approximately 4 m thick. The Lower Terrace consists of loose gravel and is situated 25–30 m above the riverbank. The Flood Plain expands along the course of the present-day Kinryu River and lies 20–25 m above the current water level. Floral and faunal remains were found on the Middle Terrace, which dates to the Late Pleistocene. The fossils were located in four peat layers (primarily from layers 3 and 4) that alternated with strata comprised of sand and clay (Figure 3; Matsumoto et al., 1959b; Takeuti and Ozaki, 1987; Yasuda, 1978).
 View Details | Figure 3. Stratigraphic profile of the Hanaizumi site. Note the four dark layers that are clearly interwoven with the sandy clay strata. Most of the faunal remains originated in the third and fourth peat layers. |
Fourteen 14C samples collected up to 1975 document the chronology of the depositional sequence (Table 1). Dates from the 1958 excavation consistently indicate a depositional sequence from ca. 21000 BP (from the second peat bed) to ca. 35000 BP (from the basal gravel). The age of the Hanaizumi deposit is also indicated by the presence of AT Tephra (24000–22000 BP), identified in the second peat layer (Yasuda, 1978; Takeuti and Ozaki, 1987). An additional 11 14C dates were analyzed in the 1980s from different areas of the Hanaizumi site (Hanaizumi Site Excavation and Research Group, 1993). In general, the ages overlap with the previous studies, though the majority of the dates are post-Last Glacial Maximum.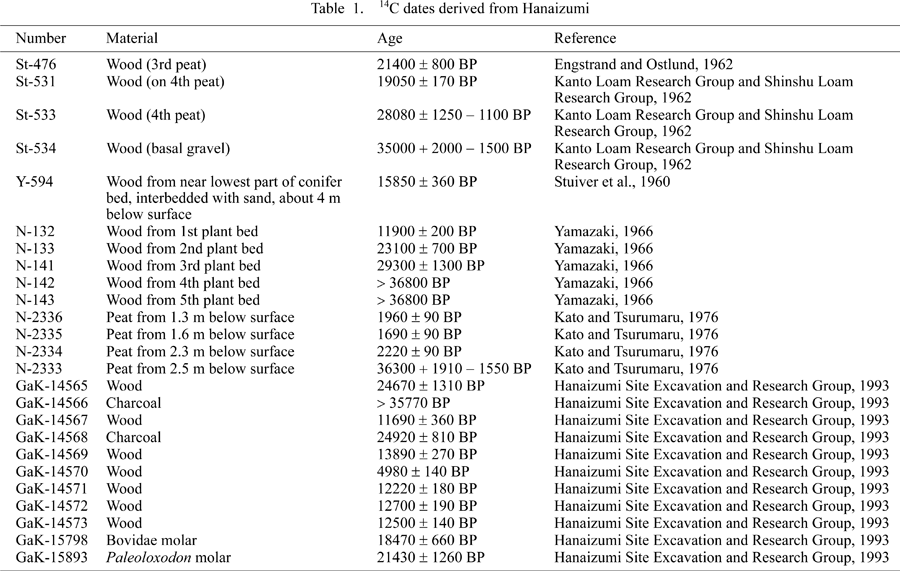
The dates for peat layers 2–4 from the 1975 excavation ranged between ca. 2220 and 1690 BP (Table 1: N-2334–N-2336), and differ significantly from data from previous studies that were taken on what are assumed to be the same peat layers. One plausible explanation for the difference in dates from the two excavations is that the series of ages derived from the 1958 pit were from deposits in the northeastern area of the site and those from the 1975 pit from deposits in the southeastern section. Perhaps peat layers from the 1975 test pits were subjected to a significant degree of postdepositional disturbance and mixing of materials between the different strata that resulted in a younger age range for the materials studied than that of the residues from the 1958 test pit. Another possibility is that the dates are correct, but are associated with a more recent cultural deposit. A second Hanaizumi faunal assemblage is stored in the National Science Museum, but is from an Edo Period (1603–1867 AD) archeological site. Kato and Tsurumaru (1976), whose research team dated the material from the 1975 test pit, are the only ones whose results appear to differ significantly from those of the other five research groups that have published chronometric dates (e.g. Stuiver et al., 1960; Engstrand and Ostlund, 1962; Kanto Loam Research Group and Shinshu Loam Research Group, 1962; Yamazaki, 1966; Hanaizumi Site Excavation and Research Group, 1993). The discrepancy between the depositional chronologies of the peat layers notwithstanding, it is generally accepted that the age range for the Hanaizumi Formation’s lowermost three peat layers is 35000–12000 BP.
Very few indisputable artifacts were discovered in the Hanaizumi deposits. Even though a number of fractured stones appear in the deposit in the same context as the faunal remains, the lack of clear striking platforms, bulbs of percussion, and concentric lines suggests that the stones are likely ecofacts (C.J.N., personal observation). One flake tool and one hammerstone were recovered during the 1975 excavation, but they may have been deposited secondarily. Due to the paucity of lithics in the site, the primary researchers (Matsumoto, Mori, Marui) proposed that the bone, antler, teeth, and wood recovered during excavations were probably used as tools. Since many of the wood specimens discovered in the peat deposits looked fresh and unworn, and appeared to have been purposely cut in distinct ways, it was proposed that this reflected human modification (Matsumoto et al., 1958, 1959a, b). It was argued that many Bison priscus ribs were purposely fractured and the edges worked in order to use them as tools. It was postulated that this bone tool technology was similar to that of Late Paleolithic Siberian hunter-gatherer groups (Kato, 1975).
