| Soichiro Kusaka, correspondence to: Soichiro Kusaka, Laboratory of Physical Anthropology, Department of Zoology, Graduate School of Science, Kyoto University, Oiwake-cho, Kitashirakawa, Sakyo-ku, Kyoto 606-8502, Japan. E-mail: kusaka@anthro.zool.kyoto-u.ac.jp Published online 21 February 2008 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.070703 |
The period of Jomon culture in the Japanese Islands lasted from 13000 to 2300 years BP. The Jomon people were hunter-gatherers, relying on marine and/or terrestrial food sources, without agriculture, despite their cultural acquisition of pottery and ground stone tools (Yamanouchi, 1964; Imamura, 1993; Kobayashi et al., 2004). This study is of human remains excavated from the Yoshigo and the Inariyama shell mounds of Aichi Prefecture in the Tokai coastal region of Honshu dating from the later part of the Late Jomon period through to the Final Jomon period (c. 3500–2300 years BP). During this period there were developmental changes in Jomon ritual practices and artifacts (Imamura, 1996), and climatically, as revealed by pollen analysis (Tsukada, 1986), the period was characterized by a cooling trend.
Many archaeological studies have investigated the daily food consumption of the Jomon people who, as hunter-gatherers, were dependent on seasonal food resources (Akazawa, 1986), and a variety of important food items have been suggested (Kaneko, 1994; Komiya, 1994; Hayashi, 2004). However, these studies based on faunal and floral remains are of limited potential for the reconstruction of diet, in that food items actually ingested disappear from the site and food remains are subject to taphonomic bias due to decomposition in the soil. A further limitation is that although food remains at a given site may indicate the consumption of particular food items by site-dwellers as a whole, they give no indication of individual differences of diet within the population.
Paleodietary reconstruction using carbon and nitrogen isotope analysis of human bone collagen is a recent development offering solutions to this problem (Ambrose and DeNiro, 1986; Sponheimer and Lee-Thorp, 1999; Bocherens and Drucker, 2003; Newsome et al., 2004; Bocherens et al., 2006). The turnover rate of bone collagen in vivo is about 10 years (Stenhouse and Baxter, 1979; Price et al., 2002), so that stable isotope ratios obtained from human skeletal remains record the proportion of protein in the diet over the 10 years prior to death. Thus, stable isotope analysis has potential for detecting individual differences in the diet of past human populations. If individuals in the same community ingested protein foods in different proportions, there should be intra-site variations in isotopic values. Furthermore, it may be possible to infer the influence of social positioning on diet by relating stable isotope ratios to such obtainable information as sex and social status, inferred from accompanying artifacts (e.g., ornaments) or from ritual dental modifications.
Numerous studies have employed stable carbon and nitrogen isotope analysis of human bones from the Jomon period (Chisholm et al., 1988; Minagawa and Akazawa, 1992; Minagawa, 2001; Yoneda et al., 1996, 2002, 2004). Minagawa (2001), for example, investigated differences in diet between a number of Jomon populations. He reported that the coastal Jomon people in Hokkaido, western Honshu and Kyushu were largely dependent on marine food sources, while inland Jomon people tended to consume food from terrestrial sources. Yoneda et al. (2004), however, emphasized the importance of salmon and trout for inland Jomon people, based on the isotope ratios of these populations.
While these studies focused on diet at the local population level, only a few studies have looked at intra-site individual differences. The possibility of sex-based dietary differences in Jomon people in Hokkaido (Chisholm et al., 1992), and in Nagano in central Japan (Yoneda et al., 1996), has been suggested. Chisholm et al. (1992) suggested that the sex-related differences in isotopic values they observed in Hokkaido resulted from different modes of food acquisition in males and females: males hunted, while females fished and gathered plant foods. The success of these studies suggests the possibility of inferring aspects of social organization in the prehistoric period from similar information.
Ritual tooth ablation, involving the deliberate extraction of frontal teeth, has been documented in prehistoric populations in Northwest Africa (Briggs, 1955), Italy (Robb, 1997), China (Han and Nakahashi, 1996), Thailand (Tayles, 1996), and Japan (Koganei, 1918; Watanabe M., 1966). In ethnographic records, ritual tooth ablation is performed on occasions varying from coming-of-age and marriage ceremonies to mourning for the death of a family member (Harunari, 1979).
