| * Correspondence to: Hajime Ishida, Department of Anatomy, Faculty of Medicine, University of the Ryukyus, 207 Uehara, Nishihara, Okinawa 203-0215, Japan. E-mail: ishidaha@med.u-ryukyu.ac.jp Published online 14 November 2007 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.070323 |
In northeastern Asia, human skeletal remains from the Late Pleistocene have been recovered at the Afontova Gora II and Mal’ta sites (Alekseev and Gokhman, 1983). Many Neolithic human skeletal remains have been recovered in the Lake Baikal region (Debets, 1951; Ishida, 1995). Recently, some skeletons from the Neolithic or Bronze Age (6000–3000 BP) were excavated in the Amur Basin (Balueva, 1978), Yakutia (Gokhman and Tomtosova, 1983), southern Primorye (Chikisheva, 1997), and the Arctic region (Gokhman, 1961). The results of previous work indicate that people in Northeast Asia were beginning to show distinctive local variation in cranial morphology by the Neolithic period.
The people inhabiting a region that includes eastern Siberia and eastern Asia in the Late Pleistocene have been identified as the antecedents of people who migrated into not only the Americas but also into the Japanese archipelago. There were three main cultural waves from Northeast Asia to the northern Japanese Islands, including Hokkaido. The first and second ones were Paleolithic microblade industries (15000–12000 BP) and Mesolithic blade arrowhead cultures (10000 BP), although no human remains associated with this cultural tradition have been recovered (Kikuchi, 1999; Yamaura and Ushiro, 1999). Another, much later, migration of people and culture, the Okhotsk culture, from northeastern Asia has been identified (Kikuchi, 1995; Amano, 2003).
The Okhotsk culture spread from southern Sakhalin Island to northeastern Hokkaido Island and the Kurile Islands during the 5th to 12th centuries AD (Amano, 2003). The Okhotsk culture developed a considerable maritime infrastructure, one which was different from that of the native population in Hokkaido (Hudson, 2004). The subsistence and trade patterns of the Okhotsk culture have been recently reported using ancient DNA and stable isotope analysis (Masuda et al., 2001, 2006; Yoneda, 2002). For example, ancient DNA analysis on brown bear crania from the Okhotsk culture revealed that brown bears born in southern Hokkaido were found with people associated with the Okhotsk culture (Masuda et al., 2001).
Modern investigation into the Okhotsk culture began when the Susuya shell mound in Sakhalin Island was excavated by several researchers from the beginning of 20th century (Kiyono, 1925). After World War II, many human skeletal remains of the Okhotsk culture were recovered from the Moyoro shell mound, Abashiri, in eastern Hokkaido Island (Kodama, 1948; Ito, 1965). The skulls associated with the Okhotsk culture possess features typical of the northeastern Asian cranial series. These features generally include a large, high face, flat nasal bones, and large maxillary and zygomatic bones (Kodama, 1948; Yamaguchi, 1974; Ishida, 1988). Okhotsk skulls have a high frequency of the supraorbital foramen, hypoglossal canal bridging, transverse zygomatic-suture vestige, and mylohyoid bridging (Kozintsev, 1990, 1992; Ishida, 1996).
Previously, one of the authors (H.I.) excavated the remains of approximately 50 individuals associated with the Okhotsk culture, work which included the analysis of nonmetric cranial variation in these remains (Ishida, 1988, 1994; Ishida et al., 1994). The markedly high, broad, and flat face of the Okhotsk contrasted sharply with the low and prominent face of the Ainu living in Hokkaido, Sakhalin, and the Kurile Islands, places where the Okhotsk people are presumed to have once lived. On the other hand, analysis of the nonmetric cranial variation suggested that the Ainu were initially closer to the Okhotsk than to other Northeast Asian series (Ishida, 1996). Further analysis of cranial metric and nonmetric variation, including the use of the method of Relethford and Blangero (1990) by Shigematsu et al. (2004) and Kondo (2005), suggested that the admixture between the ancestors of Ainu (Epi-Jomon or later) and the Okhotsk might account for regional differences among the recent Ainu crania.
Moreover, it was revealed that the Sakhalin Okhotsk crania were more similar to the Omisaki and Hamanaka Okhotsk from northern Hokkaido than to the Moyoro Okhotsk based on the analysis of cranial measurements (Ishida, 1994). Regional differences in the morphology observed in Okhotsk crania correspond to cultural differences identified by Amano (2003), suggesting that studies of the regional differences and diversity of the Okhotsk and Ainu series might help elucidate their origins, dispersal, and interactions.