Several pieces of charcoal and calcified bone were also found in the Hanaizumi deposit. However, these are few in number and fragmented, which suggested to the excavators that these residues could be the product of either dispersion or evidence of a naturally occurring fire, rather than having been the result of a concentrated hunter-gatherer campfire. No traces of post molds were discovered that could have suggested some type of structure and the notion that the site served as a campsite (Matsumoto et al., 1959b). Accordingly, besides the proposed bone and wooden tool culture, published evidence of hunter-gatherer activities at the site is minimal. Nevertheless, our recognition of cut marks on at least a few of the B. priscus bones is clear evidence of at least some degree of human involvement without the presence of unambiguous artifacts (see Bunn, 1994).
Pollen recovered from the Hanaizumi Formation facilitated reconstruction of the paleoenvironment (Figure 4; Table 2). The environment around the site can be broken down into three primary climatic phases (HA-I, HA-II, HA-III). HA-I is comprised of the Basal Gravel bed and the sand and silty sand strata directly above it. Sub-Alpine florae (e.g. Picea, Pinus, Tsuga, Abies, Betula) are most common in the HA-I palynological assemblage. However, temperate genera are represented in moderate numbers (e.g. Cryptomeria, Fagus, Quercus), suggesting a warming trend ca. 40000–30000 BP. Picea, Pinus, Tsuga, and Abies continue to be most common in HA-II (ca. 30000–10000 BP). Betula only appears intermittently and when it does appear, the percentage of Picea, Pinus, Tsuga, and Abies decreases. This has been interpreted to indicate slightly warmer sub-zones during HA-II. However, the warmer temperate element that appears in moderate percentages during HA-I (e.g. Cryptomeria, Fagus, Quercus) is less frequent during HA-II. This correlates well with the onset of the Last Glacial Maximum and it has been postulated that the environment in the region then was 5–7°C lower than it is today. HA-III is dominated by warmer floral elements (Quercus, Cryptomeria, Fagus, and Carpinus) and represents the Holocene based on associated dates (Yasuda, 1978; Takeuti and Ozaki, 1987).
 View Details | Table2 |
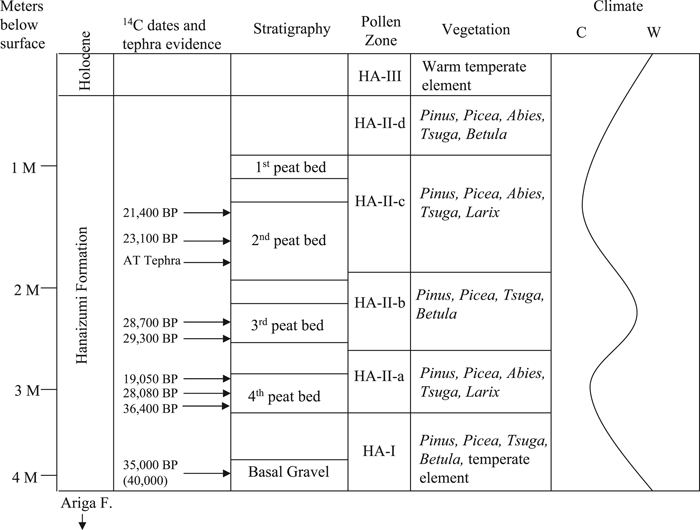 View Details | Figure 4. Stratigraphic profile of Hanaizumi site with associated 14C dates, vegetation, and climate reconstruction (after Yasuda, 1978; Takeuti and Ozaki, 1987). |
The Hanaizumi faunal remains, found primarily in peat layers 3 and 4, are comprised of Cervus, Megaceros, B. priscus, Palaeoloxodon, Alces, and Lepus (Table 3, Table 4). B. priscus is the most common animal present, comprising 88% of the faunal assemblage. All of the faunae represented suggest a cold climate, which correlates well with pollen analysis. Palaeoloxodon is present in the form of a complete tusk and a well-worn M2. One very interesting specimen recovered during the excavations was a shark tooth (Carcharodon megarodon), which is clearly not indigenous to the area (Matsumoto et al., 1959b; Naora, 1959).
