| Gen Suwa, corresponding author. e-mail: suwa@um.u-tokyo.ac.jp phone: +81-3-5841-2836; fax: +81-3-5841-8451 Published online 25 May 2007 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.061203 |
Homo erectus has been broadly defined to include fossils from Africa, Asia, and possibly Europe, or restricted to a supposedly confined Asian clade. The former view sees H. erectus as a polymorphic and polytypic species with intraspecific geographic and temporal variation (e.g. Howell, 1978; Bräuer and Mbua, 1992; Walker and Leakey, 1993; Rightmire, 1998, Rightmire et al., 2006; Asfaw et al., 2002; Dunsworth and Walker, 2002; Antón, 2003). According to this view, H. erectus is recognized by a common morphological pattern, consisting of a platycephalic cranium with developed supraorbital torus and strongly expressed ectocranial buttressing on a neurocranium with a cranial capacity usually greater than known in Homo habilis sensu lato and smaller than known in other archaic Homo (including Homo heidelbergensis/rhodesiensis). In this view, the available morphological evidence is interpreted to be consistent with regional (or continental) differentiation at the infraspecific taxonomic level, the taxon H. erectus qualifying as a paleospecies, a reproductively distinct reticulate species lineage (sensu Mayr, 1963) continuing through time. However, recent field and genetic observations suggest that reproductive isolation may not necessarily be absolute in higher primate and other mammalian species lineages (Grubb, 1999; Jolly, 2001; Tosi et al., 2003, 2005). When such interspecific gene flow is negligible, species lineages can still be considered to represent individually distinct evolutionary trajectories through geological time and can be recognized in the fossil record (e.g. Simpson, 1961; Wiley, 1981).
An alternative view would split the broader H. erectus hypodigm into several geographically and/or temporally segregated species level entities (e.g. Andrews, 1984; Tattersall, 1986; Wood, 1994; Wood and Richmond, 2000; Schwartz and Tattersall, 2003; Carbonell et al., 2005). This interpretation holds that subtle morphological indications represent phenotypic conditions that are the actual discrete evolutionary units informative of phylogenetic relationships. In such schemes, the nomen H. erectus would be (largely) restricted to the Asian hypodigm, usually represented by the Chinese (Lantien, Zhoukoudian, Hexian) and Javanese (Sangiran, Trinil, Sambungmacan) materials, although the Olduvai specimen OH 9 is commonly considered to be an African example of a predominantly Asian H. erectus (e.g. Wood, 1994; Wood and Richmond, 2000). Under this view, the earlier African remains are often attributed to Homo ergaster. One current usage of the latter taxon (Wood, 1991, 1992) considered it to be less specialized than H. erectus sensu stricto, and thus potentially ancestral to later Homo to the exclusion of H. erectus itself. Implicit in this interpretation is that both the H. ergaster lineage and the more specialized H. erectus examples with Asian affinities (such as suggested for OH 9) were represented in the early Pleistocene of Africa. Alternatively, other workers would have a single African lineage, including OH 9 in the same species as the H. ergaster material, in which case the nomen Homo leakeyi would have nomenclatural priority over “ergaster” (Clarke, 1994).
Recently discovered fossils attributable to a broadly defined H. erectus are allowing new insights into aspects of H. erectus evolution. The timing and place of H. erectus emergence must now be considered within an Afro-Asian perspective (Gabunia et al., 2000, 2002; Vekua et al., 2002; Rightmire et al., 2006), and the distinction between earliest H. erectus (or H. ergaster) of the ~1.8 to 1.5 Ma time range of the Turkana basin and H. habilis sensu lato may not be as clear as was once thought (Vekua et al., 2002; Leakey et al., 2003; Rightmire, 2006).
Geologically younger remains of African H. erectus or H. erectus-like remains have also recently been recovered in eastern Africa (Abatte et al., 1998; Asfaw et al., 2002; Potts et al., 2004). In particular, Asfaw et al. (2002, 2007) have shown that a H. erectus morphological pattern existed in Africa at ~1.0 Ma. They interpreted the available evidence to be most consistent with a chronosubspecies recognition of the earlier (the H. ergaster hypodigm of Wood, 1992) and later (Daka) African forms, and that distinct regional clades cannot be consistently retrieved from cladistic parsimony analysis (Asfaw et al. 2002; Gilbert et al., 2003; Gilbert, 2007). Others interpret the same evidence as exhibiting a continuous African lineage, but consider this lineage to be distinct from Asian H. erectus and perhaps not directly ancestral to later Homo (Manzi et al., 2003). Of relevance to such discussions is the actual tempo and mode of morphological evolution in the early Pleistocene of Africa (and elsewhere), and how this may relate to the morphological patterning seen in the Eurasian Pleistocene hominid record. However, the currently available African record predating 1.0 Ma is poor, consisting of the Turkana basin, Olduvai, and the more limited South African materials.
In 1988, the Ethiopian Ministry of Culture organized a project, the Paleoanthropological Inventory of Ethiopia (Asfaw et al., 1990, 1991, 1992; Suwa et al., 1991; WoldeGabriel et al., 1992), the aim of which was to identify and record new paleoanthropological sites within Ethiopia. More than 30 new sites were designated between 1988 and 1992, including Konso-Gardula (or Konso) at the southwestern end of the Main Ethiopian Rift. The Konso sites, initially discovered in 1991, have been further researched by the Konso Paleoanthropolgical Research Project, coordinated by the two of us (Y.B. and G.S.). This work has yielded a mammalian fauna abundantly represented between ~1.9 and 1.3 Ma (Suwa et al., 2003; White and Suwa, 2004) and early Acheulean archeological assemblages extending back in time to ~1.6 Ma (Beyene et al., 1997; Beyene, 2003). Hominid remains of both Australopithecus boisei and H. erectus were recovered (Asfaw et al., 1992; Suwa et al., 1997). All hominid remains come from stratigraphic intervals dated to between ~1.45 and ~1.3 Ma (Katoh et al., 2000; Suwa et al., 2003), and were probably slightly younger than the hominid remains of the upper sequences of the Koobi Fora and Nachukui Formations at Lake Turkana (WoldeGabriel et al., 2005; Gathogo and Brown, 2006; McDougall and Brown, 2006). The Konso hominid collection includes eight craniodental specimens that we here attribute to H. erectus (Table 1). These are fragmentary, but include KGA10-1, which is one of the better-preserved H. erectus mandibular specimens known from eastern Africa. The KGA10-1 mandible and one other molar were preliminarily described by Asfaw et al. (1992).
We consider this modest H. erectus assemblage from Konso to be a significant addition to the African early Pleistocene H. erectus hypodigm, which contributes to the characterization of the eastern African H. erectus lineage and their possible relationships not only with earlier and later African Homo but also with the Eurasian early Pleistocene hominid record, the latter being increasingly discussed recently (e.g. Antón, 2003; Kaifu et al., 2005b; Dennell and Roebroeks, 2005; Durband et al., 2005; Kaifu, 2006; Rightmire, 2006). The purpose of this paper is to provide a fuller description of the Konso H. erectus fossils in comparison with previously known African materials, and to evaluate the eastern African H. erectus lineage with the additional perspective that the Konso assemblage provides. Our aim is to provide a basic analysis of the Konso remains so as to make available the information and fossil material for further studies.
The Konso H. erectus specimens (Figure 1) and their chronological placements are summarized in Table 1. Further stratigraphic and chronological details are given elsewhere (Suwa et al., 2003; Nagaoka et al., 2005; WoldeGabriel et al., 2005). The comparative descriptive format employed in this work follows that of White et al. (2000). Mandibular and dental terminologies used in this work follow White (1977), Suwa (1990), and Suwa et al. (1996). Other terminologies, particularly those used by Weidenreich (1936) and Richards et al. (2003), are also mentioned where necessary. We followed the methods of White and Johanson (1982) and Wood (1991) for the mandibular, and White (1977), Suwa (1990), and White et al. (2000) for the dental measurements. The mandibular metrics of the Konso specimen and almost all early Homo and H. erectus dental dimensions were taken by ourselves [the exceptions being A.L.666-1 from Kimbel et al. (1997) and OH 65 from Blumenschine et al. (2003)], but we used the mandibular metric data available in the literature (Wood, 1991; Rosas and Bermúdez de Castro, 1999; Kaifu et al., 2005a). The comparative morphological assessments of the dentition are based on our own observations of Pliocene and Pleistocene materials in part discussed elsewhere (e.g. Suwa, 1990; Suwa et al., 1996; Asfaw et al., 1999; White et al., 2000).