In 2003, approximately 260 Okhotsk skeletal remains formerly in the School of Medicine, Hokkaido University were transferred to the Hokkaido University Museum, a transfer that provided an opportunity for the authors to re-examine these remains in detail. The size of this skeletal series allowed comparisons of regional variation in not only the Okhotsk but other prehistoric northeastern Asian cranial series. In this study, we report on the nonmetric cranial variation in northern and eastern Okhotsk crania to understand the population history of the Okhotsk, Ainu, and other inhabitants of Northeast Asia.
Because of differences in Okhotsk ceramics, metal ornaments, and burial patterns between the two regions (Ono and Amano, 2007), the cranial material representing the Okhotsk culture is divided into two regional groups, the northern Okhotsk and the eastern Okhotsk (Table 1, Figure 1). The northern Okhotsk series consists of the Susuya shell mound (Kiyono, 1925; Ishida, 1994) and the Minami Kaizuka shell mound (Ishida, 1994) on Sakhalin Island; Funadomari, Hamanaka (Ito and Kodama, 1963; Ishida et al., 1994; Ishida and Matsumura, 2000; Ishida et al., 2002), and Oshonnai (Ishida, 1991) on Rebun Island; the Omisaki shell mound (Mitsuhashi and Yamaguchi, 1961, 1962a, b, c; Ishida, 1988; Yamaguchi, 1995) and Pirikatai shell mound (Yamaguchi, 1991) in Wakkanai on Hokkaido Island. The eastern Okhotsk specimens are from the Moyoro shell mound (Kodama, 1947, 1948; Ito, 1965) and Omagari (Ishida and Yonemura, 1993) in Abashiri, Utoro (Ishida et al., 1993; Ishida et al., 1994) in Shari on Hokkaido Island, and Shikotan in the Kurile Islands.
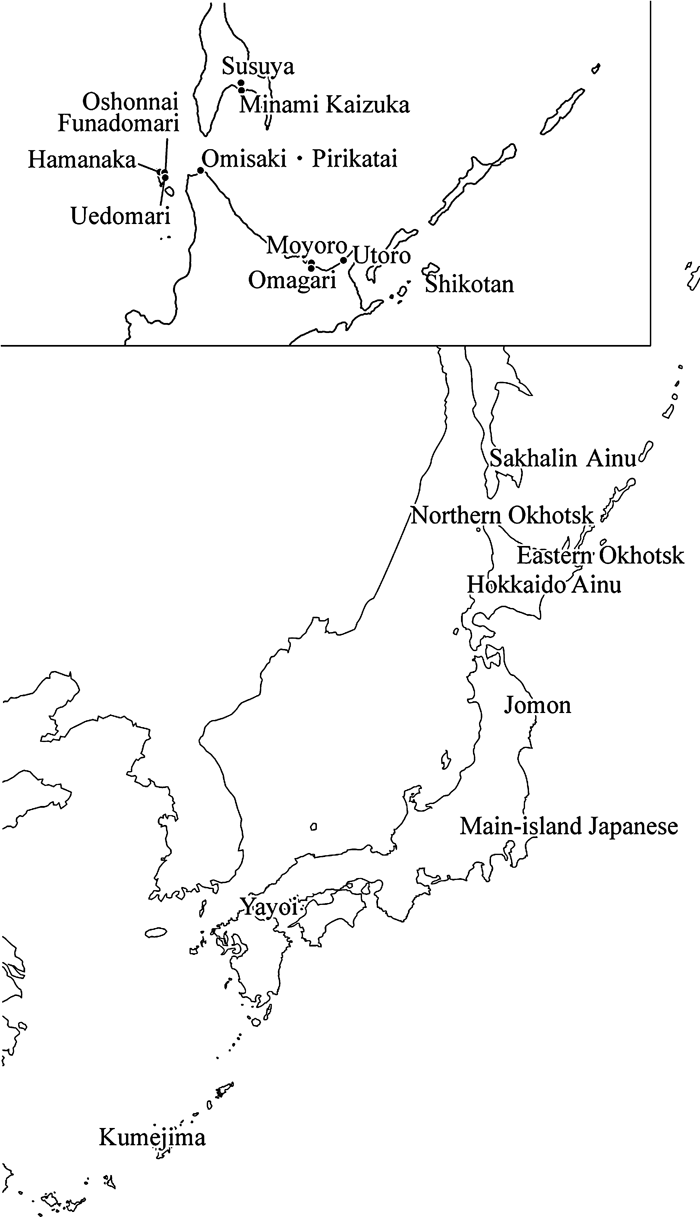 View Details | Figure 1. Location of cranial series from the Japanese islands used in this study (bottom) and a map of the Okhotsk cultural sites listed in Table 1 (top). |

As mentioned by Ito (1965), the skeletal remains from the Moyoro site possibly contained the remains of not only the Okhotsk people but also several Ainu skeletons from medieval and early modern times. To determine which of the skeletons from this site were those of the Okhotsk culture, the one of us (M.Y.) undertook the radiocarbon dating of the Moyoro series using NIES-TERRA, the accelerator facility of the Institute of Environmental Studies located in Tsukuba City, Japan (Yoneda et al., 2005; Matsumura et al., 2006). For example, since the calibrated age of 1265–1365 AD (Moyoro skull No. 1060) corresponded to medieval times, this skull was eliminated from our study. Further results of radiocarbon dating and the results of stable isotope analysis of the Moyoro and other Okhotsk samples will be reported in the near future.