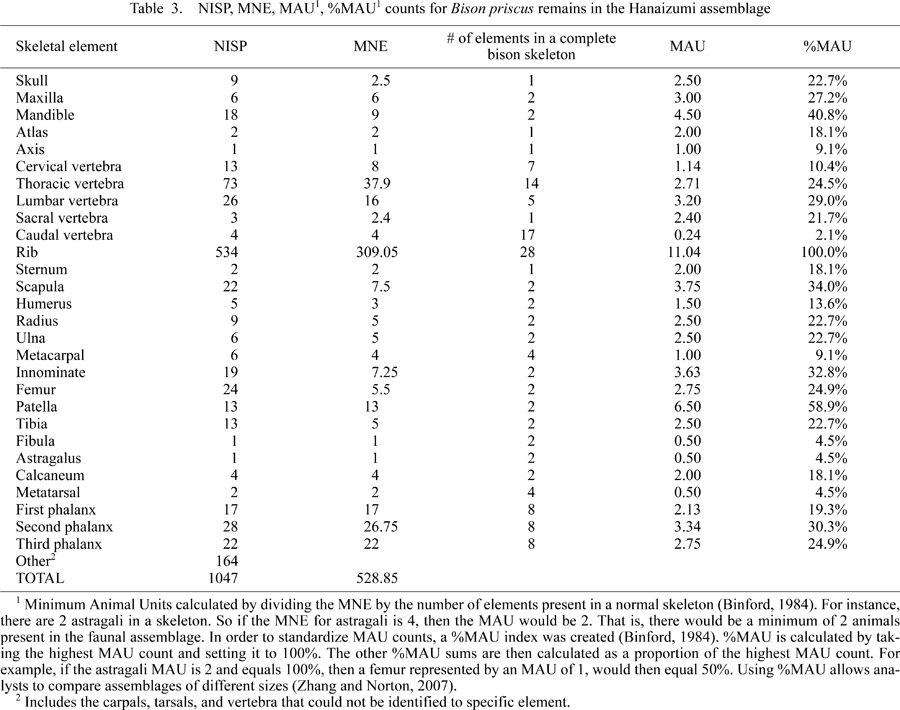 View Details | Table3 |

The Hanaizumi faunal assemblage consists of 1182 specimens. The majority of the collection is stored in the National Science Museum [number of specimens (NSP) = 1027] in Tokyo and was studied between April and May 2005, while a smaller subset is housed in the Saito Ho-on Kai Museum (NSP = 155) in Sendai and was analyzed in May 1998. The two faunal assemblages were excavated during different field seasons, but are considered to have derived from the same stratigraphic contexts. The original excavation reports and maps have been lost, which means associating each individual specimen to a specific stratigraphic level is impossible. Accordingly, they are grouped together for the present analysis.
The number of identifiable specimens (NISP) is comprised of 1056 bones (Table 3, Table 4). B. priscus rib specimens dominate the faunal collection. The Minimum Number of Element (MNE), Minimum Animal Units (MAU), and %MAU counts were derived following procedures discussed by Binford (1981, 1984), Grayson (1984), and Lyman (1994a, b).
Taxonomic and taphonomic identifications on the assemblage housed in the Saito Ho-on Kai Museum were made by C.J.N. Taxonomic identifications of the collection stored in the National Science Museum were done primarily by N.K., a curator of vertebrate paleontology, with the taphonomic data being collected by C.J.N. Data were entered into a Microsoft Access relational database that C.J.N. created specifically for the study and which follows data entry and analytical procedures utilized in other studies (e.g. Norton et al., 1999, 2007b; Norton, 2000). SPSS 12.0 was employed for statistical analyses. For the correlation analyses, rank orders were used because the Hanaizumi %MAU distribution is non-parametrically shaped (Lyman, 1994a, b; Carlson and Pickering, 2003). Statistical results were considered significant if p ≤ 0.05.
Standard zooarcheological data were recorded, including taxon, skeletal element, bone shape, bone completeness, side, sex, and ontogenetic age when applicable. Length and width measurements were recorded in addition to bone fracture angle and texture data only on the long bone fragments. The minimum number counts were based on summing the percentage of each anatomical landmark present for each individual element. The presence/absence of cut mark, percussion mark, tooth mark, tooth scoring, burning, abrasion/trampling, rodent gnawing, and root etching data were recorded, in addition to the weathering stage of the bone surface and the bone surface condition (Behrensmeyer, 1978; Brain, 1981; Shipman, 1981; Haynes, 1983, 1991; Johnson, 1985; Davis, 1987; Blumenschine and Selvaggio, 1988; Potts, 1988; Lyman and Fox, 1989; Andrews, 1990; Capaldo and Blumenschine, 1994; Lyman, 1994a; Fisher, 1995; Capaldo, 1997; Reitz and Wing, 1999). Identifications of the bone surface marks were made using a 16 × hand lens. Each specimen was examined under a strong incandescent light and carefully rotated at different angles. We found this to be the best method for studying surface marks, in particular for identifying the presence/absence of microstriations of cut marks and internal crushing of marks left by hominin percussion, and carnivore tooth pits and scores (Blumenschine, 1995; Blumenschine et al., 1996).
Throughout this analysis, the Hanaizumi collection will be compared to ethnoarcheological and actualistic/experimental taphonomic studies. Due to the lack of similar taphonomic research in NE Asia, the African record is currently the most applicable for comparative research. Some cases (e.g. actualistic carnivore, especially hyena, bone feeding experiments, Hadza use of metal knives instead of stone tools) may not be directly comparable to the NE Asian record. Despite this limitation, we chose the most appropriate models currently available, which we consider to potentially have a broad range of applications. However, such details need to be demonstrated by the development of actualistic models more directly applicable to a NE Asian Late Pleistocene setting.