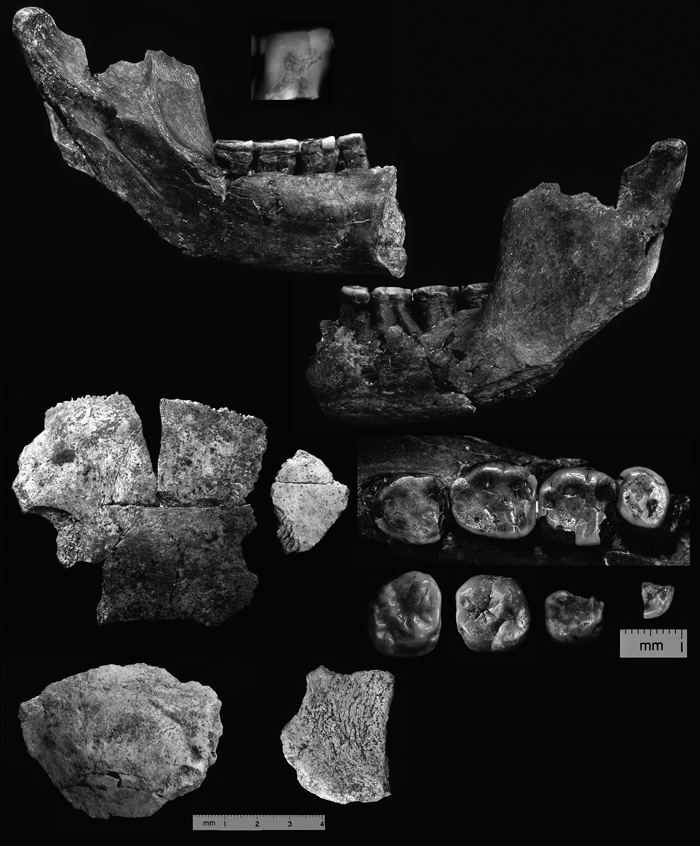 View Details | Figure 1. H. erectus remains recovered at Konso. Top right and left, KGA10-1 mandible (with enlarged lateral view radiograph of M1); middle left, KGA10-656 parietal and frontal fragment; bottom left, KGA7-395 occipital fragment; bottom middle, KGA10-620 parietal fragment; all to same scale. Middle right, occlusal view close up of dentition: top, KGA10-1 P4 to M3; bottom from left to right, KGA4-14 right upper M3, KGA11-350 left upper M1, KGA12-970 right upper dm2, KGA8-150 left lower P4 fragment; all to the same scale, buccal towards the top. |
We describe the Konso material with reference predominantly to the African early Pleistocene comparative material. This is to clarify the morphological affinities of the Konso material relative to the well-known ~1.5 to > 2.0 Ma eastern African early Homo and H. erectus records. These materials come from the Shungura, Koobi Fora, and Nachukui Formations of the Turkana basin, Kenya, and from Olduvai Gorge, Tanzania. The largest of these collections comes from the Koobi Fora Formation, which, after Wood (1992), is classically considered to represent multiple coexisting species lineages of Homo. However, we take the alternative position that this is not conclusively shown, even with the most recent stratigraphic and chronological revisions (Brown et al., 2006; Gathogo and Brown, 2006; McDougall and Brown, 2006). Therefore, we first outline our updated interpretations of the relevant Turkana basin early Homo and H. erectus materials. Following this comparative framework, the relevant dental specimens used in the quantitative analysis of the present study and their chronological estimates are listed in Table 2.
The Turkana basin early Homo fossils have been allocated to H. ergaster, H. habilis, H. rudolfensis, or Homo sp. indet. (Wood, 1991, 1992). We take the alternative approach that the H. habilis/rudolfensis hypodigm represents a single early Homo lineage evolving through time. We consider that this null hypothesis is yet to be falsified by the available evidence, as argued by Miller (1991, 2000) and Suwa et al. (1996). Scrutiny of the evidence also indicates that temporal overlap between H. habilis/rudolfensis and early H. erectus (H. ergaster) within the Turkana basin may be minimal or non-existing (see Suwa et al., 1996; and below).
The recently published stratigraphic and chronological revisions of the Turkana basin deposits have considerable impact on the chronological calibration of both the entire sequence (Brown et al., 2006) and on the estimated ages of the KNM-ER 1813 cranium and other hominid fossils recovered specifically from Area 123 (Gathogo and Brown, 2006). We continue to consider the stratigraphically lowest definitive occurrence of early H. erectus to be represented by the KNM-ER 3733 cranium (see Suwa et al., 1996). Others place a first appearance datum for the taxon on a small occipital fragment (KNM-ER 2598) at > 1.9 Ma, but this specimen was found on a desert-pavement lag surface that potentially samples overlying deposits now eroded away (White, 1996).
KNM-ER 3733 derives from the middle KBS Member and has been widely considered to be ~1.75 Ma (Feibel et al., 1989), but is more likely to be ~1.65 Ma as inferred from placement of marker horizon A2 (Brown et al., 2006), or as discussed by Gathogo and Brown (2006). According to previous stratigraphic accounts, the youngest secure H. habilis/rudolfensis specimens would be KNM-ER 1590 and 1805, which were reported to derive from the lower KBS Member (Feibel et al., 1989) with a suggested age of 1.85 Ma. However, the more recent chronological evaluations suggest a slightly younger age of ~1.8 Ma for KNM-ER 1590 [from placement of marker horizon C4 (Brown et al., 2006)], while KNM-ER 1805 is now suggested to derive stratigraphically from the upper KBS Member, closer to an unspecified Okote Tuff complex (Gathogo and Brown, 2006). We are wary of the latter newer allocation, which is so far not accompanied by the actual details of the revised stratigraphic interpretations. We thus prefer to consider KNM-ER 1805 to be stratigraphically ill-constrained, with a possible age of ~1.8–1.6 Ma.
Of further surprise is the revised date of KNM-ER 1813 and other specimens from Area 123, from the previous ~1.9 Ma to a much younger ~1.65 Ma estimate (Gathogo and Brown, 2006). If the latter point estimate of its chronology is accepted, then KNM-ER 1813 would be the youngest definitive occurrence of H. habilis/rudolfensis at Turkana, possibly contemporary with the H. erectus KNM-ER 3733 cranium. However, the ~1.65 Ma age is based on assumption of constant depositional rate for a composite section (apparently assembled from local sections > 5 km apart) that spans the KBS to Lower Ileret tuff intervals, the latter tuff occurring as a channel fill capping one of the local sections (Gathogo and Brown, 2006). From our field experience at Konso (Katoh et al., 2000; Nagaoka et al., 2005), time gaps of ~0.4 Ma may occur with no obvious erosional unconformities in comparison with sequences with a more continuous depositional record. Because of this, and because the Area 123 situation apparently lacks further chronological control, we take the conservative interpretation that KNM-ER 1813 and the other Area 123 fossils have an ill-constrained age of ~1.8–1.55 Ma. Finally, two other likely H. habilis/rudolfensis specimens, KNM-ER 3891 and 1814, with a general KBS Member attribution, are also stratigraphically ill-constrained, allowing a possible age range of ~1.85–1.6 Ma (Feibel et al., 1989; Brown et al., 2006).
We conclude that the recent chronostratigraphic revisions of the Turkana basin (Brown et al., 2006; Gathogo and Brown, 2006) fail to demonstrate the co-occurrence of H. habilis/rudolfensis with early H. erectus (contra Gathogo and Brown, 2006). What appears to be the case is that all Homo specimens that definitively derive from the lower portion of the KBS Member and lower stratigraphic levels either belong to H. habilis/rudolfensis or are not diagnostic at the species level. Conversely, all Homo specimens that derive definitively from the upper KBS Member and higher stratigraphic levels are either attributable to H. erectus or are not diagnostic at the species level. Finally, we consider KNM-ER 1805 and 1813 to belong to H. habilis/rudolfensis but at an ill-constrained age of ~1.8–1.55 Ma. These may or may not have overlapped in time with early H. erectus at ~1.7–1.6 Ma. For the purpose of analysis of the present study, we allocate these an arbitrary age of 1.7 Ma. Based on size and morphology, we also consider the KNM-ER 1814 dental set of ill-constrained age to represent H. habilis/rudolfensis, which also potentially overlaps with early H. erectus in time. This and two other dentognathic specimens, KNM-ER 1507 and 1508 deriving from Area 127, are given an arbitrary age of 1.7 Ma, although Wood (1991, 1992) attributed KNM-ER 1814 to sp indet. and 1507 to H. ergaster.