The comparative samples used in this study consist of the following 19 groups: Jomon, Hokkaido Ainu, Sakhalin Ainu, Yayoi, modern main-island Japanese, Kumejima, Northern Chinese, Aleut, Neolithic Baikalian, Buryat, Chukchi, Ekven (Iron Age), Asian Eskimo, Kazakh, Mongolian, Amur (Nanay, Negidal, Oroch, and Ulch), Nivkh, Russian, and Tagar (Iron Age, southern Siberia). The Jomon from the middle to latest periods (c. 5300–2300 BP) were mainly from the main Japanese islands of Hokkaido and Honshu. The Jomon collections used in this study are housed in the National Museum of Nature and Science, Tokyo, University Museum of the University of Tokyo, Tohoku University School of Medicine, and Sapporo Medical University. The Yayoi samples were excavated from the northern Kyushu and Yamaguchi regions, and are believed to be immigrants from continental East Asia or their descendants. The archeological, geographical, and ethnographic contexts of the other comparative samples are provided in Table 2 and Figure 2, and in our previous papers (Ishida, 1992, 1995, 1996, 1997; Ishida and Dodo, 1992, 1996; Fukumine et al., 2006).
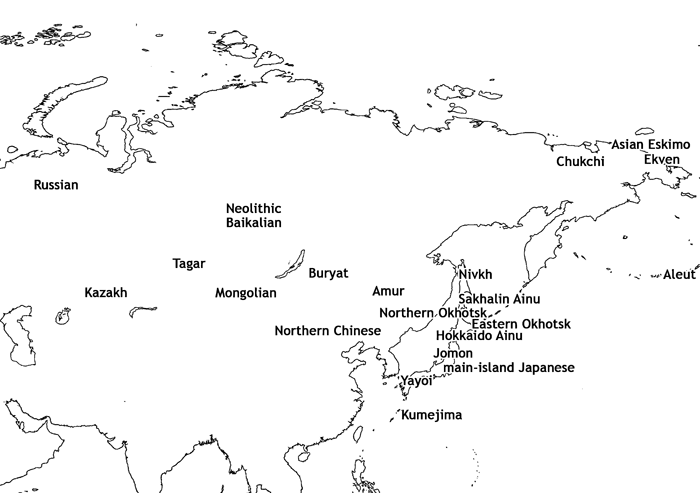 View Details | Figure 2. Location of the cranial series used in this study. |
In an attempt to eliminate interobserver error, H.I. scored all 19 nonmetric cranial traits (as present or absent) for northern and eastern Okhotsk, and all comparative samples of both sexes in this study, following the criteria of Dodo (1974, 1986a) and others (Hanihara and Ishida, 2001a, b, c, d, e). Three other traits normally recorded, the biasterionic suture vestige, tympanic dehiscence, and foramen of Vesalius, were eliminated from this study because of high intra- and interobserver errors (Ishida and Dodo, 1990; unpublished data). As is common in nonmetric studies (Hanihara et al., 2003), the frequencies of the 19 nonmetric cranial traits were calculated based on pooled-sex individual counts. That is, a trait was scored as present if it was observed on one or both sides (Dodo, 1974).
Fisher’s exact probability test was used to evaluate the frequency differences between the northern Okhotsk and eastern Okhotsk, Hokkaido Ainu, Sakhalin Ainu, and Jomon, respectively. The modified Smith’s mean measure of divergence (MMD) statistic was computed between the pairs of samples used (Sjøvold, 1973), excluding the Nivkh due to small sample size. MMD has been commonly used in the analysis of nonmetric cranial traits and its efficacy has been summarized in a previous paper (Hanihara et al., 2003).