One problem that often biases skeletal element profiles of faunal assemblages collected before the 1980s was the discard or loss of many difficult-to-identify specimens (Marean and Kim, 1998; Norton et al., 2007b). However, even though the excavations were conducted over four decades ago it appears that many difficult-to-identify specimens were retained (C.J.N., personal observation). Additionally, given the importance of identifying and including long bone midshaft fragments in any calculation of minimum numbers (Marean and Spencer, 1991; Bartram and Marean, 1999; Pickering et al., 2003), every attempt was made to identify as many long bone midshaft fragments as possible. After a vertebrate paleontologist (K.N.) made the initial taxon and element identifications, C.J.N. examined the entire assemblage in order to assign specimens to skeletal elements, with an emphasis on the limb bones. Once the data collection stage was completed, 126 specimens were still ‘non-identifiable’; most of these were probably either rib shafts or vertebral spinous or transverse processes. Of the non-identifiable specimens, only 17 could be considered to have come from long bones, of which only 10 were midshafts (the other 7 specimens were non-identifiable epiphyses and near-epiphyses fragments). Due to the absence of any identifying features on these specimens, it was impossible to ascertain confidently from which long bone each originally derived. However, even if these specimens were identifiable, and we assume a random distribution, each limb bone count would increase by less than two specimens (humerus, radius, femur, tibia, metacarpal, and metatarsal). This would not significantly change the overall skeletal element representation, which is dominated by rib fragments. Exclusion of the difficult-to-identify long bone midshaft fragments does not bias the bison skeletal element profile.
In addition to utilizing anatomical landmarks associated with midshaft regions of appendicular bones (e.g. femoral linea aspera, tibia internal and external nutrient foramen), whenever possible refitting was attempted (Marean and Kim, 1998). Only four refits were found (tibia, femur, ulna, mandible), with two of them from long bones that included midshaft sections. The refits did not significantly alter the MNE counts of the bison assemblage.
To ascertain the level of density mediated destruction, %MAU values were compared with bone mineral density scores derived for bison (Kreutzer, 1992) utilizing the scan sites that Lyman (1994a) refers to as ‘traditional’ (Table 5). There are two reasons why Kreutzer’s bone density data were used here rather than similar data derived from computed tomography (CT) (see Lam et al., 2003; Lam and Pearson, 2004). First, currently no CT data that we are aware of exist for bison, and second, Lam et al. (2003) admit that shape-adjusted photon densitometry [which Kreutzer (1992) used] is better than standard photon densitometry (see Lyman, 1984) and can closely match results from CT scanning.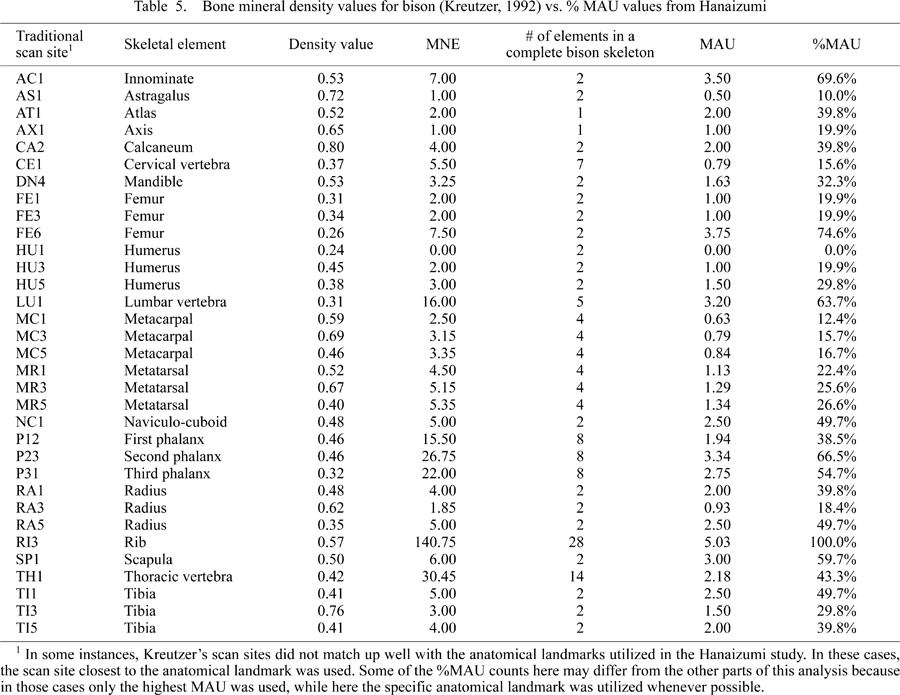
Strongly correlated with density-mediated destruction is the effect of carnivore ravaging on a faunal assemblage (Brain, 1981; Marean et al., 1992; Blumenschine and Marean, 1993; Munson and Marean, 2003; Pickering et al., 2003). The supposition is that if a strong positive correlation exists between %Survivorship derived from experimental studies involving carnivore ravaging of carcasses and the Hanaizumi %MAU, then it suggests that carnivores heavily influenced the formation of the skeletal part profile. We have evaluated this potential influence on the faunal assemblage (Table 6).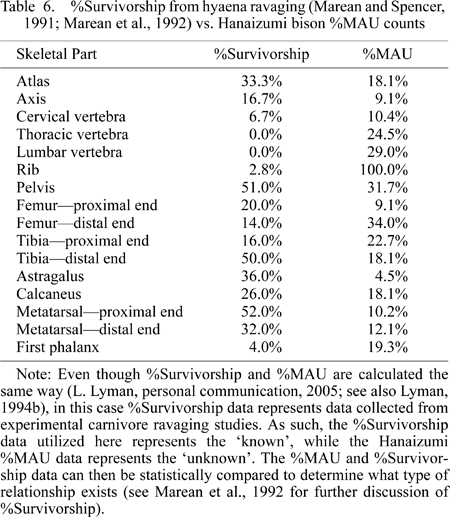
The Hanaizumi %MAU values were compared with a food utility index to determine whether human transport decisions may have influenced the frequencies of each skeletal element. Food transport indices have been created for a variety of taxa and are based on a combination of meat, fat, grease, and marrow that each element normally contains (Binford, 1978; Jones and Metcalfe, 1988; Metcalfe and Jones, 1988). The Bison Modified Total Products (BMTP) index (Emerson, 1990; Lyman, 1994a) was utilized because it is the closest food utility index that might be applicable to the faunal assemblage, which is dominated by B. priscus remains (Table 7).