Preservation
KGA10-1 is a left mandible with most of the body and ramus preserved. The corpus is preserved from the canine position posteriorly but lacks the inferior corpus under the M3. The buccal alveolar margin has been mostly lost by damage from C through M2, but small segments of alveolar bone at or approximating the alveolar margin are retained at mesial P4 and mid M1 positions. The lingual alveolar margin is damaged at M3 but is otherwise intact. The symphyseal region is broken superiorly just anterior to the mesial wall of the canine alveolus. Basally, preservation extends more medially just at or close to the midline. Much of the ascending ramus and the neck and condyle are present, but the entire mandibular (gonial) angle region, the tip of the coronoid process and most of the mandibular notch are missing. The P4 and all molars are preserved, with some damage on M1 and M3, as detailed below. The alveolus of the P3 is preserved to allow some idea of root form. The surface detail of both bone and teeth is excellent.
Mandibular morphology
The corpus metrics of the KGA10-1 mandible are given in Table 3. Corpus dimensions and shape must be seen in light of the possible effects of alveolar bone pathology. The lingual alveolar bone is intact from M2/M3 to P4 position, and shows definite signs of periodontal resorption. Its alveolar margin is dislocated lingually from the molar roots by ~1–2 mm at mesial M2 and M1 positions. A 4 mm pocketing occurs at the P3/P4 lingual alveolus. However, corpus height measured lingually appears to be reliable, judging from the unaffected distal M2 and distal P4 alveolar margins. Most of the buccal alveolar margin is lost by damage anterior to M2. But a minute segment of the superolateral surface of the alveolar border is preserved at P4 position. This enables reliable estimates of buccal corpus height and contour, in congruence with the lingual alveolar position. It is inferred that the KGA10-1 corpus had roughly parallel superior and inferior borders under the molars, as seen in KNM-ER 992 and KNM-ER 730. However, it appears to lack the strong anterior corpus shallowing observed in the latter and other H. erectus specimens. The extensively exposed P4 and M1 root systems of KGA10-1 most likely indicate accelerated continuous eruption, possibly related to an idiosyncratic craniofacial form and occlusal relationships.
Corpus dimensions of KGA10-1 are comparable to the known Turkana basin and Olduvai H. erectus mandibles in size and shape. Corpus size at M1 is larger than in KNM-ER 730 and KNM-ER 992 (Wood, 1991), but the latter is larger than KGA10-1 at the M2 position. The robusticity index of KGA10-1 at the M1 position is slightly greater than that of KNM-ER 992, and at the uppermost range of variation of both H. habilis/rudolfensis and eastern African H. erectus mandibles (Wood, 1991).
The lateral corpus exhibits an expressed lateral prominence at M2/M3 position, making the corpus thickest at that position. Although the lateral prominence is well developed and distinct in KGA10-1, it is weaker and situated more posteriorly than the condition seen in KNM-ER 992. The extramolar sulcus of KGA10-1 is broad, measuring 9.0 mm at mid M3 (taken from ramus crest to alveolar margin). This is slightly broader than that reported for KNM-ER 992 (8.5 mm: Rightmire, 1990).
The lateral corpus surface of KGA10-1, anterior to the lateral prominence, is characterized by superior and inferior lateral tori. The superior lateral torus continues anteriorly from the lateral prominence, and fades into a shallow depression below P4. This depression is, in turn, bounded anteriorly by a bony swelling lateral to the P3 alveolus. A similar depression is expressed in OH 22 below the P3 level.
The inferiorly extending bony eminence of the lateral prominence forms the anterior border of the masseteric fossa. This ill-defined structure apparently continues to the inferior lateral torus at the basal corpus margin. The inferior lateral torus of KGA10-1 is indistinct, but bears the clear impressions of the platysma along the lateral corpus base. The platysmal impression extends anteriorly to the canine position, with a maximum superoinferior dimension of 12 mm under P4. No swelling of bone corresponding to the anterior marginal tubercle (e.g. Tobias, 1991; Bräuer and Schultz, 1996) occurs at the anterior end of the inferior torus. Below the two anterior molars a shallow depression, the intertoral sulcus, occurs between the two lateral tori. The intertoral sulcus of KGA10-1 is broad and shallow, unlike the more distinct conditions traditionally considered to be typical for H. erectus (e.g. Ternifine and Zhoukoudian materials). The intertoral sulcus ends anteriorly at the mental foramina in KGA10-1. The Turkana basin mandibles (e.g. KNM-ER 992) do not exhibit a definable intertoral sulcus.
Two large mental foramina occur below the P4/M1 position. The anterosuperior one is ~4 mm long, opens anterosuperiorly, and is placed approximately at mid-corpus height (its inferior rim is 6.5 mm above corpus base), below the distal-most P4 crown. The posteroinferior foramen, also ~4 mm long, opens posteriorly, and is placed under mesial M1 (its inferior rim is 14 mm above corpus base) at a position 3.5 mm inferior and 4 mm posterior to the anterior foramen. A 3.3 mm wide bony bridge running obliquely anteroinferiorly separates the two foramina.
Oblique striations occur on the medial aspect of the KGA10-1 alveolar border at M1 and M2. This feature has been described in the Zhoukoudian material by Weidenreich (1936), and is also seen in KNM-ER 730 (Day and Leakey, 1973) and KNM-ER 992. The medial corpus surface shows a submandibular fossa (subalveolar fossa) that is weaker and less defined than the condition seen in KNM-ER 992. In KGA10-1, an indistinct mylohyoid line occurs along a bulbous rounded eminence that forms the superior boundary of the submandibular fossa.
The submandibular depression extends anteriorly towards the midline to the genioglossal fossa, which defines and separates the weak superior transverse torus from a stronger inferior transverse torus of the symphyseal area. Small remnants of the genial spine are preserved in the genioglossal fossa. An inferiorly facing digastric fossa of 17 mm × 6 mm occurs anteriorly, adjacent to the inferior transverse torus. The symphyseal area appears to be thick, with the posterior surface of the inferior transverse torus placed at the P4/M1 position when viewed perpendicular to the occlusal plane. The superior transverse torus does not project as posteriorly, but is still placed at the P3/P4 position. The planum alveolare was apparently as developed as it is in KNM-ER 992.
The anterior border of the ascending ramus originates at the mesial M3 position, rather than at the mid-M3 position as it does in KNM-ER 992. The preserved anterior and posterior borders of the ascending ramus slope superoposteriorly relative to the occlusal plane or corpus borders. The orientation of the ascending ramus is much more vertical in KNM-ER 992. The height of the ramus measured from corpus base perpendicular to the condyle is ~72–75 mm. Condylar height is 43.5 mm above the occlusal plane. The minimum width of the ramus is 49.5 mm between the preserved anterior and posterior borders, narrower than it is in KNM-ER 992.
The masseteric fossa is strongly concave in its anteroinferior portion, more so than in KNM-ER 992. Two strong vertical ridges for intramuscular tendons occur at the mid ramal surface, and weaker ones occur more anteriorly. Posterosuperiorly towards the neck and mandibular notch, a localized concavity is formed on the lateral ramal surface. This fossa is delimited posteriorly by a rounded eminence running to the mandibular condyle, the crista ectocondyloidea of Weidenreich (1936).
The medial surface of the ascending ramus shows developed crista endocoronoidea and endocondyloidea that converge inferiorly to form the planum triangulare of Weidenreich (1936). However, in KGA10-1, topographical depth of the ramus is accentuated by damage at the preserved superior break, and the endocoronoid ridge is weaker than it is in KNM-ER 992. The two ridges merge anteroinferiorly to form the crista pharyngea, which is the posteromedial boundary of the buccinator gutter. The latter is a broad and hollow feature in KGA10-1, as it is in KNM-ER 992, but is more extended anteroposteriorly than it is in KNM-ER 992 [or the even more restricted condition seen in H. habilis (OH 13) and Australopithecus mandibles (e.g. Rosas and Bermúdez de Castro, 1998)].