The multidimensional scaling method (Sneath and Sokal, 1973) was applied to the MMD distance matrix to represent mutual relationships. The neighborjoining method (Saitou and Nei, 1987), commonly used in phylogenetic analyses, was also used to sort the MMD distance matrix. As constructing an unrooted tree to hypothesize a constant rate of evolutionary change is not necessary for the neighbor-joining method, we applied the neighborjoining method to the phenotypic distance matrix rather than the unweighted pair group method with arithmetic mean (Dodo et al., 1992).
In addition to MMDs, we calculated the standard relationship matrix (R-matrix) to analyze the relationship within and among populations (Harpending and Ward, 1982; Relethford and Blangero, 1990; Relethford and Harpending, 1994).
The R-matrix method was originally developed for the frequency of genetic traits by Harpending and Ward (1982). Because Relethford and Blangero (1990) developed a model for the R-matrix method to analyze quantitative morphological traits, many researchers now use this method to elucidate probable human history and to compare variances within and among geographical groups (Relethford, 1991, 1994; Relethford and Harpending, 1994; Powell and Neves, 1999; Steadman, 2001; González-José et al., 2003; Roseman and Weaver, 2004; Stojanowski, 2004, 2005; Hanihara and Ishida, 2005; Schillaci and Stojanowski, 2005). Thus, the effects of differential external gene flow can be assessed by comparing observed and expected variances (Relethford and Balngero, 1990).
On the other hand, Hallgrímsson et al. (2004) used nonmetric morphological trait frequencies as analogous to allele frequencies in computing the R-matrix and other genetic parameters. In a previous study, we calculated the R-matrix and related parameters under the assumption that nonmetric dental trait frequencies behave like allele frequencies because of the very high correlation between the R-matrix and other distance measures, such as MMDs (Haneji et al., 2007); however, the matrix correlation is not necessarily a theoretical justification, so the results of this analysis should be interpreted cautiously.
As phenotypic traits are not completely under genetic control, the actual values of parameters based on the R-matrix method, such as inter- and intraregional variation, should be assessed using estimates of the average heritability of phenotypic traits. Only a few previous studies have reported the heritability estimates of nonmetric cranial traits based on human family studies, or human or other primate skeletons (Saunders and Popovich, 1978; Cheverud and Buikstra, 1981a, b; Sjøvold, 1984; Mouri, 1997; Velemínský and Dobisíková, 2005). Cheverud and Buikstra (1981a, b) reported the heritability of nonmetric cranial traits in rhesus macaques. The mean heritability rate of the nonmetric cranial traits used in their study was c. 0.64 (ranging from 0.48 to 0.77). Sjøvold (1984), using the human skull collection from Hallstatt village in Austria, estimated the heritability of nonmetric cranial traits to be approximately 0.30 (from 0.10 to 0.56). Other studies also showed that the heritability rate of nonmetric cranial traits is lower than that of craniometric traits (Saunders, 1989).
However, Dodo and Ishida (1990, 1992) have demonstrated the stability of nonmetric cranial traits used against environmental stress on the basis of the trait frequencies of prehistoric and historic Japanese crania. Hanihara and Ishida (2001a, b, c, d, e) have successively investigated nonmetric cranial traits worldwide. Furthermore, the ability of nonmetric cranial analysis to differentiate cranial series worldwide was reconfirmed by Hanihara et al. (2003).
Taking into account the findings of this previous research, we tentatively selected estimates of average heritability for nonmetric cranial traits of 0.4, 0.5, and 0.6, respectively. Since the heritability of metric cranial and dental traits was estimated as 0.55 (Relethford, 1994; Hanihara and Ishida, 2005; Toma et al., 2007), the results obtained in this study can be directly compared with previous work applying the R-matrix method.