Experimental studies have indicated that fluvial processes will preferentially move different skeletal elements based on weight, size, and shape. In general, the smaller, lighter specimens will move first, and the larger, heavier elements last (Voorhies, 1969; Behrensmeyer, 1975; Hanson, 1980). In order to exam the relationship, the bison %MAU data were plotted against the Saturated Weight Index (SWI) derived for zebra. In addition, correlation statistics were calculated in order to determine whether fluvial transport may have influenced the formation of the skeletal part profile (Behrensmeyer, 1975; Frison and Todd, 1986; Lyman, 1994a). The SWI for zebra was employed here because zebra bones are similar in overall size to the Hanaizumi bison (Table 8). In addition, osseous material moving through water will display signs of abrasion by silty sand (Hanson, 1980; Shipman and Rose, 1983). Abrasion data were also collected for the faunal assemblage.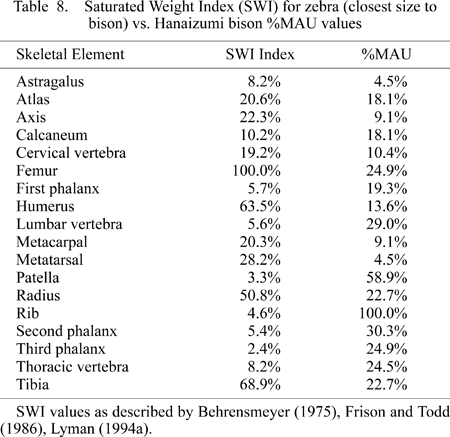
In general, it is assumed that a higher percentage of cut marks and percussion marks on long bones suggest primary access by hominins (see O’Connell et al., 2002; Blumenschine and Pobiner, 2006 for review; see Lyman, 2005 for critique of cut mark analyses and interpretations). This in turn has been interpreted to indicate successful hunting and/or confrontational scavenging large game procurement strategies. However, a high percentage of tooth marks, particularly on the long bone midshafts, suggest primary access by carnivores and secondary access by hominins to the same carcass. Cut mark, percussion mark, and tooth mark data derived through experimental (Blumenschine, 1995) and ethnoarcheological research (Lupo and O’Connell, 2002) were compared to the Hanaizumi faunal collection in order to determine timing of carcass access by hunter-gatherers and carnivores. We are well aware of the vast dataset accumulated from actualistic/experimental taphonomic and ethnoarcheological research conducted in Africa to distinguish primary from secondary access to a carcass (e.g. Binford et al., 1988; Bunn et al., 1988; O’Connell et al., 1988; Bartram, 1993; Selvaggio, 1994, 1998; Capaldo, 1995, 1997, 1998; Bartram and Marean, 1999). We chose Blumenschine’s actualistic study because it is the foundation from which many of these carnivore feeding experiments developed, and Lupo and O’Connell’s ethnoarcheological data, because they published cut mark and tooth mark counts broken down by skeletal region.
In a series of actualistic and experimental studies carried out in the Serengeti plains, Tanzania, Blumenschine (1986, 1988, 1995) showed that the varying percentages of tooth mark, percussion mark, and cut marks can suggest timing of access to a carcass. Specifically, Blumenschine carried out a series of experiments that modeled a situation in which carnivores had primary access to the meaty long bones. Appendicular elements presented to ravaging carnivores were unmodified by the researcher and were referred to as ‘carnivore-only experiments.’ In other actualistic studies, Blumenschine simulated instances in which hominins had primary access to the long bones, and he broke the bones open for the marrow. Ravaging carnivores then scavenged what was left of the long bones. These experiments were referred to as ‘hammerstone to carnivore.’ These experimentally derived data are compared to the long bone assemblage, which contains evidence of tooth marks and cut marks (Figure 5, Figure 6).
 View Details | Figure 5. Examples of tooth-marked bison bone from Hanaizumi. |
 View Details | Figure 6. Examples of cut-marked bison bone from Hanaizumi. |
The cut mark and tooth mark data are compared with the published cut mark and tooth mark ratios from Hadza home bases and butchering stands (O’Connell et al., 1988, 2002; Lupo and O’Connell, 2002). Butchering stands represent locations where the Hadza successfully hunted (i.e. primary access) and butchered the carcasses that were later transported to a base camp. Even though data on the Hadza base camp large game processing strategies were also available, the lack of clear artifacts or other cultural traces (e.g. hearths, fire-cracked rock) suggests that if Hanaizumi is to be considered an archeological site, then it was probably a kill site and not a base camp locality. Long bone frequencies are utilized here because they are thought to be the most important group of elements for addressing questions dealing with hominin-carnivore interactions (Marean et al., 1992; Blumenschine and Marean, 1993; Blumenschine, 1995). In addition, rib cut mark and tooth mark data are also compared, as ribs are by far the most common element in the assemblage and also displayed evidence of bone surface modification.
