| Erksin Savas Güleç, corresponding author. e-mail: gulec@humanity.ankara.edu.tr phone: +90-312-309-3761; fax: +90-312-309-3761 Published online 24 August 2007 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.070501 |
The Çorakyerler locality is located at the northeast (NE) corner of the Çankiri basin in central Anatolia (Figure 1). Fossiliferous sediments were discovered here in the early 1970s during a joint German-Turkish mineral exploration program (Sickenberg et al., 1975). The age of the exposed deposits, based on mammalian biochronological correlations, was suggested to be late Miocene (Kaymakçi et al., 2001). Ten years ago our field team initiated new research at Çorakyerler, resulting in the discovery of a hitherto unknown species of hominoid (Sevim et al., 2001; Sevim and Kiper, 2002; Begun et al., 2003), younger than any known previously in western Asia. Biochronological evaluations (see below) suggest correlation with MN11 (Ünay et al., 2006), which is currently placed between 8.7 and 7.4 Ma (Kostopolous et al., 2003).
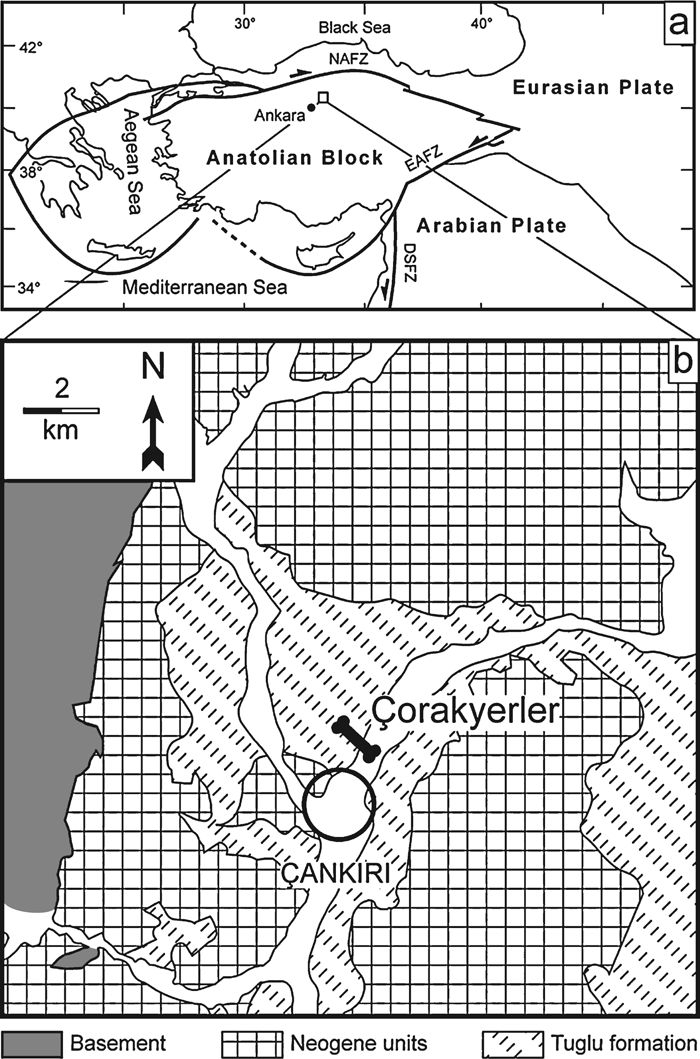 View Details | Figure 1. Location of the Çorakyerler locality. (a) Present-day configuration of Anatolia relative to the Eurasia, Arabian plates, and the Black and Mediterranean seas. Major faults are illustrated. NAFZ, Northern Anatolian Fault Zone; EAFZ, Eastern Anatolian Fault Zone; DSFZ, Dead Sea Fault Zone (a northern extension of the East African Rift System. (b) Geological map of the northeastern corner of the Çankiri-Çorum basin showing the location of Çorakyerler, north of Çankiri. The fossil horizons are included in the Tuglu Formation. (Modified from Kaymakçi et al., 2001). |
The fossil-bearing unit has a rich mammalian fauna and consists of pale-green mottled marl that forms the lower part of a thick succession of continental red beds and overlying evaporites. The Çorakyerler fossils derive from deposits of a large complex of mud flats, probably representing the distal portion of a large alluvial fan-floodplain interfingering with shallow, ephemeral lakes in the northwest (NW) margin of the Çankiri Basin. The overlying evaporites suggest enhanced aridity and an inland sabhka-type environment (Kaymakçi et al., 2001).