It has been shown that, in Japan, the form of ritual tooth ablation changed through the Jomon period and into the Yayoi period (Watanabe M., 1966; Harunari, 1979, 1986). In Hokkaido, Honshu, and Kyushu in the Middle Jomon period (c. 5000–4000 years BP), one or two lateral maxillary incisors were extracted (Watanabe M., 1966; Harunari, 1986). From the Late Jomon to the Final Jomon period (c. 4000–2300 years BP), the removal of two maxillary canines plus two mandibular canines (type 2C) or four mandibular incisors (type 4I) was typically, and most actively, practiced in Honshu and Kyushu (Harunari, 1979). In the Yayoi period (c. 2300–1700 years BP, with an agricultural culture), extraction of the maxillary lateral incisors increased, suggesting a Chinese influence (Han and Nakahashi, 1996). The variety of forms and the frequency of extraction in the Late–Final Jomon period are high compared with that recorded in the ethnographic data for Taiwan (Watanabe M., 1966). Tooth filing of the upper maxillary incisors is also found in Osaka and Aichi Prefectures during the Final Jomon period, and individuals with such filing are thought to be the leaders of a group (Harunari, 1986). Hasebe (1919) suggested that tooth ablation was a customary component of coming-of-age ceremonies among the Jomon people. This is supported by Watanabe M. (1966), Fujita (1997), and Funahashi (2003) on the grounds of estimated age at the time of tooth ablation.
Knowledge of the social structure of the Jomon people is important for the interpretation of ritual tooth ablation. Harunari (1979) classified ritual tooth ablation into five types, with type 0 (see Figure 1) denominating coming-of-age ablation. The people comprising a single community were also classified into types: type 4I people are local to the settlement, type 2C people are migrants from another settlement through marriage to type 4I people, and type 4I2C and type 2C2I are people who have remarried. It was because type 4I people are frequently buried with personal ornaments, such as hip accessories or shell bracelets, suggesting greater prestige, that Harunari identified them as original members of the community. A cranial nonmetric trait study provided support for this hypothesis (Mouri and Oku, 1998). However, Harunari’s hypothesis has been questioned by Funahashi (2003) on the grounds that the tooth ablation took place before marriage and child birth. These suggestions remain hypothetical, and more investigations employing a variety of methods are needed in order to clarify the social function of ritual tooth ablation.
 View Details | Figure 1. Ritual tooth ablation types, after Harunari (1979). |
In the present study, we report the first stable isotope data from human and nonhuman mammal skeletal remains at the Yoshigo and Inariyama sites, as described above. Individual dietary differences can be inferred from the isotopic values, and we analyze their relation to sex, ritual tooth ablation characteristics, age at death, tooth filing, personal ornaments, and burial posture. The Yoshigo and Inariyama isotopic values are also compared with those from other Jomon populations, and differences and similarities in the protein food sources indicated by the data are discussed. Possible inferences from the isotopic values as to the social function of ritual tooth ablation are also raised and discussed.
Skeletons from the Yoshigo and Inariyama shell mounds dated to the later part of the Late Jomon period to the Final Jomon period (c. 3500–2300 years BP) were subjected to stable isotope analysis. The sites were dated using radiocarbon analysis and pottery-based chronology. Pottery chronology is an accurate method for estimation of the date of sites in the Jomon period because of the multiplicity of systematic studies of the chronology of Jomon pottery (see Imamura, 1996).
The Yoshigo shell mound is located in Tahara Town, Atsumi County, Aichi Prefecture, on the coast of the Bay of Mikawa (Figure 2). This site is dated as extending from the later part of the Late Jomon period (c. 3500–3000 years BP, as characterized by the Yoshigo-Kaso pottery type; Hisanaga, 1952) to the Final Jomon period (c. 3000–2300 years BP; Gokanmori pottery type: Sugihara and Toyama, 1964). The radiocarbon dates of two shells from the Yoshigo shell mound have been reported: 2870 ± 250 and 2800 ± 600 uncalibrated years BP (Watanabe N., 1966). In 1922, K. Kiyono excavated this site and recovered 304 human skeletons (Kiyono, 1969), and ritual tooth ablation can be recognized in about 100 individuals. The Inariyama shell mound is located in Kozakai Town, Hoi County, Aichi Prefecture (Figure 2). This site is dated to the middle of the Final Jomon period by the Inariyama pottery type (c. 2800–2500 years BP; Sugihara and Toyama, 1964). The radiocarbon dates of remains from the Inariyama shell mound have not been investigated. In 1922, the site was excavated by K. Kiyono and 53 human skeletons were recovered (Kiyono, 1969). These skeletal collections are stored in the Laboratory of Physical Anthropology, Department of Zoology, Graduate School of Science, Kyoto University. Because the Inariyama pottery type is also found in the Yoshigo shell mound (Sugihara and Toyama, 1964), the Inariyama people can be expected to be contemporary with the Yoshigo people.