This analysis concentrates on reconstructing the taphonomic history of the most abundant animal present in the Hanaizumi assemblage, B. priscus. Even though the intent was to reconstruct mortality profiles, the sample of mandibles (NISP = 9 left and 9 right) and loose teeth (NISP = 44, highest count was incisors = 21) is too small to draw statistically meaningful conclusions. Accordingly, taphonomic analysis of the assemblage focuses on skeletal element representation and bone surface modification.
Density-mediated destruction
The relationship between the %MAU and the ranked ‘traditional’ bone mineral density scores indicates no correlation (rs = −0.165, n = 33, p = 0.360; Figure 7). Even though it appears that relatively little density-mediated destruction affected the faunal assemblage, we compare the bison %MAU counts with hyena-ravaged collections to ascertain if carnivores may have significantly influenced the composition of the bone collection. Spearman’s correlation analysis of the ranks supports the visually apparent relationship (Figure 8); a negative relationship exists (rs = −0.458, n = 16, p = 0.074). However, the correlation is not significant.
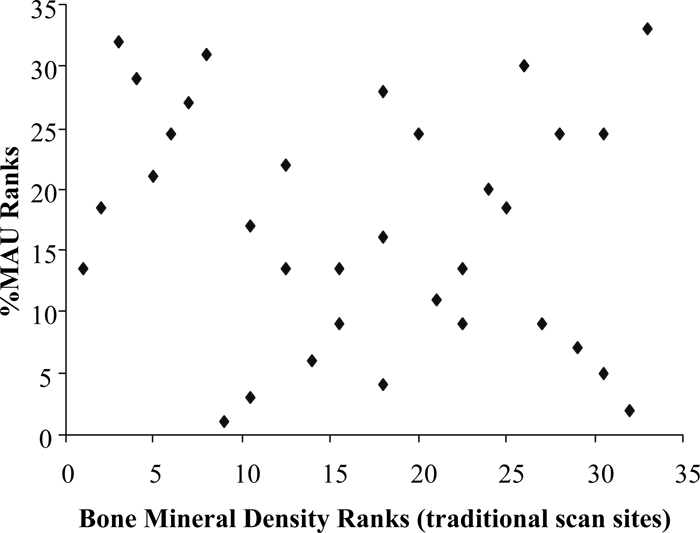 View Details | Figure 7. Scatterplot of rank orders for bison bone mineral density (after Kreutzer, 1992; Lyman, 1994a) and Hanaizumi %MAU. |
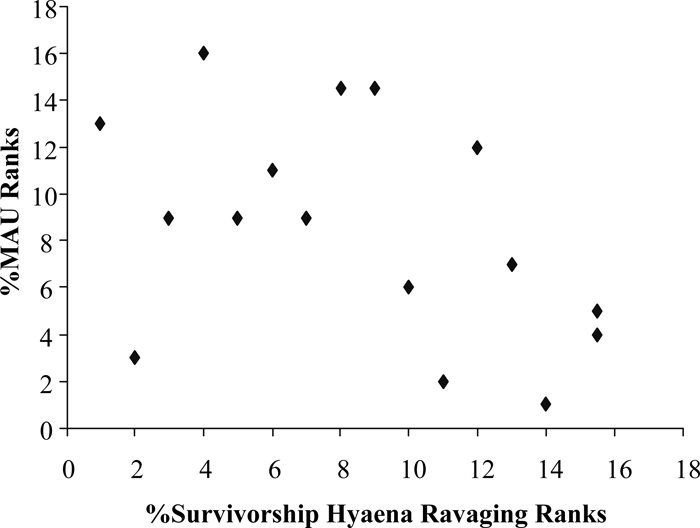 View Details | Figure 8. Scatterplot of rank orders for %Survivorship Hyena Ravaging (after Marean et al., 1992) and Hanaizumi bison %MAU. |
Bison modified total products (BMTP) index
A scatterplot of the Hanaizumi %MAU and BMTP suggests a positive, but weak, relationship (Figure 9). Correlation statistics were calculated to test this hypothesis further. Correlation analysis supports the observation that no relationship exists (rs = 0.280, n = 30, p = 0.134).
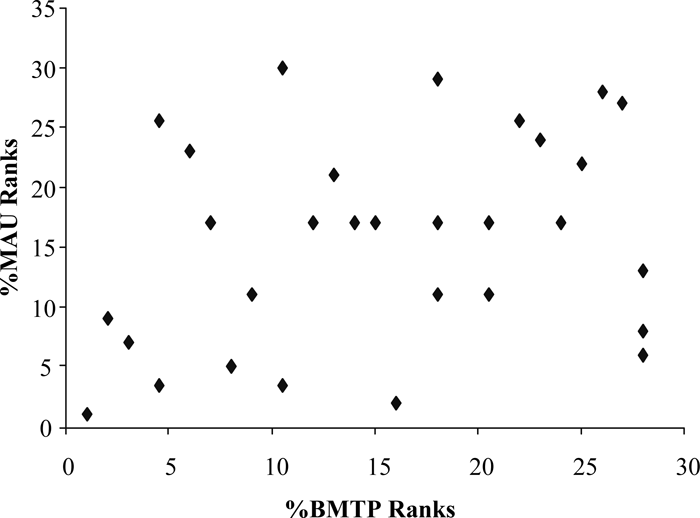 View Details | Figure 9. Scatterplot of rank orders for bison %BMTP (Bison Modified Total Products; after Emerson, 1990; Lyman, 1994a) and Hanaizumi %MAU. |
Evidence for fluvial transport?