The Çorakyerler vertebrate assemblage (Table 1) is dominated by rhinocerotids, hipparionines and bovids, and rarer cervids, suids, gomphotherids, carnivores, and the hominoid primate. The rodent assemblage suggests a savannah-like environment and an MN11 age (Ünay et al., 2006). The greatest faunal affinities are with Kemiklitepe in western Anatolia, Middle Marageh, Iran, and Pikermi, Greece (Bernor, 1986; de Bonis et al., 1994; Bernor et al., 1996; de Bonis and Koufos, 1999). The predominance of spiral-horned antelopes, hipparionines and grazing rhinos (Chilotherium) indicate open woodland conditions, although suids and cervids may indicate slightly more closed woodland than at Kemiklitepe (de Bonis et al., 1994). In total, the Çorakyerler vertebrate fauna suggests a younger age and a more open or at least drier/cooler/more seasonal habitat than hitherto known for any other European or west Asian hominoid locality or taxon (de Bonis and Koufos, 1999; Solounias et al., 1999).
The adult hominoid maxilla (CO-205), with LI1–M3 and RC–M2, is of a large individual, presumed to be male (Figure 2a, b, c). It preserves portions of the right palatine process to the midline, a small fragment of the left palatine process, most of the left alveolar process, and much of the palatal surface of the right premaxilla. There is minor distortion along both tooth rows. The canine is fully erupted and in an early stage of wear, as is the M3.
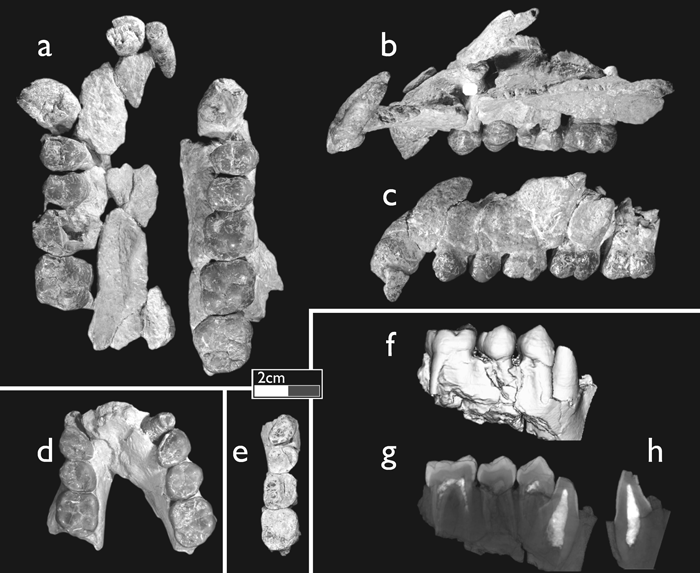 View Details | Figure 2. The Çorakyerler hominoid fossils. CO-205: (a) occlusal view; (b) lingual view of right side and left I1 and I2; (c) buccal view of left C–M3. CO-300: (d) occlusal view; (f) micro-CT-based imagery of right dental row, buccal view; (g) summed voxel projection of right dental row; (h) summed voxel projection of lower canine perpendicular to previous view. CO-710: (e) occlusal view. A summed voxel projection adds CT values of all projected voxels and corresponds to conventional X-ray images. |
A partial subadult mandible (CO-300) is distorted and lacks its base, but retains the upper symphysis, erupting right canine, left and right P3 to M1, and right M2 (the latter tooth was found isolated 50 cm from the mandible) (Figure 2d, f, g, h). The size and morphology of the P3, and a computerized tomography (CT) image of the unerupted canine, suggest that this individual is also male.
A small partial right mandible lacking the base, CO-710, preserves P3 to M2 (Figure 2e). The dentition is more worn than the other specimens, and based on size is probably from a female. It is very small compared to CO-300, and has a smaller dentition than RPL-54a, a probable female mandible of O. macedoniensis (de Bonis et al., 1974; de Bonis and Melentis, 1977).
The uniquely derived dental and palatal features of the Çorakyerler primate specimens warrant recognition as a new hominoid species.
Order Primates Linnaeus 1758
Suborder Anthropoidea Mivart 1864
Superfamily Hominoidea Gray 1825
Genus Ouranopithecus Bonis and Melentis 1977
Species Ouranopithecus turkae sp. nov.
Etymology. Turkae for the people who live in Anatolia.
Holotype. CO-205, a maxilla fragment with right C–M2 and left I1–M3.