 View Details | Figure 2. Geographical location of the Yoshigo and Inariyama shell mounds. |
The present study analyzed bone samples from a sample of 38 human skeletons from the Yoshigo shell mound and from 28 from the Inariyama shell mound, and also from two skeletons of Japanese macaque and two dogs from the Yoshigo shell mound. We made use of the ritual tooth ablation classification into five types (Figure 1) proposed by Harunari (1979), as follows: Type 0 (zero): removal of the two maxillary canines; Type 4I: removal of the four mandibular incisors in addition to the two maxillary canines; Type 2C: removal of the two mandibular canines in addition to the two maxillary canines. An additional two ablation types, which derive from type 4I and type 2C, respectively, were identified in the Yoshigo shell mound. Type 4I2C: removal of the four mandibular incisors and the two mandibular canines in addition to the two maxillary canines; and Type 2C2I: removal of the two mandibular canines and the two central mandibular incisors in addition to the two maxillary canines.
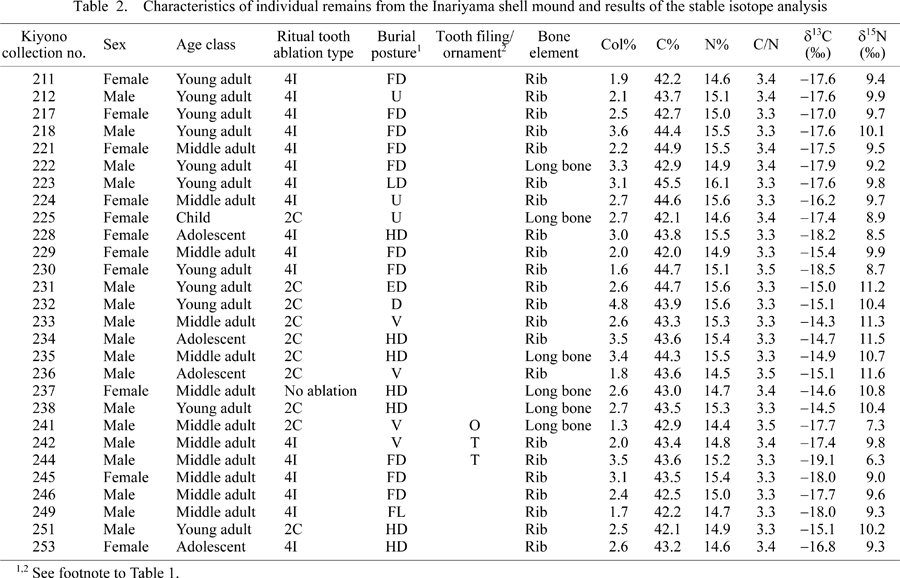
Bone samples were taken from the ribs of the Yoshigo skeletons and from the ribs or long bone shafts of the Inariyama skeletons. Extraction of bone collagen was carried out in the Laboratory of the Research Institute for Humanity and Nature. Extraction followed the procedure described by Yoneda et al. (2004). About 1 g of bone sample was ultrasonically cleaned in pure water in order to remove any soil. Humic and fulvic acids were removed from the sample by soaking in 0.2 M NaOH overnight and the sample was neutralized in pure water. The sample was lyophilized and crushed by a mill. The sample was then sealed in a cellulose tube and soaked in 1 M HCl overnight to remove hydroxyapatite. The remaining material was separated by centrifugation and lyophilized. The sample was heated at 90°C for 12 h in pure water. Both fractions were freeze-dried, and extracted gelatin samples were used for isotopic analyses.