Correlation analysis between the SWI for zebra and the bison %MAU indicates a statistically significant negative relationship (rs = −0.499, n = 18, p = 0.035; Figure 10). This correlates well with the fact that 38% of the faunal collection displays evidence of abrasion (Figure 11).
 View Details | Figure 10. Scatterplot of rank orders for Saturated Weight Index (SWI) for zebra (after Behrensmeyer, 1975; Lyman, 1994a) and Hanaizumi bison %MAU. Values for zebra were utilized here because in terms of general size they are closest to the Hanaizumi bison. A negative relationship exists indicating the lighter specimens (e.g. ribs and vertebrae) are more represented than the heavier skeletal elements (e.g. femora, tibiae). |
 View Details | Figure 11. Examples of abrasion on bison rib bone from Hanaizumi. Note the randomly oriented, shallow striations. |
Hanaizumi and experimental long bone tooth mark and percussion mark data
The percentage of tooth marked bones is not significantly different from Blumenschine’s hammerstone to carnivore percentages (χ2 = 0.171, df = 1, p > 0.05; Table 9). However, significantly fewer tooth marks appear on the Hanaizumi bison long bones vis-à-vis Blumenschine’s carnivore only experimental data (χ2 = 112.2, df = 1, p < 0.001). Significantly fewer long bones display evidence of percussion marks when compared with Blumenschine’s hammerstone to carnivore percussion mark data (χ2 = 8.625, df = 1, p < 0.05).
Hanaizumi and ethnoarcheological cut mark and tooth mark data
The cut mark data on the long bones indicate no significant difference between Hanaizumi and the Hadza butchering stand (χ2 = 1.53, df = 1, p > 0.05; Table 10). However, statistical comparison found that the long bones are tooth marked at a significantly higher rate than the Hadza butchering stand carcasses (χ2 = 22.21, df = 1, p < 0.05). Comparison of the rib data also resulted in a mixed outcome. The cut mark percentage for the ribs is significantly lower than that of the Hadza butchering stand (χ2 = 5.22, df = 1, p < 0.05), but the tooth mark on ribs percentages are not significantly different (χ2 = 1.21, df = 1, p > 0.05).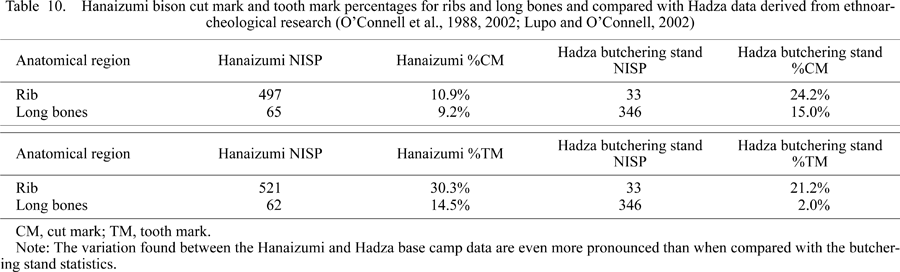
Evidence for burning?
Very few specimens in the Hanaizumi faunal assemblage display evidence of having been burned. Among the two most common elements, ribs and vertebrae (NISP = 705), only one specimen appears to have been calcined. Only one of the 66 limb bone specimens displays evidence of burning. Pieces of charcoal were discovered during the excavations, but whether they were found in a primary context or were a secondary deposit is not clear (Matsumoto et al., 1959b).
Comparison of the Hanaizumi bison skeletal part profile with bone mineral density and with hyena ravaging data indicates a negative, statistically non-significant relationship. Bone mineral density and hyena ravaging are not significant predictors of the bison skeletal part profile. The lack of a significant correlation of the Hanaizumi %MAU ranks with the BMTP index suggests that human transport decisions were not a significant influence on the formation of the faunal assemblage.
Statistical comparisons of cut mark, percussion mark, and tooth mark percentages between the Hanaizumi dataset and ethnoarcheological and experimental data reveal no clear pattern in terms of timing of access to the bison carcasses. If Hanaizumi was a kill site, then the cut mark and tooth mark data from the long bones and ribs should have been more in line with the Hadza butchering stand data, which is the equivalent of a kill site, or to Blumenschine’s percussion-to-carnivore data reflecting primary access to the carcass by hominins. If hunter-gatherers had secondary access to the large game found at the site, then the percentages for the tooth marks should be closer to Blumenschine’s experimental carnivore-only studies. In general, the cut mark and tooth mark data appear to correlate better with the Hadza butchering stand percentages. However, some of the results (e.g. percussion mark data) conflict with the model in which hunter-gatherers successfully hunted and killed the bison at the locality, processed the carcasses there, and transported selective body parts to or from the site.
Fluvial activity may have transported some of the specimens from or to the site, and we believe this is the primary explanation for the accumulation of B. priscus remains at Hanaizumi. The only strong, significant correlation that exists is when the dataset is compared to the SWI. The correlation was significantly negative, which indicates that lighter body parts were more abundant in the assemblage than expected. This result correlates well with the fact that 38% of the collection display damage from abrasion by silty sand and/or trampling.