Paratypes. CO-300, a subadult mandible with right C–M2 and left P3–M1; CO-710, an adult partial right mandible with P3–M2; holotype and paratypes are housed at the University of Ankara.
Diagnosis. O. turkae is distinguished from other Miocene hominoids, including Ankarapithecus as well as the probable sister taxon Ouranopithecus macedoniensis, by a constellation of dentognathic features that includes shorter and more vertical premaxilla, palate narrow relative to postcanine occlusal size, and homomorphic P3 and P4. The P3 is nearly symmetrical and oval in occlusal outline and the pre-and postparacrista are subequal in length. O. turkae is further distinguished from O. macedoniensis by its smaller relative canine to cheek tooth size proportions, shorter-crowned male canines, maxillary incisors nearly aligned with the canines, and perhaps larger (male) size.
The upper incisors are very heteromorphic in size and shape. The I1 is robust and squat, with nearly equivalent mesiodistal and labiolingual dimensions (Table 2). Its lingual surface is marked by deep vertical fissures, including prominent mesial and distal lingual grooves. The worn incisal edge angles steeply lingually. A prominent basal tubercle, contributing to the labiolingually thick crown, is slightly worn along the incisal wear plane. The I2 crown is wider labiolingually than mesiodistally, and has a strong basal lingual cingulum and marked lingual relief. Relative to the cheek teeth, the incisors are small.
In buccal aspect, the upper canine crown shoulders are set near the crown base. The basal canine outline is a labiolingually elongate oval. The mesial groove is narrow and deep. The right canine tip was fractured antemortem and the dentine was slightly worn apically. There is a distolingual honing facet formed by contact with the P3 and an extensive and mammelated lingual cingulum. Relative to postcanine tooth size, CO-205 has among the smallest canines of presumed males of any known late Neogene hominoid (Figure 3).
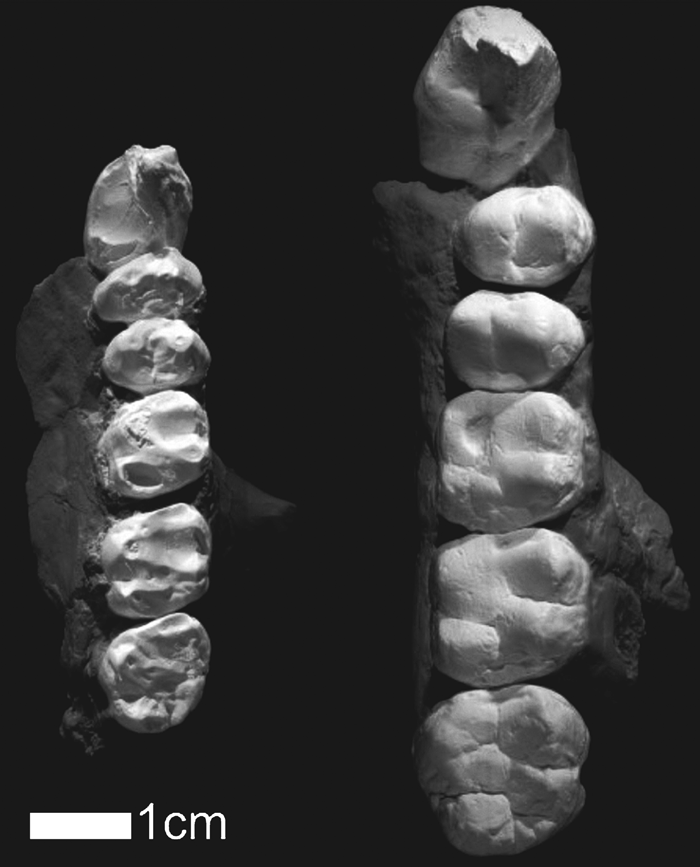 View Details | Figure 3. The Valles-Penedes Dryopithecus laietanus IPMC 18000-5 (left) and Corakyerler CO-205 (right). IPMC 18000-5 is mirrored to facilitate comparisons (from Moyà Solà and Köhler, 1995). |
The O. turkae upper postcanine teeth are in the size range of living gorillas. However, they feature broad, low, rounded cusps, weak cristae, broad, shallow occlusal basins, and simple occlusal morphology. The summed postcanine occlusal area of the CO-205 exceeds that of all other Miocene hominoid primate taxa except Gigantopithecus.