About 0.5 mg of the extracted collagen was measured for carbon and nitrogen isotopic ratios, using an elemental analyzer (FLASH EATM) connected to an isotope ratio mass spectrometer (DELTA plus XPTM). The natural abundances of 13C and 15N are expressed as per mil (‰) deviation from international standards: δ13C or δ15N = (Rsample/Rstandard − 1) × 1000, where R in δ13C or δ15N is 13C/12C or 15N/14N, respectively. Pee Dee Belemnite and atmospheric nitrogen were used as the international standards for carbon and nitrogen, respectively. Based on the standard materials measured with unknown samples, the measuring errors for each measurement were less than 0.2‰ for δ15N values and 0.2‰ for δ13C values.
An analysis of covariance (ANCOVA) was used to test for linear regression between δ13C and δ15N values and differences in the slopes of the regression lines for the Yoshigo and Inariyama data. Differences between the isotopic values for the Yoshigo and Inariyama subject groups, and between two subgroups defined by sex and by ablation types 4I and 2C, were analyzed using a two-sample t-test, and equality of variance in the groups was tested by an F-test. Differences between isotopic values in further subgroups based on the four ritual tooth ablation types and burial posture were analyzed by one-way analysis of variance (ANOVA).
The carbon and nitrogen stable isotope ratios of all the subjects’ bone samples are listed in Table 1, Table 2, Table 3. Based on the following three criteria, it was confirmed that diagenetic effects did not influence these ratios: first, C/N ratios are within the range 2.9–3.6 (DeNiro, 1985); second, concentrations of carbon and nitrogen in the sample were greater than 13% and 4.8%, respectively (Ambrose, 1990); and third, the recovery rate of collagen was more than 1% (Van Klinken, 1999). All samples met these criteria.

The carbon and nitrogen isotope ratios obtained are plotted in Figure 3. The average δ13C value at the Yoshigo site was −16.3 ± 1.7‰ (mean ± 1 SD), and the average δ15N value was 10.8 ± 1.9‰. The average δ13C value at the Inariyama site was −16.7 ± 1.4‰, and the average δ15N value was 9.7 ± 1.2‰. The differences between the average δ13C and δ15N values at the Yoshigo and Inariyama sites were not significant.
 View Details | Figure 3. Regression lines of isotopic values from stable carbon and nitrogen isotope analysis of human bones recovered from Yoshigo (unbroken line) and Inariyama (dotted line) and nonhuman mammal bones recovered from Yoshigo. Data of estimated protein sources (Marine finfish, Marine shellfish, Terrestrial herbivore, and C3 plants) from Yoneda et al. (2004): means ± 1 S.D. |
δ13C and δ15N values for both sites were linearly correlated, and the slopes of the regression lines at the two sites differed significantly (ANCOVA for δ15N with site as independent variable and δ13C as covariate: interaction of δ13C and site effect: F = 22.4, P < 0.001; site effect: F = 27.5, P < 0.001; δ13C effect: F = 346.7, P < 0.001). The Yoshigo people showed a significantly steeper slope than the Inariyama people.
The isotopic values of potential protein sources (C3 plants, terrestrial herbivores, marine finfish, and marine shellfish) are also shown in Figure 3. The specific protein sources were selected based on the evidence of organic remains collected from both sites (Yawata, 1952; Nakamura and Deguchi, 1992). C4 plants do not appear in Figure 3 because there is no archaeological evidence suggesting significant consumption of C4 plants. Isotopic values of the protein sources are cited from Yoneda et al. (2004). In the process of assimilation, heavy isotopes accumulate in human tissue, and isotopic enrichment of the δ13C or δ15N values in collagen occurs (DeNiro and Epstein, 1981; Ambrose and DeNiro, 1986). Therefore, 4.5‰ for the δ13C values and 3.4‰ for the δ15N values (Ambrose, 1993) were added to the isotopic values of the protein sources. The isotopic values of bone collagen at both sites were distributed within the range of the isotopic values of the selected protein sources.
The δ13C and δ15N values found in the bones of the two dogs were very similar to those in the human bones, while the δ13C and δ15N values in the bones of the two macaques were distributed within the range of isotopic values of C3 plants. Because dogs are omnivorous and macaques are herbivorous, these results indicate that the isotope measurements and the isotopic values of the protein sources examined were valid.