We suppose that the silty sand that is found deeply embedded in many of the specimens (Figure 12) indicates that the Kinryu River, which currently flows roughly 200 m north of the excavation site, once flowed much closer or even directly over the area. Some of the specimens are worn on only part of the element. Sand and silt particles appear deep in the crevices of the worn regions of bones (Figure 13). This suggests that at least some of the faunal specimens were partially buried before water started to flow over them and may indicate that more than one bone depositional event occurred at the site.
 View Details | Figure 12. Example of sand and silt embedded in bison bone from Hanaizumi. |
 View Details | Figure 13. Example of sand and silt embedded in bison bone from Hanaizumi. |
It is difficult to make a strong case for Hanaizumi to have served as either a bison kill site or a hunter-gatherer home base. Even though we suppose that hunter-gatherers accounted for at least some of the faunal assemblage, we propose that procurement of the bison probably occurred up-river where the skeletons were partially processed. Later, fluvial processes transported carcasses to the site. Multiple depositional events are suggested because some of the elements display only localized abrasion while other specimens have abrasion damage over the entire specimen. Based on the 14C studies on pre-1980 materials, the time range of the deposition is roughly 14000 years (ca. 35000–21000 BP) and the bison MNI is only 11. This means that bison carcasses or parts thereof accumulated at the site at the rate of less than one per thousand years. Even though it is likely that the carcasses accumulated at a faster rate than that, we consider it is unlikely that the assemblage represents a one-time catastrophic mortality event. Nevertheless, the faunal materials were concentrated in two separate peat layers (3 and 4), so it may be possible that there were two separate catastrophic mortality events. We are currently unable to examine this further because of the lack of three-dimensional provenance records from the excavations.
Additional evidence that Hanaizumi is a non-human accumulation is the lack of clear artifactual materials. Due to the absence of stone tools, it was proposed that bone and wood implements were utilized at the site. However, these are dubious ‘tools.’ Water action, over a period of time, can alter the shape of a bone or a piece of wood, sometimes making it appear to have been intentionally worked (Hanson, 1980; Shipman and Rose, 1983). No evidence of a settlement or hearths was discovered during any of the excavations. Even the excavators noted that the small pieces of charcoal that were recovered could be secondary deposits. In addition, only two of 771 long bone, rib, and vertebrate specimens appear to have been calcined.
It is important to re-examine faunal collections that were excavated and interpreted before modern experimental and actualistic taphonomic approaches became available. This study of the Hanaizumi faunal assemblage serves this purpose, particularly in East Asia, where few taphonomic studies have been conducted. Additional taphonomic analyses of faunal assemblages from other regions of NE Asia are critical for the development of a clearer reconstruction of the nature of hominin-carnivore interactions over large game resources. Furthermore, they will also provide significant insights into hunter-gatherer processing and transport decisions of large game in this region of the Old World during the Late Pleistocene.
Actualistic and experimental taphonomic studies specific to NE Asian contexts are necessary, particularly due to variation in the Old World carnivore guild. For instance, hyenas were not present on the Japanese archipelago at any time during the Pleistocene (Kawamura, 1982), leaving only the large cats, wolves, and bears as the primary large carnivores that may have competed with hominins in the region. In order to overcome the lack of comparable data that address taphonomic questions specific to the NE Asian Plio-Pleistocene faunal record, C.J.N. is currently designing a series of actualistic and experimental studies to be conducted in NE China. In addition, comparative studies with the North American taphonomic record (e.g. Old Crow Basin) are warranted, since similarities in the carnivore guild composition between the two regions existed during the Pleistocene.
Study of the Hanaizumi materials has made several contributions to reconstruction of the paleoenvironment of the Japanese archipelago. For instance, Palaeoloxodon naumanni was discovered there, suggesting that Naumann’s elephant spread into that region of the Japanese archipelago before 20000 B.P. In addition, interpretations of the pollen recovered from the stratigraphic profiles have helped form the foundation of paleoenvironmental reconstructions of Pleistocene Japan. Furthermore, the series of chronometric dates has contributed to a solid reconstruction of the depositional history of the site and region.
Nevertheless, the faunal dataset has limited utility for addressing paleoanthropological debates. The intent here is not to minimize the work that Matsumoto et al. (1959a, b) contributed with their excavations at Hanaizumi. These excavations were, for the time (1950s), considered by many to be cutting edge. However, Pleistocene faunal assemblages that were excavated (and interpreted) before taphonomic research became commonplace need to be re-examined from a taphonomic perspective in order to determine whether the assemblage should be considered solely as a paleontological collection or whether it can contribute to paleoanthropological debates. We suggest that Hanaizumi is best considered an allochthonous accumulation of fluvial transported bison carcasses, at least some of which were procured and processed by hunter-gatherers.
C.J.N. would like to thank Sadako Takeuti, curator at the Saito Ho-on Kai Museum in Sendai, for permission to study the Hanaizumi assemblage stored there, and Hiroshi Kajiwara for logistical support during his stay at Tohoku Fukushi University also in Sendai. This study was supported by grants to C.J.N. from the US National Science Foundation (BCS 9907159), the Bigel Endowment for Research (Rutgers University), and the Center for Human Evolutionary Studies (Rutgers University). This manuscript was partially supported by the Major Basic Research Projects (2006CB806400) of MST of China. We would like to thank Rob Blumenschine, Susan Cachel, Mark Diab, Jack Harris, Lee Lyman, Akira Ono, Briana Pobiner, and one anonymous reviewer for many constructive comments on an earlier draft of this manuscript. C.J.N. takes full responsibility for any errors that may be present in this paper.
|