Except for P3, O. turkae molars and premolars lack cingula and their enamel is thick. The enamel thickness of the lateral crown faces in CO-300 measured by recently standardized micro-CT methodology (Suwa and Kono, 2005) at the major cusps ranges from 1.81 to 2.35 mm in the M2 (Figure 4) and from 1.55+ to 2.03+ mm in the M1. Average enamel thickness in the mesial cusp section of the little-worn M2 is 1.94 mm. The relative enamel thickness value (enamel area divided by square root of dentine area; Smith et al., 2003, 2004; Kono, 2004) of M2 is 27.3, making this the thickest enameled Miocene hominoid molar yet examined based on these parameters (Smith et al., 2003, 2004).
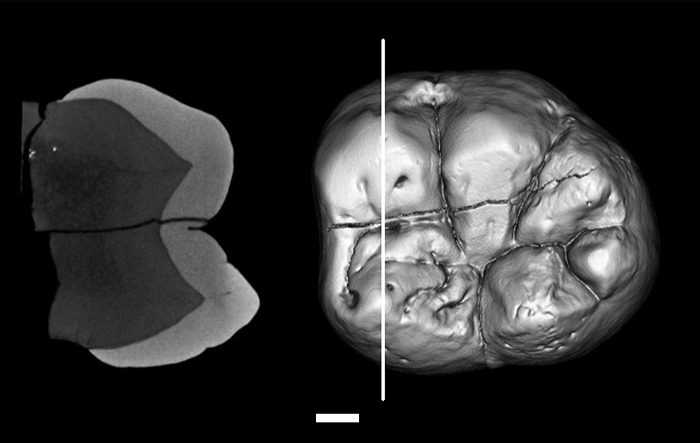 View Details | Figure 4. Radial enamel thickness of the lateral crown face at each of the five major cusps measured by microfocal X-ray CT. Right, occlusal view surface rendered image of the M2 (CO-300) with white line indicating position of the mesial cusp section (section running through the protoconid and metaconid dentine cusp tips); left, corresponding grayscale image of the mesial cusp section; white bar indicates 2 mm. Maximum lateral enamel thickness in the little-worn lower M2 mesial cusp section (Suwa and Kono, 2005) was 2.35 mm in the protoconid and 2.08 mm in the metaconid. The three-dimensional radial lateral enamel thickness ranged from 2.26 to 2.31 opposite the buccal M2 main cusps and 1.81 to 2.15 opposite the lingual M2 main cusps. In the right M1 with flattened buccal cusps, the maximum lateral enamel thickness in the mesial cusp section was 1.55+ in the protoconid and 1.73 in the metaconid. The three-dimensional radial lateral enamel thickness ranged from 1.63+ to 2.03+ opposite the buccal M1 main cusps, and 1.68 to 1.73 opposite the lingual M1 main cusps. Scans were taken at 150 micron pixel resolution for the mandibular canine through M1, and at 40 micron pixel resolution for the mandibular M2 |
The O. turkae P3 is oval, rather than of an asymmetrical triangular form as seen in most other fossil and modern apes. The P3 protocone and paracone are of subequal size. In buccal aspect, the crown profile also lacks the asymmetry seen in other ape P3s. This is because of the absence of a strong rootward mesiobuccal projection of the enamel line, and because the distobuccal (postparacrista) and mesiobuccal (preparacrista) occlusal edges are nearly equal in length. The P4 crown is also more oval and symmetric than in most other apes. The mesial and distal foveae of both upper premolars are bounded by thick marginal ridges.
In keeping with the large and robust premolars, the upper molars are massive, with distal cusps well expressed and not reduced as in some Miocene hominoids. The M1 is smaller than the M2, which is slightly smaller than M3. The latter tapers distally, is mesiodistally elongate, and bears accessory distal cusps. The M1 cusps were worn low before enamel perforation, with dentine exposed only at the protocone. The enamel thickness of the upper molars was not measured, but the enamel bounding the exposed dentine suggests a thickness comparable to that of the measured lower molars.
Only the roots and alveoli of the mandibular incisor are preserved in CO-300. The erupting canine had not reached occlusion, but it is tall, narrow, and pointed. In keeping with the morphology of the upper premolars, the P3 is more oval in occlusal outline than in most apes and even some early hominids. A thin lingual cingulum is present. The protoconid is sharp and prominent. A small honing facet can be seen on the mesio-buccal face of the right P3. The P3 of CO-710 has a prominent wear facet from protoconid to metaconid and also has a slight honing facet on the mesiobuccal surface. As is the case for the P3, O. turkae has slightly more oval P3s than those of O. macedoniensis.