The comparison of stable isotope ratios in the subgroups based on age at death, presence of tooth filing, and personal ornaments at the two sites produced no significant differences. However, an interesting difference was found in the comparison on the basis of sex. While there were no significant differences in average δ13C and δ15N values between the sexes at either site, the SDs of the δ15N values were significantly different for males and females (F-test, P < 0.05; Figure 4). The SDs of the δ15N values of males were higher by 0.7‰ than those of females at both sites.
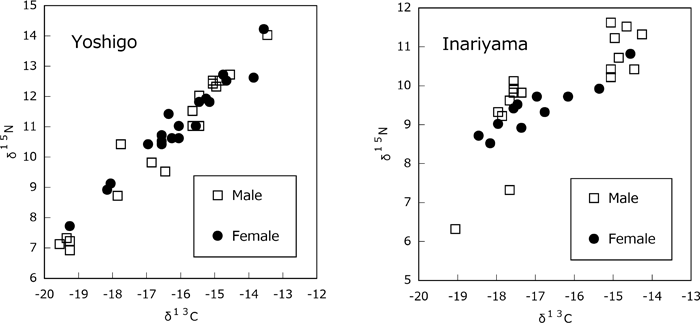 View Details | Figure 4. Stable carbon and nitrogen isotope ratios of the Yoshigo and the Inariyama samples, categorized by sex. |
At the Inariyama site, type of ritual tooth ablation was differentially associated with isotope ratio (Figure 5). Individuals with type 2C tooth ablation had significantly higher δ13C and δ15N values than individuals with type 4I ablation (t-test, P < 0.01 for δ13C; P < 0.05, for δ15N), and a similar difference occurred in relation to burial posture. Isotopic ratios of δ13C and δ15N values were significantly higher for extended dorsal burial and ventral burial than for flexed dorsal burial (ANOVA, P < 0.05). These associations were not identified in the Yoshigo sample.
 View Details | Figure 5. Stable carbon and nitrogen isotope ratios of the Yoshigo and Inariyama samples, categorized by ritual tooth ablation type. |
The average δ13C and δ15N values from the Yoshigo and Inariyama sites are very similar to those of other coastal Jomon populations in Honshu and Kyushu, but lower than those from the Kitakogane and Takasago sites of Hokkaido Jomon (Minagawa, 2001). It is considered that high isotopic ratios, as found in the Hokkaido Jomon populations, reflect a diet more heavily dependent on marine fauna than on land mammals (Minagawa, 2001). Inland Jomon populations such as the Boji (Yoneda et al., 2004) and the Tochibara (Yoneda et al., 2002) have lower isotopic ratios again, compared with the Honshu and Kyushu coastal Jomon populations. This trend is explicable as resulting from a relatively low consumption of marine food resources at inland sites. There are no features peculiar to the animal remains found at Yoshigo and Inariyama, compared to other Honshu and Kyushu coastal Jomon sites (Yawata, 1952; Nakamura and Deguchi, 1992), so it is not unexpected that Yoshigo and Inariyama populations should have isotopic ratios similar to those of other coastal Jomon communities in Honshu and Kyushu.
An interesting finding is the greater degree of dietary variability in the Yoshigo and Inariyama samples compared with other Jomon populations. The SDs of both the δ13C and the δ15N values at both sites were greater than 1‰. These values are surprisingly high, as SDs for Jomon sites are usually less than 1‰, with only a few exceptions greater than 1‰ (Minagawa, 2001). The high variability observed at these sites presumably reflects atypical individual differences in the daily protein consumption sources. Nevertheless the δ13C and δ15N values correlated positively at both sites (Figure 3). This correlation suggests that the Jomon people at these sites mainly consumed two types of food as their main protein source, but with individual differences in the proportions consumed, these being marine resources (marine finfish and shellfish) vs. terrestrial resources (C3 plants and terrestrial mammals). Positive linear correlations between δ13C and δ15N values are also reported from other Jomon populations (Minagawa, 2001), from southern Californian Native Americans (Walker and DeNiro, 1986), and from coastal Mesolithic people in Europe (Richards and Hedges, 1999). This correlation is probably characteristic of populations that have access to both marine and terrestrial food resources.