The mandibular foramen is located approximately 42 mm anteroinferiorly to the condylar surface and 17 mm anterior to the preserved posterior medial ramus border. It is positioned approximately equidistant between the latter and the crista pharyngea, as it is in KNM-ER 992 and later examples of Homo, whereas Australopithecus and H. habilis (OH 13) tend to exhibit a mandibular foramen more closely situated to the crista pharyngea. The mylohyoid sulcus runs anteroinferiorly for 23 mm from the inferomedial margin of the mandibular foramen to a location ~15–20 mm posteroinferior to distal M3. It is inclined ~50° from the lingual alveolar plane. The mandibular lingula is broken but would have projected more than 4 mm posteriorly. A strongly expressed linea pterygoidea occurs as a distinct crest running for ~10 mm posterosuperiorly from the commencement of the mylohyoid sulcus. This crest becomes ill-defined posterosuperiorly, but appears to continue to a faint thickening of bone at the medial surface of the posterior ramus border, possibly a variant expression of the tuberculum sulcus colli of Richards et al. (2003). A deeply concave anterior component of the sulcus colli is defined between the linea pterygoidea and the crista endocondyloidea. The sulcus colli takes the form of an anteroinferior to posterosuperiorly directed linear concavity that rapidly shallows posterosuperiorly.
The posterior ramus border is preserved to a point ~40 mm below the condylar surface. On the medial neck surface at ~10 mm below the superior condylar surface there is a 10 mm × 2 mm fossa for the lateral pterygoid insertion running superoinferiorly. The tuberculum sulcus colli equivalent structure referred to above and a more prominent tubercle occur just above and below the posterior ramus border break, respectively. The latter takes the form of an 8 mm long, anteroinferiorly directed shelf on the medial ramus surface adjacent to the posterior ramus border. This bony eminence represents the superior portion of the septum 6 of the medial pterygoid insertion (terminology of Richards et al., 2003). The preserved medial ramus exhibits the anterosuperior ends of four additional ridges emanating from the missing gonial region. These are best interpreted as associated with septa 4, 2III, 2II, and 2I of the medial pterygoid muscle insertion.
The mandibular condyle measures 26.2 mm mediolaterally and 11.2 mm anteroposteriorly. Its superior surface is relatively flat laterally, but the medial portion is convex both mediolaterally and anteroposteriorly. The root of the mandibular notch is placed laterally at roughly a one-fifth of the distance between the condyle’s lateral and medial margins.
Dental morphology
Most of the crown and root of the P4 through M3 are preserved. Crown dimensions are given in Table 4. The KGA10-1 P4 abnormally erupted into a rotated position by approximately 90° (clockwise in occlusal view). Large interproximal facets for the P3 and M1 are formed on the anatomically buccal and lingual crown surfaces, respectively. Measurements and descriptions are given in anatomical orientation.
In absolute crown size, the postcanine dentition of KGA10-1 is comparable to the largest of the eastern African materials attributed to H. erectus (Figure 2). The M2 > M1 size gradient is shared with both early H. erectus and H. habilis/rudolfensis, but the size discrepancy between the two molars is smaller in H. erectus and KGA10-1 than it is in the other African early Homo remains. Compared to both H. habilis/rudolfensis and known early African H. erectus, the KGA10-1 M3 is distinctly reduced in size relative to M1, and the P4 is also relatively small. In KGA10-1, the shape index of the relatively diagnostic M1 is estimated to be ~105.5 (Table 4, Figure 3), thus not exhibiting the mesiodistally elongated shape characteristic of both early H. erectus and other early Homo M1s (usually above 110).
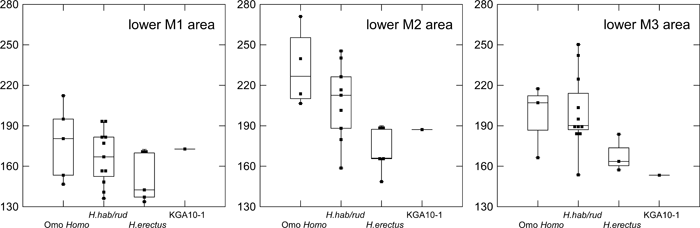 View Details | Figure 2. Mandibular molar crown areas (mesiodistal × buccolingual diameters in square mm). Omo Homo, Shungura Formation early Homo at > 1.9 Ma; H. hab/rud, Koobi Fora Formation and Olduvai H. habilis/rudolfensis (~1.9–1.65 Ma); H. erectus, early African H. erectus (~1.65–1.5 Ma) (see text for further information on taxonomic attributions). Box plots show the median (horizontal line), the range of the central 50% values (box margins), values within the inner fence (1.5 times the box range from the box margin) joined by a vertical line, and further outliers. Each filled square represents a specimen. |
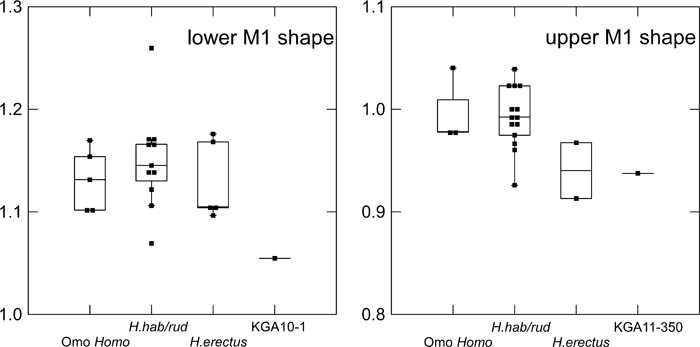 View Details | Figure 3. Molar crown shape (ratio between mesiodistal and buccolingual diameters). Omo Homo, Shungura Formation early Homo at > 1.9 Ma; H. hab/rud, Koobi Fora Formation and Olduvai H. habilis/rudolfensis (~1.9–1.65 Ma); H. erectus, early African H. erectus (~1.65–1.5 Ma) (see the text for further information on taxonomic attributions). Box plots as in Figure 2. |
Parts of the P4 crown and root are broken at its distolingual portion. The crown is worn flat with virtually no cusp topography remaining. A large dentine exposure is seen on the P4 protoconid (2.7 mm × 1.3 mm), and a smaller 0.5 mm diameter exposure occurs on the metaconid. A premortem macrofracture of the enamel is seen at the anatomical distolingual corner of the talonid adjacent to the mesiobuccal M1 corner. The occlusal view outline of the P4 crown was probably an irregular oval with obtuse mesiolingual corner and an evenly rounded distal contour. From the dentine exposures and what remains of the fissure pattern, the following crown morphology is inferred. The metaconid is positioned directly lingual to the protoconid, and a transverse crest interrupted the longitudinal groove. A small anterior fovea occurs mesially, and a larger fovea is formed in the talonid where the longitudinal groove terminates and the other secondary fissures converge. Two oblique occlusal ridges occurred on the distal occlusal surface of the metaconid, and a single comparable ridge is discernible on the distal occlusal slope of the protoconid. At least one crenulation incised the distal marginal ridge, and the distolingual corner was delineated by a groove. However, dentine horn prominence at the distolingual crown is minimal, so that the P4 is inferred to have lacked a distinct distolingual cusplet. The talonid was most likely weakly expressed and ill-demarcated from the main cusps (contra H. habilis/rudolfensis). It probably consisted of a thick and crenulated distal marginal ridge, as is seen in the slightly less worn KNM-ER 992. The P4 root, observed within the alveolus, appears to be single, with a broad, flattish, oval cross section of 11.5 mm × 6 mm at the alveolar margin level. A single large root canal was observed before the P4 fragments were glued together. No trace of a groove exists on its convex mesial surface, contra a Tomes root pattern. The P3 also probably possessed a single flattish root, as inferred from its preserved alveolus. Thus, the KGA10-1 P4 exhibits the more progressive morphological condition within the early Homo/early H. erectus range of variation in both crown and root morphology.
The M1 is mostly well preserved, but lacks its mesiolingual crown corner and lateral enamel of the entoconid. A premortem macrofracture occurred at the mesiobuccal protoconid position. The estimated extent of the original breakage is ~2–3 mm on the M1, continuous with a 1 mm diameter chipping of the adjacent P4 crown. The then-fractured surfaces and sections were affected by subsequent occlusal wear. Aside from this, the M1 crown is flatly worn, with very little cusp topography retained, only in the metaconid. All five main cusps are present, and exhibit dentine exposures that are more extensive in the buccal cusps (protoconid 3.5 mm × 3.0 mm, hypoconid 2.1 mm × 1.7 mm, hypoconulid 1.8 mm × 1.5 mm) and smaller in the lingual cusps (metaconid not measurable, entoconid 1.0 mm diameter).