Table 3 shows the frequencies of 19 nonmetric cranial traits in the Okhotsk, Ainu, and Jomon samples. The frequency of the supraorbital foramen in the Okhotsk series is higher than in the Hokkaido Ainu and Jomon, while the incidence of the transverse zygomatic suture vestige in the Okhotsk series is as high as in the Jomon and higher than in the Sakhalin Ainu (P < 0.05). The Okhotsk, especially the eastern Okhotsk (P < 0.05), and Ainu generally have higher frequencies of hypoglossal canal bridging than the Jomon.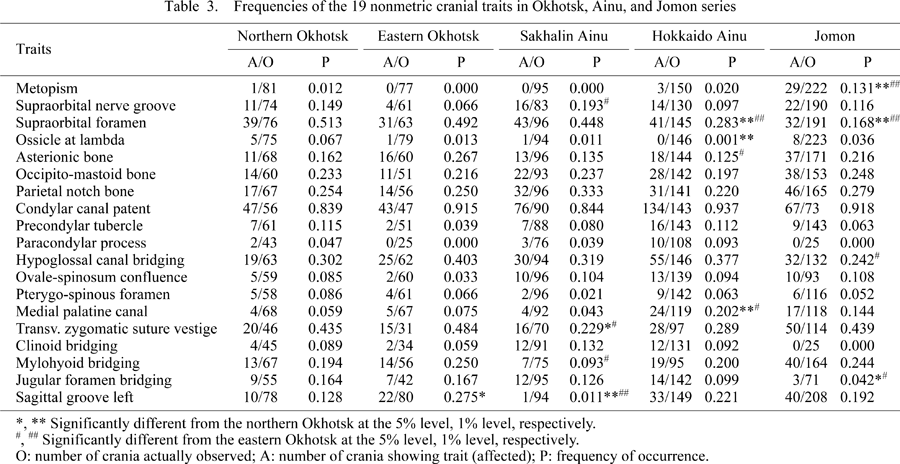
Fisher’s exact probability test was performed between the northern and eastern Okhotsk, and the Ainu and Jomon samples, respectively. Between the northern and eastern Okhotsk, only one trait was significantly different. Two traits in the Sakhalin Ainu and three traits in the Jomon and Hokkaido Ainu were found to be significantly different from the northern Okhotsk, while the eastern Okhotsk had more significant traits.
Figure 3 shows a graph of the frequency patterns of the five main traits, which are the most effective in discriminating the East Asian series among the 19 traits (Dodo and Ishida, 1990), in the Amur, Nivkh, northern Okhotsk, eastern Okhotsk, Sakhalin Ainu, Hokkaido Ainu, and Jomon series. The frequency of the supraorbital foramen gradually decreased from the Amur to the Jomon. The frequency of the transverse zygomatic suture vestige and mylohyoid bridging roughly increased from the Amur to the Jomon, although the Ainu had a relatively low frequency. Figure 4 shows a graph of the five trait frequencies for a wider geographic comparison. The frequency of the supraorbital foramen also decreased from the Asian Eskimo to the Jomon, whereas that of the medial palatine canal increased. The eastern Okhotsk and Hokkaido Ainu had high frequencies of hypoglossal canal bridging, as did the Asian Eskimo.
 View Details | Figure 3. Frequencies of the five main nonmetric cranial traits (1). |
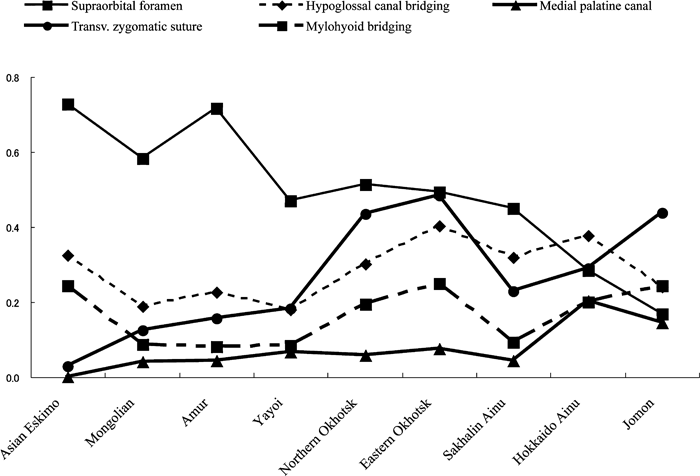 View Details | Figure 4. Frequencies of the five main nonmetric cranial traits (2). |
MMDs were calculated using 19 nonmetric cranial traits for 20 series (the Nivkh series was removed due to small size). The distance matrix and standard deviations are given in Table 4.
The MMD between the northern and eastern Okhotsk is small and insignificant. The northern Okhotsk is closest to the Sakhalin Ainu and Neolithic Baikalian. The groups closest to the eastern Okhotsk are the Hokkaido Ainu and Sakhalin Ainu. Interestingly, the Hokkaido Ainu is closer to the Okhotsk than to the Jomon based on MMDs. The northeastern prehistoric series had generally closer affinities to other northeastern prehistoric series in different areas than to recent samples. For example, the northern and eastern Okhotsk series were nearer to the Neolithic Baikalian than to the recent Amur and Buryat, or to the Iron Age Ekven than to recent Asian Eskimo, respectively.