The slopes of the regression lines of δ13C on δ15N were significantly different in the Yoshigo and Inariyama data, being steeper in the former (Figure 3). The Yoshigo people might in general have consumed proportionally more marine finfish, which has higher δ15N values, than marine shellfish, while the Inariyama people ate more marine shellfish than finfish. This explanation is in keeping with the archaeological evidence that only a few fragments of marine finfish were recovered from the Inariyama shell mound (Nakamura and Deguchi, 1992), and many kinds of marine finfish were found in the Yoshigo shell mound (Toizumi and Yamasaki, 2007). However, a quantitative comparison of marine finfish and shellfish remains at the Yoshigo and the Inariyama sites is needed to substantiate this explanation.
Furthermore a study of carious tooth and enamel hypoplasia in these two samples revealed a lower frequency of carious tooth and a higher frequency of enamel hypoplasia in the Inariyama people than in the Yoshigo people (Temple, 2007). This suggests that the Inariyama people suffered greater stress from seasonal resource depletion, while the Yoshigo people compensated with consumption of plant foods such as starchy roots, i.e. that environmental food availability affected their diets differentially.
We have interpreted the variability of isotopic data as reflecting individual variability of diet within the sites, but this variability could in fact reflect changes in diet over time resulting from environmental change during the Late–Final Jomon period. Pottery based chronology indicates that remains were accumulated for at most 1200 years at the Yoshigo site and for 300 years at the Inariyama site. The quality and quantity of natural food resources may have changed because of environmental cooling during the Late–Final Jomon period (Tsukada, 1986). If diets changed in association with environmental change, then the variations in isotopic values we describe would not reflect individual dietary differences throughout the period, but rather changes in diet during the Late–Final Jomon period. Radiocarbon dating of the Yoshigo and Inariyama skeletons is needed in order to evaluate the time depth of these samples. Furthermore, environmental change as reflected in animal skeletal remains over the period should be investigated.
An important finding of this study is the effect of sex on variability of individual diets. Although there was no difference by sex in average δ13C and δ15N values, the SD of δ15N values in males was significantly greater than in females at both sites (Figure 4). This indicates that male diets differed more widely between individuals than did female diets.
It is largely accepted in anthropology that a sexual division of labor and food sharing between males and females is fundamental to hunter-gatherer subsistence economies (Ember, 1975; Bird, 1999), with the result that dietary differences by sex are not expected. Contrary to this expectation, variability in individual diets differed by sex in our Yoshigo and Inariyama samples. This may be because males were predominantly engaged in hunting or in fishing away from the village, and these males consumed large quantities of the foods acquired by their hunting or fishing while away. If so, then the larger SD of δ15N values in males than in females reflects not only the sexual division of labor, but also occupational differentiation in males in the Yoshigo and Inariyama populations.
Reports from other Jomon sites have also suggested differences between the sexes in dietary intake. At the Kitamura site in Nagano, males had a higher average δ13C value than females (Yoneda et al., 1996), while in Hokkaido, females had a higher average δ13C value than males (Chisholm et al., 1992). The Hokkaido Jomon finding was explained as resulting from the sexual division of labor, whereby hunting was exclusively performed by males and fishing and plant gathering by females (Chisholm et al., 1992). It is not unreasonable that different environmental settings and differences in food resources should promote differences in the division of labor both between and within the sexes, and in turn result in differences in dietary profile between and within the sexes.
The most important finding of the present study is the relationship between ritual tooth ablation type and δ13C and δ15N values in the Inariyama population. Average δ13C and δ15N values differed significantly between individuals with type 4I and type 2C ablation (Figure 5). These differences suggest that type 4I people were hunters and/or gatherers and more dependent on terrestrial resources, and type 2C people were fishers and more dependent on marine resources. The isotopic values of type 4I males are also clearly different from those of type 2C males (Figure 6). Unfortunately the number of type 2C females in our sample was too small for a reliable comparison with type 4I females. In hunter-gatherer subsistence societies, males usually hunt larger game while females gather plants and catch smaller animals (Bird, 1999). In males, at least, a differentiation in ritual tooth ablation associated with hunting vs. fishing as a male occupation might have occurred. Ethnographic observations of the Ainu record the presence of hunting-oriented and fishing-oriented families (or lineages) in the same population (Watanabe H., 1972, 1983, 1990). A similar occupational differentiation based on family and distinguished by ritual tooth ablation type might also have occurred in the Inariyama people.