The occlusal view outline of the KGA10-1 M1 crown is relatively rectangular, as is common in early Homo and some H. erectus M1s. The relatively squared mesiolingual corner (inferred) is associated with a metaconid apex situated slightly distal to the crown corner itself. The buccal crown contour is relatively straight, with only weak constriction of the contour between protoconid and hypoconid. Distal projection of the hypoconulid is weak. The metaconid is large mesiodistally, amply contacting (1.5 mm) the hypoconid to form a clear Y5 fissure pattern. A small deflecting wrinkle is likely to have been present, suggested by a longitudinally directed secondary fissure. Likewise, a small C7 or postmetaconulid probably existed, judging from the short but transversely oriented secondary fissure, the corresponding indentation on the lingual crown face, and the distally deflected lingual segment of the main transverse groove. Distally, a small remnant of the original posterior fovea is expressed as a minute pit retained between the worn entoconid and hypoconulid. The distally broken enamel exposes the dentine core of the distal marginal ridge, showing a plain contour that suggests the lack of a definable C6, as in most early Homo and H. erectus M1s. The root system of the KGA10-1 M1 is characterized by a low position of mesial and distal root furcation, at approximately 10 mm and 8 mm below the lingual and buccal cervical lines, respectively. Corresponding to this, its pulp chamber is superoinferiorly tall, but also exhibits an anteroposterior constriction (Figure 1), unlike later Pleistocene examples of taurodont molars (e.g. Skinner and Sperber, 1982). The mesial root is slightly larger (buccolingually broader and anteroposteriorly thicker) than the distal root and is directed relatively perpendicular to the crown. The distal root slopes more inferodistally. The KGA10-1 M1 is thus morphologically conservative amongst the early Homo and H. erectus M1s in crown contour and in cusp position and fissure patterns. It differs from early Homo and early African H. erectus in its relatively broad crown shape.
The M2 is almost complete, with little damage to its roots and cervical region. Occlusal wear has created a predominantly flat occlusal surface, with minimal cusp topography retained in the metaconid. The occlusal wear surface of the M2 consists of two adjacent, weakly concave surfaces, a smaller mesial and larger distal one on the talonid. The two surfaces are delineated and separated by the weakly projecting metaconid and a relatively high area of the worn occlusal surface at the protoconid-hypoconid junction. Although no major premortem damage is seen on the M2, enamel chipping and subsequent wear is inferred at the mesiobuccal occlusal corner, where a tiny worn pit straddles the occlusal and lateral crown surfaces. All five main cusps can be discerned. Dentine exposure is large at the protoconid (4.7 mm × 2.7 mm) and pinhole sized (0.5 mm diameter or less) at the hypoconid, metaconid, and entoconid. The remaining fissure and groove patterns attest to the presence of the hypoconulid, but no dentine exposure is seen.
In occlusal view, the M2 crown exhibits an angular mesiolingual corner, as inferred for the M1. It differs from the M1 in its more rounded buccal contour that tapers distally toward the distally extended hypoconulid. Thus, the M2 crown appears more rounded than the M1. The remaining occlusal fissures indicate that, as with the M1, this molar had a Y fissure pattern, with over 2 mm of hypoconid-metaconid contact. The presence of a metaconid deflecting wrinkle is inferred, but the possible presence of C7 and/or postmetaconulid cannot be assessed, because occlusal wear at this particular crown portion is more advanced than in the M1. The hypoconulid is situated more buccally and distally than in the M1, and the posterior fovea shows fissure branching that suggests the presence of a small C6. As with the M1, no evidence of a protostylid complex is seen on the buccal crown surface, although a weak furrow/depression just mesial to the M2 mesiobuccal groove may suggest some form of its expression in an unworn condition. The root system is similar to that of the M1 in showing a low position of furcation of the larger mesial and slightly smaller distal roots. Both roots are directed slightly distally and without the extreme distal root flare seen in the M1. The M2 of KGA10-1 is large in size and can be considered to be a morphologically conservative example among early Homo and H. erectus M2s in its well-developed metaconid forming a Y fissure pattern, and in its developed distal cusps.
The M3 crown is damaged over most of its mesial face. There is also a small area on the distal face that lacks enamel. A ~2 mm premortem macrochip with subsequent wear is observed mesiobuccally. Occlusal wear is generally flat, but wear topography includes weak metaconid saliency and a shallow concavity dominating the occlusal surface. The latter surface slopes distolingually, forming a helicoidal relationship with anterior M3 or the M2. Only the protoconid shows a dentine exposure (2.2 mm diameter), but in addition to the discernible five major cusps, fissure remnants suggest the presence of a C6 (or C6 area). However, dentine topography along the hypoconulid to C6 area must have been minimal, judging from the non-perforated dentinal apices and flat topography of the exposed dentine core. Thus the distal M3 crown was probably a crenulated area without distinct cuspal expression. In occlusal view, both lingual and buccal crown contours are evenly rounded with weak distal elongation and/or tapering of the crown. Very limited contact between the protoconid and entoconid indicates an X fissure pattern (but close to a + condition). On the buccal crown face, an oblique furrow runs mesially adjacent to the buccal groove. This suggests that the protostylid complex was somewhat expressed in the M3. The root system of the KGA10-1 M3 is not preserved distally, and cannot be described adequately within the alveolus. It appears to exhibit juxtaposed or fused mesial and distal root portions. This M3 thus shows morphological features associated with M3 reduction, such as its crown contour and cusp/fissure patterns. A similar crown shape and helicoidal wear pattern is seen in the KNM-ER 730 M3.
Enamel thickness was measured at appropriate natural sections, where radial thickness (Kono et al., 2002; Kono, 2004; Suwa and Kono, 2005) could be estimated for lateral crown faces. Lateral enamel thickness of the mesial and distal P4 faces was 1.3 mm and 1.2 mm, respectively. Lingual P4 enamel was at least 1.5 mm (at the interproximal facet position). Lingual M1 enamel at the metaconid was 1.4+ mm (thicker than 1.4 mm measured at the worn mesial break). The buccal enamel thickness of the M2 (natural fracture going through mid hypoconid) was 1.8+ mm as worn. Thus, the enamel of the KGA10-1 postcanine teeth appears to have been moderately thick, and more so buccally.
Interproximal grooves occur from mesial P4 to M3. The superoinferior height of the grooves is 1.5 mm at M2/M3, ~2–3 mm at P4/M1, ~2–4 mm at M1/M2, and 5 mm at mesial P4. The groove is strongest at P3/P4 and M1/M2 positions, corresponding to the areas affected by the periodontal disease. Buccolingually oriented coarse striations were observed under a low-powered microscope. These observations suggest that interproximal grooves were formed by intrusive material, perhaps related to irritation caused by periodontal disease.
KGA4-14 is an erupted maxillary right third molar crown with minimal wear. It has a small occlusal facet on the protocone tip and adjacent mesial marginal ridge, but lacks faceting on the other cusps. There is a faint and small interproximal facet placed 1.5 mm below the mesial marginal ridge.
The crown dimensions of KGA4-14 are given in Table 4. In crown size, this M3 lies at the smaller end of the H. habilis/rudolfensis range of variation, and is larger than the two measured African early H. erectus upper M3s from Turkana (Figure 4). The shape index of KGA4-14 is 87.6, well within the range of variation of the eastern African comparative samples. Later H. erectus M3s often exhibit a mesiodistally reduced crown, resulting in lower shape index values.
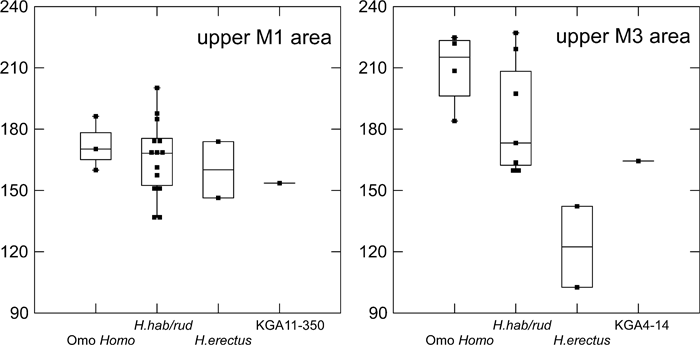 View Details | Figure 4. Maxillary molar crown areas (mesiodistal × buccolingual diameters in square mm). Omo Homo, Hadar and Shungura Formation early Homo at > 1.9 Ma; H. hab/rud, Koobi Fora Formation and Olduvai H. habilis/rudolfensis (~1.9–1.65 Ma); H. erectus, early African H. erectus (~1.65–1.5 Ma) (see the text for further information on taxonomic attributions). Box plots as in Figure 2. |
The occlusal view crown contour of KGA4-14 is rhomboidal, with relatively straight and distally converging buccal and lingual crown contours. The small distal cusps result in the strong obliquity of the buccal crown contour, unlike the more full and curved contour common in pre-2 Ma hominid M3s. The rhomboidal crown outline is seen in KNM-ER 807 and other early Homo examples.