Next, we applied the multidimensional scaling method to the MMD matrix to represent two-dimensional relationships. Figure 5 and Figure 6 show the graph of the first and second, and the first and third axes, respectively. The relatively isolated Jomon are located in the lower left portion in Figure 5. The Okhotsk and Ainu are relatively close to the Jomon in this diagram. The Arctic and Russian series are situated in the right upper and lower portions, respectively. The prehistoric series, including the Okhotsk, are nearer the Jomon, as well as the Ainu, as mentioned above. In Figure 6, the Okhotsk, especially the eastern Okhotsk, located in the middle left portion of this diagram, are separate from the Ainu.
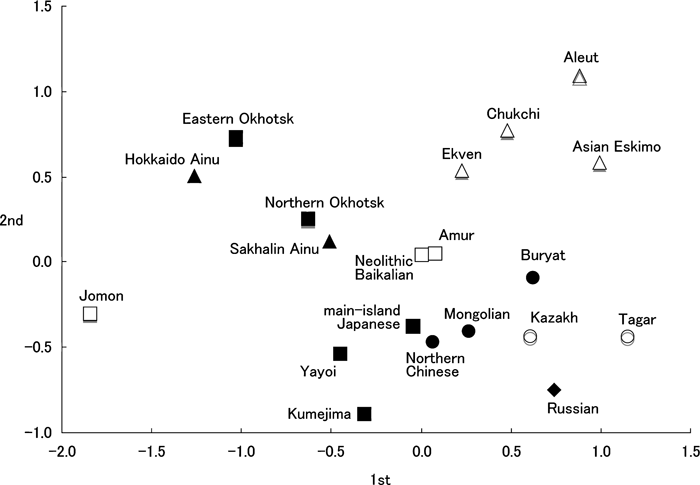 View Details | Figure 5. Multidimentional scaling analysis using first and second axes of the MMD matrix based on 19 nonmetric cranial traits. |
 View Details | Figure 6. Multidimentional scaling analysis using first and third axes of the MMD scores based on 19 nonmetric cranial traits. |
The results of applying the neighbor-joining method (Saitou and Nei, 1987) to the MMD scores are shown in Figure 7. The Jomon and Hokkaido Ainu are the first to connect to form a cluster. Interestingly, this cluster, and the eastern and northern Okhotsk, are grouped together. This branch joined with a cluster comprising the Yayoi and Kumejima series. Two other distinct branches emerged from the center of this diagram; the first consists of the Arctic series, and Neolithic Baikalian and Amur; the second branch culminates in the Russian, Tagar, Kazakh, and East Asian series.
 View Details | Figure 7. A tree of 20 cranial series based on application of the neighbor-joining method applied to the MMD scores based on 19 nonmetric cranial traits. |
Assuming that the samples used in this study have the same effective population size, interregional variation was estimated by Fst values. Table 5 shows the minimum Fst, using an average heritability rate (h2) = 1, and estimated Fst (h2 = 0.4, 0.5, 0.6) for nonmetric cranial traits. The results of estimated Fst, varying from 0.0655 to 0.0951, show that nonmetric cranial variation across regions is fairly limited, which is approximately concordant with the low levels of interregional variation in analyses of genetic, cranial, and dental data (Nei and Roychoudhury, 1982; Relethford, 2002; Hanihara and Ishida, 2005; Haneji et al., 2007).
Table 6 gives the results of Relethford and Blangero’s (1990) method, estimating intra-regional variation under the assumption of an equal effective population size. Large Rii values (distance from the centroid) of the Jomon (0.0973), Aleut (0.0905), Hokkaido Ainu (0.0669), and eastern Okhotsk (0.0653) indicate less than expected variation, or less genetic variability (Harpending and Ward, 1982). The northern Okhotsk (0.0340) has lower Rii than the eastern Okhotsk. As mentioned in the MMD analysis, the prehistoric samples of Ekven and Neolithic Baikalian also show smaller Rii than recent series. Greater observed variation compared with the expected variation is found in the northern Okhotsk and Kumejima. On the other hand, the main-island Japanese, Amur, Neolithic Bailkalian, and Sakhalin Ainu showed less phenotypic variation than expected.