 View Details | Figure 6. Stable carbon and nitrogen isotope ratios of the Inariyama sample, categorized by sex and ritual tooth ablation type. |
Mizoguchi (2002) hypothesized that in Jomon societies the seasonal economies were associated with different procurement groups, membership of which endowed social identity. The present study supports this hypothesis with its evidence that the social identity endowed by ritual tooth ablation type relates to subsistence procurement group.
Occupational differentiation in Ainu society was tied to religious differentiation and social status (Watanabe H., 1972). Hunting-oriented families were more prestigious than fishing-oriented families, in that hunting-oriented families owned the ritual of the brown-bear deity, which was the Ainu’s highest-ranking deity. Harunari (1979, 1986) proposed that type 4I people were more prestigious than 2C people on the basis of their personal ornaments, and the results of the present study suggest that type 4I people were the hunters among the Inariyama people. The Inariyama people and the Ainu people may have shared a common feature, as postulated by Watanabe H. (1990), that hunting-oriented families are more prestigious than fishing-oriented families.
Individuals 241 and 244 from the Inariyama site had unusually low δ13C and δ15N values compared with other individuals of the same tooth ablation type from the same site (Figure 6). Ornaments were found with the remains of individual 241, and individual 244 showed filing of his anterior dentition. Both may be signs of a special social status, which may have resulted in dietary differences compared with ordinary members of the society.
A relationship between ritual tooth ablation type and diet did not appear in the Yoshigo data. A possible explanation is that ritual tooth ablation was primarily indicative of kinship, and not of subsistence procurement group, as postulated by Tanaka (1998) and Funahashi (2003). Studies of nonmetric cranial variants of the Yoshigo and Inariyama people (Mouri and Oku, 1998) and nonmetric tooth variants of the Inariyama people (Hashimoto and Baba, 1998) indicate the possibility that type 4I people are hereditarily different from type 2C people, in which case ritual tooth ablation type was a signifier of social identity in terms of kinship group. In Ainu society, there was lineage-based division of labor and both intrafamily and inter-lineage cooperation as regards access to food resources (Watanabe H., 1972). Cooperation was usually between families in the same settlement group.
We suggest that among the Yoshigo people, families that shared the same type of ritual tooth ablation would cooperate with other families regardless of type of tooth ablation; whereas among the Inariyama people, type 4I families shared food resources only with other type 4I families, and type 2C families cooperated only with other type 2C families. Thus, our findings of differing isotopic values may not, in fact, indicate that subsistence procurement group determined ritual tooth ablation and social identity among the Inariyama people, but not among the Yoshigo people: the determining factor may have been lineage in both groups.
It has been proposed that ritual tooth ablation in Jomon culture was a coming-of-age ceremony (Hasebe, 1919; Watanabe M., 1966; Fujita, 1997; Funahashi, 2003), based on observations that it is almost always present in adults and never in children. The typology of ritual tooth ablation indicates additional meanings. The findings of the present study, while consistent with the coming-of-age theory, suggest that ritual tooth ablation typology identified a lineage-based subsistence procurement group.
Harunari (1979) hypothesized that type 4I people were local to the Yoshigo and Inariyama settlements, and type 2C people were migrants to the settlements through marriage with type 4I people, a suggestion radically at odds with our own conclusions. Although studies of nonmetric cranial variants (Mouri and Oku, 1998) and nonmetric tooth variants (Hashimoto and Baba, 1998) suggest that type 4I people are hereditarily different from type 2C people, an analysis of dental and cranial measurements in the Yoshigo people indicates similar parameters and mobility patterns in type 4I and type 2C individuals (Temple and Sciulli, in press). Funahashi (2003) found that extraction of maxillary and mandibular canines and mandibular incisors occurred before marriage and childbirth. These studies and our conclusions refute the possibility that ritual tooth ablation signifies marriage patterns.
The authors wish to thank M. Yoneda (The University of Tokyo) for instructing us in the experimental methodology of this study, M. Nakatsukasa (Kyoto University) for his insightful comments and editing of the manuscript, members of the Laboratory of Physical Anthropology, Kyoto University for their helpful discussions, D.H. Temple (The Ohio State University) for his helpful comments, and members of Project D-02 of the Research Institute for Humanity and Nature for the help they gave with this study. The study was conducted as a part of Project D-02, ‘A New Cultural and Historical Exploration into Human-Nature Relationships in the Japanese Archipelago’ at the Research Institute for Humanity and Nature.
|