In KGA4-14, the metacone is small, especially in its mesiodistal dimension, but forms a distinct, occlusally projecting cusp that defines an obtuse but distinct buccal notch. The hypocone is considerably smaller, and occupies the distolingual corner of the crown, only weakly differentiated from the distal marginal ridge. Its occlusal expression is reduced to a low apex with no occlusal crest/ridging. It projects occlusally from the lingual notch by a mere 0.5 mm. On the lingual crown face, the hypocone is separated from the protocone by a weak lingual groove that fades at mid-crown level. Reduction of the M3 hypocone is characteristic of H. erectus and later Homo M3s (often seen in extreme form in modern Homo sapiens), whereas pre-2 Ma hominid upper M3s with distal crown reduction tend to have larger hypocones and smaller metacones, and the lingual groove tends to be better developed and deeper than seen in KGA4-14.
Occlusally, mesial to the main ridges of the protocone and paracone, an epicrista occurs and forms the distal boundary of the anterior fovea. The latter is represented buccally by a deep, curved groove and lingually by a shorter shallow depression. The anterior fovea is contiguous with the longitudinal groove, which completely separates the buccal and lingual segments of the epicrista. The two segments of the epicrista run slightly obliquely distally, rather than being aligned straight transversely. The epicrista is a conservative trait commonly seen in early Homo and other Pliocene hominid M3s, while the discontinuity of the crest seen in KGA4-14 reflects a tendency towards its structural reduction.
The lingual segment of the crista obliqua commences from just distal to the protocone apex and curves distobuccally towards the metacone. This crest continues across the longitudinal groove to the metacone apex, notched but not incised by the longitudinal groove. Such a continuous crista obliqua is a conservative feature often seen in H. habilis/rudolfensis and other pre-2 Ma hominid M3s.
The distal marginal ridge exhibits two accessory cuspules. The metastyle occurs at the buccal-most distal marginal ridge, and a small distoconule occurs at the mid-distal marginal ridge position. The mesial marginal ridge is simple and featureless, without crenulation or formation of cuspules. Aside from the occlusal features referred to above, only weak surface topography occurs, so that KGA4-14 is not excessively crenulated. The degree of occlusal crenulation seen in KGA4-14 is comparable to or weaker than the condition seen in other early Homo specimens such as OH 13 and OH 24.
On the lingual face of the protocone, the Carabelli’s feature is expressed as a mesiolingual pit, with associated triangular enamel thickening and four smaller enamel swellings that run superoinferiorly at mid-crown height.
The cervix is preserved bucally and distally, and crown height above the cervical line is 8.5 mm, 7.5 mm, and 7.0 mm, at the paracone, metacone, and hypocone cusp tips, respectively. The distal marginal ridge is 5.5 mm high at its lowest position. The root is not preserved.
KGA11-350 is a complete crown of an erupted maxillary left first molar with little wear. Small interproximal facets occur on both mesial and distal crown faces, and all four major cusps are worn and facetted, indicating that the M2 had already erupted in this individual. None of the cusps shows dentine exposure.
The crown dimensions of KGA11-350 are given in Table 4. In crown size, this M1 lies within the smaller half of the H. habilis/rudolfensis range of variation (Figure 4). The shape index of KGA11-350 is 93.8, lower than seen in H. habilis/rudolfensis (circum 100), indicating a relatively broader crown (Figure 3). KNM-ER 807, one of the two early H. erectus upper M1 measured, also has a low shape index.
The occlusal view crown contour of KGA11-350 is almost square, with a weakly oblique buccal contour and a rounded distolingual corner. All four major cusps are well differentiated, and the metacone is larger than the paracone, as is usually the case in M1s as opposed to the posterior molars. The buccal groove demarcating the two cusps on the buccal crown face is weak, and a stronger groove separates the protocone and hypocone on the lingual crown face. On the mesiolingual face of the protocone, the Carabelli’s feature is expressed as a simple mesiolingual pit situated close to the occlusal surface. On the buccal crown face, the parastyle is expressed by a fine vertical groove occurring at the mesial end of the buccal crown face.
The anterior fovea region is occupied lingually by a worn, swollen enamel area. This structure is located adjacent to the Carabelli’s feature, and if distinct would be called the protoconule. Because this structure extends distally, there is no space for the lingual segment of the epicrista. The epicrista homologue is expressed buccally, and consists of a thin ridge of enamel that runs from the mesiobuccal occlusal corner obliquely distolingually. The crista obliqua commences from the protocone apex and curves distobuccally towards the metacone. This crest continues to the metacone apex, notched but not incised by the longitudinal groove.
The distal marginal ridge exhibits fine wrinkling but no distinct cuspules. Crown height there is 6.8 mm from the cervical line. Crown height at the little worn mesial marginal ridge is 6.6 mm. The root is not preserved sufficiently for description, but appears to have had the usual three roots.
KGA12-970 preserves most of the crown, but lacks a substantial portion of the paracone and much of the buccal face of the metacone. The preserved crown is broken just above the cervix. Crown height is low, as evinced by the slanting lingual protocone profile in mesial view. Wear occurs in all cusps, with dentine exposures of 0.5 mm and 0.3 mm in diameter at the metacone and protocone, respectively. The paracone tip is not preserved, and no dentine exposure occurs on the hypocone. Small interproximal facets occur on both mesial and distal crown surfaces.
The mesiodistal dimension of KGA12-970 is given in Table 4. It is smaller than the dm2 of OH 39. The latter is the smallest-toothed individual of the small early Homo comparative sample, but KGA12-970 is smaller in size. In occlusal view, KGA12-970 exhibits a tight distolingual crown contour, due to the lingually projecting hypocone position. The hypocone itself is well expressed but smaller than the metacone. It is separated from the protocone by a deep groove both occlusally and on the lingual crown face.
A continuous epicrista delimits a buccolingually directed, slit-like anterior fovea. The longitudinal groove notches, but does not incise, the epicrista into a long and linear buccal segment and a very short lingual portion. The crista obliqua consists of an occlusally concave crest running from the metacone cusp tip to the distal protocone crest at a position distal to the protocone tip. The longitudinal groove notches but does not incise the crista obliqua. The distal marginal ridge appears to be thick but simple without distinct cuspules, and terminates at the distoinferior end of the distal metacone crest. The Carabelli’s complex is represented by two short vertical grooves at the mesiolingual crown face and an enamel thickening between the two grooves. The lateral enamel thickness on the mesial face break is 0.7 mm above the interproximal facet.
KGA8-150 is a small premolar or molar fragment with a preserved dimension of only 7 mm. It is likely to be a mesiolingual fragment of a lower P4. Another positional candidate would be the distobuccal corner of an upper molar, but the crown height is too low and the exposed root canal structure is incongruent. We therefore consider this to be a fragment of left lower P4.
The lingual half of the mesial face is preserved and shows a portion of the mesial interproximal facet. The mesiolingual crown contour of this P4 was somewhat obtuse in occlusal view. The metaconid is small and its saliency is limited. Such morphologies correspond better with H. erectus P4s showing relative simple talonid expression, rather than with early Homo homologues with well-differentiated talonids. Lateral enamel thickness at the distal metacone face is moderately thick at 1.5 mm.
Preservation
This specimen consists of a large part of a right parietal and a small fragment of the frontal that joins to the parietal at the coronal suture. The smooth surface texture, thin bone, and complex and open sutures suggest that the skull fragment belongs to a young individual.
The parietal preserves an almost complete coronal border (a small segment of the inferior-most corner appears to be broken off), but lacks the entire lambdoidal border. The maximum anteroposterior cord and arc lengths of the preserved parietal squama are approximately 75 mm and 85 mm, respectively. The maximum superoinferior cord and arc lengths are approximately 70 mm and 80 mm, respectively. Along the sagittal border, a 10 mm (maximum length) × 30 mm wedge-shaped portion of bone is missing from the middle section. From the posterior part of the parietal towards the lambdoid border the bone has suffered pre-fossilization damage, as seen from the scalloped bone breakage. The latter suggests that the damage took place while the bone was still fresh. The edge of the break is uneven, with the external and the internal bone plates broken at different positions; the preserved internal bone plate extends maximally ~10 mm more posteriorly than the external plate. There is an approximately 10 mm × 20 mm area of bone missing at the preserved posteroinferior corner, at the squamosal border, which represents post-fossilization damage. The external bone surface is marked by modification in the form of fine and shallow grooves. These occur as sets of striae, concentrated mostly in the lower half of the parietal squama.