Figure 8 and Figure 9 show the multidimensional scaling of distances transformed from the R-matrix using the average heritability rate (h2) = 0.6 (distance matrix not shown). The arrangement of this series is much the same in Figure 5 and Figure 6, based on MMDs. The eastern Okhotsk was more isolated from the northern Okhotsk and Hokkaido Ainu in Figure 8.
 View Details | Figure 8. A scattergram based on the first and second eigenvectors of the R matrix. |
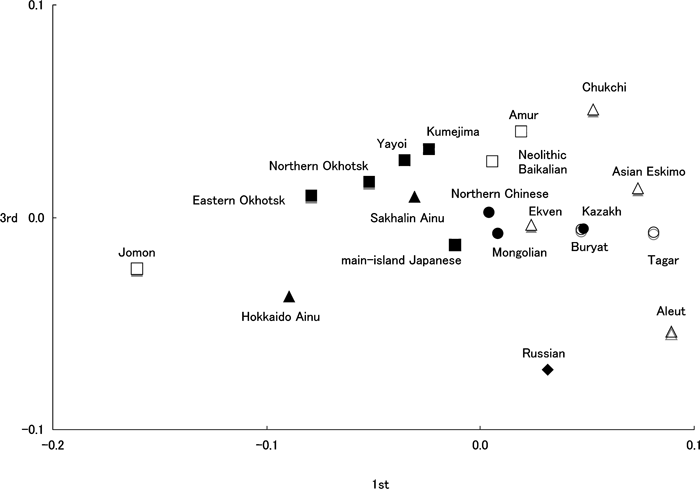 View Details | Figure 9. A scattergram based on the first and third eigenvectors of the R matrix. |
Although the MMD between the northern and eastern Okhotsk series is quite small (0.003), the distance based on the R-matrix is not very small (0.033) compared with that between the northern Okhotsk and Sakhalin Ainu (0.034) in this study. Moreover, the results of Relethford and Blangero’s (1990) method indicate that the eastern Okhotsk had a larger Rii value (distance from the centroid) and lower observed variation than the northern Okhotsk (Table 6). Because the large Rii indicates that the eastern Okhotsk is peripheral (Harpending and Ward, 1982), they should lose phenotypic variability. Few differences between the observed and expected variations in the eastern Okhotsk confirm the low phenotypic variability for this site. On the other hand, the northern Okhotsk shows greater observed variation than the expected variation. The positive residual may result from long-term gene flow from an outside source or larger effective population size (Harpending and Ward, 1982; Relethford, 1994).
The Okhotsk culture was present only in southern Sakhalin and northern Hokkaido in the early phase, before it dispersed into eastern Hokkaido and the Kurile Islands during the middle phase (Amano, 2003; Ono and Amano, 2007). Amano’s observation (Amano, 2003) that the adaptive strategy of the Okhotsk people was to migrate into the eastern area because of its small population size is supported by the low phenotypic variability found in this study for the eastern Okhotsk series. Thus, the R-matrix method may give a more accurate representation of population history than methods such as the MMD statistic.
Except for the Ainu, the northern Okhotsk is more similar to the Neolithic Baikalian and Amur (Table 4, Figure 5, Figure 8). Debets (1951) divided recent Siberians into four major groups: the Arctic group (Asian Eskimo, Chukchi, Aleut), Baikal group (Ulch, Nanay, Negidal, Oroch, Evenki and Yukagir), Central Asian group (Mongolian, Buryat, Yakut and south Siberian peoples), and Uralic group based on craniology and somatology. His classification is supported by other researchers (Alekseev, 1979; Kozintsev, 1988; Ishida, 1995, 1997). The grouping of the Amur (Ulch, Nanay, Negidal, Oroch) and Neolithic Baikalian has been previously noted in a number of cranial studies (Alekseev, 1979; Alekseev and Gokhman, 1983; Ishida and Dodo, 1992; Ishida, 1995, 1997; Hanihara et al., 2003).
This study also finds affinities between the Okhotsk, Amur, and Neolithic Baikalian, as has been previously reported in other work (Yamaguchi, 1974, 1981; Ishida, 1988, 1996; Kozintsev, 1990, 1992). Because the Nivkh series was not used in the present study, we are unable to confirm the findings of Kozintsev (1992) and Kikuchi (1995) who claimed that the Nivkh in northern Sakhalin are closest to Okhotsk based on analyses of morphological and archeological data.