As in the P3, in buccal view, the mesial occlusal edge (preprotocristid) of the P3 in CO-300 is subequal (but slightly longer) in length compared to the distal occlusal edge (postprotocristid). This morphology is unlike the distinctly longer preprotocristid of most other Miocene and modern apes. Hence, the O. turkae P3 is rather less sectorial in shape than in most other known Miocene hominoids, resembling the modern chimpanzee in this respect. The mandibular dental dimensions are given in Table 3.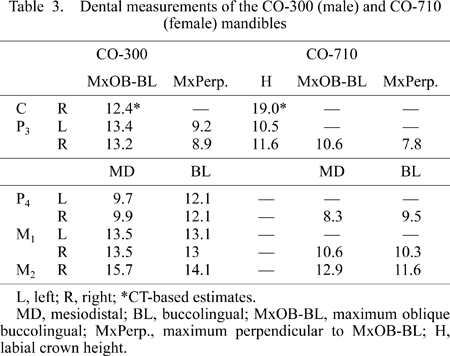
The palate of CO-205 is distorted, but judging from the posterior margins of the incisor and canine alveoli, as well as a fragment of bone along the right lateral edge of the nasal aperture, it featured a short and fairly vertical premaxilla. As the incisors and canines are relatively reduced, the anterior palate is presumed to be relatively narrow, whereas the massive postcanine teeth render the entire palate long and seemingly narrow relative to postcanine size and palatal length. The incisors extend minimally beyond the anterior transverse plane of the canines and are notably vertically implanted, particularly for a young adult. There was little or no precanine diastema. The maxillary palatine processes are thin. We consider O. turkae to have had a more vertical face than O. macedoniensis. These apparently derived features may relate to the younger age of the Turkish fossils.
O. turkae is most similar to Ouranopithecus among known Miocene hominoids. The two taxa share many derived features, such as the weakly asymmetric upper and lower P3s, absolutely and relatively large postcanine size, and hyper-thick molar enamel, probably related to a diet that required heavy mastication. This is also evidenced by instances of heavy wear in upper (RPL-128, XIR-1) and lower (RPL-56) dentitions of this genus. This wear was probably associated with an adaptation to more open habitats than those of extant apes. Indeed, Ouranopithecus lived in Eurasia during a time of considerable decline in forest cover, and development of more open country and seasonable environments (Bernor, 1983; de Bonis et al., 1994; Bernor et al., 1996; de Bonis and Koufos, 1999; Begun and Kordos, 1997; Begun, 2005).
Previous interpretations of Ouranopithecus had suggested that this genus was a plausible ancestor of African Pliocene Australopithecus based on dentognathic morphological similarities (Begun et al., 2003). Indeed, the resemblance of the upper premolars of O. turkae and non-robust Pliocene Australopithecus such as Au. africanus, is striking. However, most early Australopithecus dentitions, A. anamensis (Ward et al., 2001) and A. afarensis (White, 1977), actually have more primitive postcanine dental features than seen in Ouranopithecus. Furthermore, the African late Miocene hominids Orrorin (Senut et al., 2001), Ardipithecus (White et al., 1994; Haile-Selassie, 2001; Haile-Selassie et al., 2004), and Sahelanthropus (Brunet et al., 2002) lack dentognathic specializations associated with tough/abrasive diets such as that inferred for Ouranopithecus and in early Australopithecus. Such considerations lead us to interpret Ouranopithecus as manifesting substantial dentognathic parallelism with Australopithecus (Begun and Kordos, 1997; Begun, 2001). Hence, we do not consider these features of Ouranopithecus to indicate placement within the hominid (African ape-human) clade (contra de Bonis et al., 1990, 1999; de Bonis and Koufos, 1993, 1994, 1997; Koufos, 1993).
O. turkae, the youngest and largest of known Turkish Miocene hominoids, is unlike all modern and other fossil great apes in what is known of its preserved anatomy and its environmental circumstances.
We thank the Republic of Turkey and the Turkish Ministry of Culture and Tourism, The Governor of Çankiri and National Science Foundation (including the Revealing Hominid Origins Initiative/HOMINID program, Grant No. 0321893) for financial support of field and laboratory research. We thank all the members of the Çorakyerler research expedition and particularly to Dr. David Begun who prepared the maxilla and provided useful observations for this specimen. Our work benefited from the comments and assistance of Ray Bernor, Faysal Bibi, Hans de Bruijn, Denis Geraads, F.C. Howell, Mustafa Karabiyikoglu, Nuretdin Kaymakçi, Pinar E. Kaymakçi, Jay Kelley, Gen Suwa, Engin Ünay, and Tim White. Enamel thickness was investigated by micro-CT in collaboration with G. Suwa and R.T. Kono.
|