Morphology
The ectocranial surface is smooth, with no sign of the temporal line. On the posterior preserved part close to the sagittal border parasagittal flattening is seen that extends anteriorly for ~20 mm from the posterior break. Parasagittal flattening is a feature consistently seen in adult H. erectus. However, in adults the flattening covers most of the area between lambda and bregma. The presence of localized flattening in the KGA10-656 juvenile parietal suggests that, ontogenetically, parasagittal flattening may start posteriorly and extend anteriorly as the individual matures. The existence of such a growth pattern needs to be confirmed by further evidence. The parietal squama is well-curved in the coronal plane, as indicated by the arc and chord dimensions reported above. The squamosal border has a gentle curve, which suggests that the parietal at this stage of growth was not rectangular in shape, but rather narrow inferiorly and wider superiorly, a condition seen in young individuals. The striae parietalis are very localized and confined to an area adjacent to the squamosal suture. They do not extend superiorly, suggesting minimal overlap of the parietal and the temporal bones, characteristic of Homo. On the contrary, Australopithecus boisei parietals, even at a young age, exhibit striae parietalis that extend superiorly, revealing a more extensive area of overlap of the two bones.
The thickness of the bone is given in Table 5. The broken edge shows minimal development of the diploe, so that bone thickness is formed almost entirely from the external and internal bone plates. In endocranial aspect, the groove for the superior sagittal sinus is well marked from a position ~10 mm posterior to bregma and further posteriorly. A single parietal foramen is visible along the sagittal sinus ~30 mm posterior to bregma. The impressions of the meningeal vessels are well-marked. The anterior and posterior rami are strong. The anterior ramus does not follow the coronal margin, but rather takes a superoposterior direction away from the coronal border. Arising from the same sphenoid corner as the anterior ramus, there are three branches that form the posterior rami complex. The posterior two of these are strongly marked and course posteriorly close to the squamosal border before curving superiorly. The posterior-most branch is truncated by the bone break, but the anterior one has its full course preserved. It extends to the superior parietal area, close to the sagittal sulcus.
This is a small fragment of the posteroinferior part of a right parietal, approximately 45 mm × 35 mm as preserved. The specimen preserves small segments of the lambdoidal and parietomastoid sutures at the asterionic region. Although these sutures are open, the overall thick bone and apparently dense cortex suggests that this specimen may represent an adult or near-adult individual. Parietal thickness at asterion and comparisons are given in Table 5. The external surface exhibits a mildly developed angular torus, much smaller than that observed, for example, on the Bodo parietal (BOD-VP-1/1) (Asfaw, 1983) or characteristic of Asian H. erectus; its angular torus development is comparable to the Daka and OH 9 conditions. About 8 mm anterior to the angular torus a faint palpable expression of the supramastoid crest extension is seen, in contrast to the well-developed crest on the Bodo parietal. This is another similarity with Daka and OH 9, which exhibit a weak or apparent lack of expression of this feature (Asfaw et al., 2007).
This is a fragment of a right occipital, preserved from close to midline laterally to the asterionic region. The asterion to approximately the midline dimension is 61.5 mm, similar in size to KNM-ER 3733 and Daka, but smaller than OH 9. At asterion, it preserves ~10 mm of the right lambdoidal suture and 20 mm of the occipitomastoid suture, which were not fused in their preserved portions. Internally it preserves the right transverse sulcus, a small segment of the sagittal sulcus, and parts of the upper and lower cerebral fossae. The surface bone of the cruciform eminence region at the midline is exfoliated. Externally, the bone surface is rugged both sub-periosteally and at the muscle attachment areas. Because of this and the generally thick bone, this specimen may represent an adult or near-adult individual.
The superior and inferior nuchal lines are visible from medial to lateral extents. The superior nuchal line is more marked throughout its course, and forms the inferior border of a very weakly expressed occipital torus homologue, demarcated by a shallow supratoral sulcus. At ~20 mm from asterion, this divides into two branches, one extending horizontally towards just below asterion, and the other curving inferolaterally. When we compare the occipital torus and superior nuchal lines of KGA7-395 and the Daka and OH 9 occipitals, we see a strong crest-like morphology in Daka, a stronger torus-like expression medially in OH 9, and the weakest structure in KGA7-395. The lower occipital scale below the torus is convex as it is in OH 9. Occipital flexion is difficult to evaluate from the limited preservation. However, from the existing curvature lateral to the midline, a flexion comparable to or weaker than OH 9 and Daka can be inferred; this is less flexed than in examples of Asian H. erectus (e.g. Sangiran and Zhoukoudian). The thickness of the occipital at the preserved lambdoid border is 11.4 mm, and the maximum thickness along the occipitomastoid border is 12.2 mm.
The comparative descriptions presented above show that dentognathic morphological differentiation between Homo habilis/rudolfensis and early African H. erectus is incomplete, with large amounts of overlap in ranges of variation. This being stated, it is instructive to summarize the taxonomic and/or temporal morphological trends that are most evident. Relative to the H. habilis/rudolfensis condition, early African H. erectus from Olduvai and the Turkana basin is characterized by a tendency for smaller, less robust jaws and advanced posterior molar reduction (e.g. Wood, 1991; Bräuer and Schultz, 1996). As a part of a more robust morphology, H. habilis/rudolfensis mandibles tend to show a better-developed lateral prominence situated more anteriorly. Rosas and Bermúdez de Castro (1998) and Kaifu et al. (2005a, b) discussed the possible association of these and other corpus morphologies with a relatively narrow dental arcade and resultant corpus to ramus spatial relationships. The large postcanine dentition of H. habilis/rudolfensis tends to be expressed more explicitly in the two posterior molars (Figure 2, Figure 4). When some of the relevant dental metrics are plotted by taxon and time (Figure 5, Figure 6, Figure 7), it is suggested that posterior molar reduction is perhaps the most obvious easily quantified dentognathic change correlated with the H. habilis/rudolfensis to H. erectus transition.
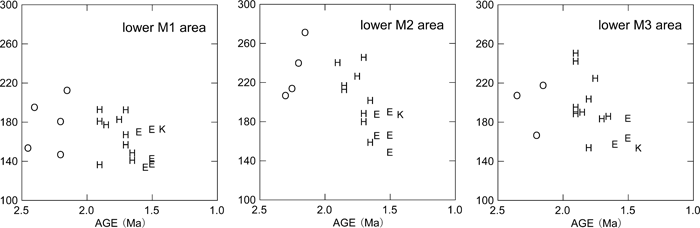 View Details | Figure 5. Mandibular molar crown areas (mesiodistal × buccolingual diameters) plotted against estimated chronological age. K, Konso; O, Shungura Formation early Homo; H, Koobi Fora Formation and Olduvai H. habilis/rudolfensis; E, early African H. erectus (see the text for further information on taxonomic attributions). |
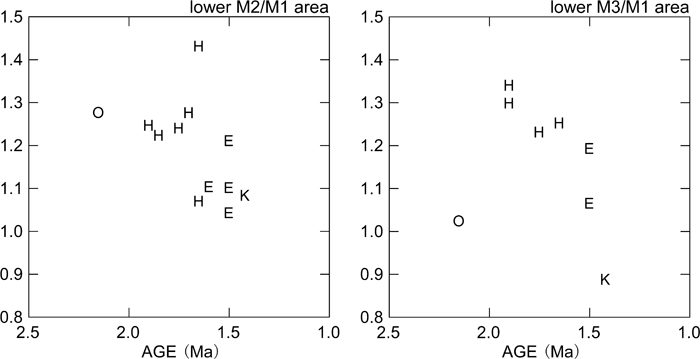 View Details | Figure 6. Mandibular molar relative sizes (M2 and M3 areas divided by M1 area) plotted against estimated chronological age. K, Konso; O, Shungura Formation early Homo; H, Koobi Fora Formation and Olduvai H. habilis/rudolfensis; E, early African H. erectus (see the text for further information on taxonomic attributions). |
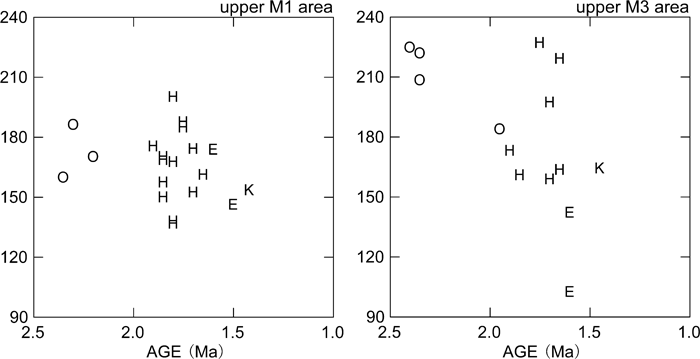 View Details | Figure 7. Maxillary M1 and M3 crown areas (mesiodistal × buccolingual diameters) plotted against estimated chronological age. K, Konso; O, Hadar and Shungura Formation early Homo; H, Koobi Fora Formation and Olduvai H. habilis/rudolfensis; E, early African H. erectus (see the text for further information on taxonomic attributions). |
The Konso hominids described above show a mosaic of primitive and derived morphological features. In mandibular and dental size, KGA10-1 occupies the largest end of the known African early H. erectus range of variation. Together with its thick corpus, which results in a high robusticity index, the KGA10-1 mandible appears distinctly more primitive than many examples of H. erectus sensu lato, especially those postdating 1 Ma (e.g. Bräuer and Schultz, 1996; Rosas and Bermúdez de Castro, 1998; Kaifu et al., 2005b). Some aspects of molar occlusal morphology detailed above also suggest a conservative condition broadly comparable to that of H. habilis/rudolfensis and African early H. erectus. In other features, the KGA10-1 mandible exhibits a condition more advanced than characteristic of H. habilis/rudolfensis. For example, development of the lateral prominence is weaker than in most H. habilis/rudolfensis mandibles or in KNM-ER 992. The KGA10-1 premolar crown and root exhibit the relatively reduced morphological condition, and the posterior two molars are reduced absolutely and relatively to M1 size. With M3 size, KGA10-1 appears to take postcanine reduction further than seen in the small Turkana basin early Homo and H. erectus mandibular samples currently available. Thus, attribution of the KGA10-1 mandible to the eastern African H. erectus lineage appears justified. The other Konso dental specimens are also compatible with an early H. erectus attribution. These teeth exhibit size overlap with both H. habilis/rudolfensis and early H. erectus, while showing a mixture of conservative and advanced morphological details of occlusal morphology. The information available from the fragmentary Konso cranial remains also suggest morphological similarities with known eastern African H. erectus examples that span the ~1.65 to 1.2 Ma time period. These include weak angular and occipital tori, and an apparently weak occipital flexion, when compared with Asian H. erectus.