As the northern Okhotsk is closer to the Neolithic Baikalian than to the Amur, northeastern prehistoric samples have generally closer affinities to each other in this study. For example, the Iron Age Ekven are more similar to the northern Okhotsk and Neolithic Baikalian than recent Arctic populations, as previously reported in metric and nonmetric studies by Ishida (1996, 1997). The smaller Rii values of these three series may support this; however, the lower observed variation in the Neolithic Baikalian and Ekven found in the present study, which uses the method of Relethford and Blangero (1990), contrasts with the greater observed variation in the northern Okhotsk. This suggests a complex population history influenced by gene flow and/or genetic drift during prehistoric times in Northeast Asia.
These three prehistoric skeletal series of different periods have typical northeastern Asian, or ‘classic Mongoloid’, morphology (Ishida and Dodo, 1996). As Hanihara (1994) found in his review of the Far Eastern skeletal series in the Late Pleistocene and Holocene based on cranial measurement analysis, this study confirms that the Neolithic Baikalian and Ekven are similar to other series in the same region. Given the presence of only a few Late Pleistocene human fossil specimens and three prehistoric skeletal series that represent the vast northeastern Asian region, it will be difficult to resolve the differentiation and dispersal of people in Northeast Asia. Further research using the larger skeletal series associated with the Okhotsk culture, including ancient DNA analyses (Sato et al., 2007), will contribute to understanding the population history for the region.
The results of this new analysis of nonmetric cranial variation demonstrate a close relationship between the Okhotsk, Ainu, and Jomon. This study also supports the view that the Jomon in Hokkaido were the probable ancestors of the Ainu, a finding that is supported by previous analyses of cranial measurements, cranial nonmetric traits, and dental morphology (e.g. Turner, 1976; Yamaguchi, 1982; Ossenberg, 1986; Ossenberg et al., 2006; Dodo, 1986b; Dodo and Kawakubo, 2002; Ishida, 1990, 1992, 1993; Ishida and Kida, 1991; Ishida and Dodo, 1992, 1997; Ishida and Kondo, 1999; Kozintsev, 1992; Pietrusewsky, 1999, 2000, 2004; Shigematsu et al., 2004; Fukumine et al., 2006; Nagaoka and Hirata, 2006). Further, previous morphological and genetic studies have repeatedly suggested that the Ainu shifted northwards (Cavalli-Sforza et al, 1988; Kozintsev, 1990; Omoto, 1995; Omoto and Saitou, 1997; Saitou, 1995; Ishida, 1996; Ishida and Kondo, 1999; Bannai et al., 1999; Tokunaga et al., 2001; Shigematsu et al., 2004; Tajima et al., 2004; Tanaka et al., 2004; Fukumine et al., 2006; Hammer et al., 2006).
In this study, the Okhotsk, Ainu, and Jomon form a single cluster, a finding that is supported, at least in part, by previous studies (Ishida, 1996; Shigematsu et al., 2004). However, the metric cranial and postcranial traits displayed large differences between the Okhotsk, Ainu, and Jomon (Ishida, 1993, 1996). Several researchers have also hypothesized that the Jomon and Ainu show close affinities with Southeast Asian populations in terms of dental morphology and cranial measurements (Turner, 1987, 1989, 1990; Hanihara, 1992; Matsumura, 1994; Matsumura and Hudson, 2005). Possible relationships between the Jomon or Ainu and prehistoric Americans have also been suggested (Brace et al., 2001; Tokunaga et al., 2001; Jantz and Owsley, 2003). Koganei (1893) believed that the Ainu represented an isolated group unrelated to any other group. The large Rii for the Jomon and Ainu may support their peripheral existence (Harpending and Ward, 1982). The results of the present study that indicate a close relationship between the Okhotsk and Jomon-Ainu series would appear to confound the understanding of Jomon and Ainu origins. However, the unusual circumstances of the Jomon and Ainu among prehistoric and historic groups in Asia may hold a key for resolving the dispersal and adaptive strategy of modern humans in Asia.
We are grateful to H. Baba of the National Museum of Nature and Science, Tokyo, G. Suwa of the University Museum, the University of Tokyo, and H. Matsumura of Sapporo Medical University for their kind permission to examine the Jomon skeletal collections under their care. In addition, we thank Y. Mizoguchi (the associate editor) and two anonymous reviewers for their very helpful comments.
This study was supported in part by a Grant-in-aid for Scientific Research on Priority Areas “Medieval Studies in East Asia” (No. 15068210) from the Ministry of Education, Culture, Sports, Science and Technology, and by a Grant-in-aid for Scientific Research (No. 18370099) from the Japan Society for the Promotion of Science.
|