Finally, in some features, the Konso specimens exhibit morphologies not necessarily characteristic of eastern African early H. erectus. The relatively narrow upper and lower M1 shape characteristic of early Homo is also present in early H. erectus of eastern Africa (Tobias, 1991; Wood, 1991; Suwa et al., 1996), but this was not seen in either of the available upper or lower Konso M1s (Figure 3). Another feature of relevance is the intertoral sulcus of the KGA10-1 mandible, which is not as localized and distinct as usually seen in later H. erectus (e.g. Tighenif and Zhoukoudian materials). This and the occurrence of two large mental foramina are not seen in the other known eastern African early H. erectus mandibles.
The above summarized cranial and dentognathic evidence of the Konso collection are consistent with the interpretation that H. habilis/rudolfensis and early H. erectus can be distinguished by some differences in central morphological tendencies, and that within the ~1.65 to 1.0 Ma period, the African H. erectus populations show evidence of morphological continuity. The Konso material is largely indistinguishable from the ~1.65 to 1.5 Ma eastern African H. erectus remains represented by the Turkana basin fossils, although, as seen above, some features are possibly indicative of more advanced and/or distinct modal conditions. These include mandibular intertoral sulcus expression and double mental foramina, which are a part of the classic characterization of Zhoukoudian H. erectus (Weidenreich, 1936), also shared by some of the Tighenif (Arambourg, 1963), Dmanisi (Bräuer and Schultz, 1996; Gabunia et al., 2002), and Sangiran mandibles (Kaifu et al., 2005a). Compared to the post-1 Ma African and Asian fossils of Homo, the Konso remains are distinctly conservative in mandibular robusticity and in some details of dental morphology. The Konso evidence supports a hypothesis of relatively little morphological evolution in African H. erectus prior to 1.4 Ma and further dentognathic gracility occurring sometime thereafter.
Recently, Rightmire et al. (2006) described in detail the morphology of the ~1.7 Ma Dmanisi crania, which combine derived H. erectus features with primitive H. habilis-like retentions. Similarly, Kaifu et al. (2005a, b) demonstrated that the chronologically earlier Javanese H. erectus dentognathic remains [with uncertain dates of anywhere between > 1.6 and ~1.0 Ma (e.g. Matsu’ura et al., 2006)] exhibit morphologies intermediate between the H. habilis/rudolfensis and African early H. erectus conditions. If these occurrences reflect populational affinities rather than mere homoplasy, the earlier Asian H. erectus populations were most likely established at the earliest stages of H. erectus emergence, prior to the full development of the African early H. erectus morphology. However, Asian populations derived from an African ancestry probably experienced modal shifts stemming from genetic drift, including founder effect. Given the substantial variation seen within even the known small samples of African early H. erectus, such an evolutionary history of the Asian populations could result in aspects of morphologies that appear either primitive or distinct. The Dmanisi cranial and mandibular remains are compatible with such an interpretation, which combines primitive cranial morphology with advanced posterior molar reduction and a polymorphic premolar root system (Bräuer and Schultz, 1996; Rosas and Bermúdez de Castro, 1998; Gabunia et al., 2002; Rightmire et al., 2006). Some of the Dmanisi morphologies may stem from genetic drift, and others from an early timing of populational differentiation from the eastern African H. habilis/rudolfensis to H. erectus lineage.
At Konso, H. erectus remains derive from a variety of paleoenvironmental settings, whereas A. boisei occurrences are more confined (Suwa et al., 1997, 2003). Taken together with the abundance of early Acheulean assemblages at ~1.6 Ma and thereafter from varied paleoenvironmental settings at Konso (Beyene, 2003), this may be indicative of a more versatile adaptation of early H. erectus compared to that of A. boisei. The Konso A. boisei remains are associated with a dry grassland fauna (Suwa et al., 2003), and preliminary isotope data suggests an extreme C4 diet for the species (Sponheimer et al., 2003). It is possible that H. erectus ranged more broadly, and A. boisei more restrictively in the eastern African Pleistocene landscape. The morphological evidence presented above suggests that shifts in subsistence patterns at the early Homo to H. erectus transition were associated with increased dentognathic gracility.
Another implication is that population expansion of H. erectus into Eurasia could have occurred at multiple occasions once this broader adaptation was established sometime prior to ~1.6 Ma. Early Acheulean assemblages similar to those found at Konso are known from Ubeidiya in the Middle East, together with limited fauna of African aspects (Tchernov, 1986). Our biochronological assessments based on Kolpochoerus M3 morphology suggest a post-1.4 Ma dispersal event for the Ubeidiya fauna [the Ubeidiya examples are morphologically more advanced than the Konso homologues, see Suwa et al. (2003)]. It is likely that the Levantine record of the early Acheulean was associated with a dispersal event that postdates an earlier one represented by the Dmanisi fossils. Although the evolutionary pattern of Asian H. erectus populations may be clarified only by further morphological and chronological evidence from Asia itself, the Konso evidence suggests that the African populations represent a continuous and successful adaptation that was a likely source of successive events of gene flow to the Eurasian continent.
The Paleoanthropological Inventory of Ethiopia and the Konso Paleoanthropological Research Project were undertaken under the administrative authority of the Center for Research and Conservation of Cultural Heritage (CRCCH), Ministry of Culture and Sports Affairs, Ethiopia, and the Authority for Research and Conservation of Cultural Heritage (ARCCH), Ministry of Youth, Sports and Culture, Ethiopia. We thank the CRCCH, ARCCH and the National Museum of Ethiopia for permission and support for our research projects. We thank the Bureau of Culture, Information and Tourism of the Southern Nations, Nationalities, and People’s Regional Government of Ethiopia, the Konso Special Administrative District, and especially the Konso people for their kind support for the projects. We thank all participants to the fieldwork, especially Awoke Amzaye, who were essential to the success of the research. The Japan Ministry of Education, Science, Sports and Culture, the Japan Society for the Promotion of Science, the Mitsubishi Foundation, and the National Geographic Society provided most of the financial support for the field research at Konso. Further support for laboratory research has been provided by the Japan Society for the Promotion of Science, the National Science Foundation, the Miami University, and the Earth and Environmental Sciences Division Electron Microprobe laboratory at LANL.
|