| Correspondence to: Etty Indriati, Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, Gadjah Mada University, Yogyakarta, Indonesia. E-mail: ettykurtz@idola.net.id Published online 21 May 2008 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.070814 |
Hominins first dispersed from Africa to Eurasia in the earliest Pleistocene (Swisher et al., 1994; Gabunia and Vekua, 1995; Gabunia et al., 2000). The identity of these dispersers, their relations across geographic regions, and the factors that facilitated dispersal remain a source of great interest (Antón et al., 2002). The evidence for dispersal rests on those early Pleistocene fossil hominins found outside Africa. Here we describe the Homo erectus cranium, Sangiran 27. This cranium represents the only substantial facial remains recovered from the earliest Pleistocene sediments of the Sangiran Formation, Java. Although other cranial remains with associated facial elements have been found in the Sangiran Formation, such as Sangiran 4 which preserves the palate, Sangiran 27 is the only extensive representation of upper (e.g. supraorbital) and middle (e.g. maxillary, zygomatic, pyriform aperature) facial features from the older chronostratigraphic levels at Sangiran.
The Sangiran region of central eastern Java is a dome-shaped anticline formed between the volcanoes Mount Lawu and Mount Merapi. Four formations are exposed in the region, designated from lower to upper stratigraphic levels, the Puren (or Kalibeng) Formation, the Sangiran (or Pucangan) Formation, the Bapang (or Kabuh) Formation, and the Pohjajar (or Notopuro) Formation (Figure 1). Terrestrial fauna come principally from the Bapang Formation, comprised largely of cross-bedded sandstones. The underlying Sangiran Formation is characterized by black clays (Watanabe and Kadar, 1985), and the Puren Formation is largely marine. The sequence is dated by a series of 40Ar/39Ar ages of tuffs situated throughout the four formations (Swisher et al., 1994; Swisher, 1997; Larick et al., 2001), by microfossil correlation (Siesser and Orchiston, 1978; Ninkovich et al., 1982; Orchiston and Siesser, 1982), and by the geomagnetic polarity of the deposits (Hyodo, 2001). The earliest vertebrate remains from the dome are about 2 million years old, but the oldest hominins are found only in the uppermost levels of the Sangiran Formation, overlain by the Grenzbank, the latter dated to 1.58 Ma and underlain by Tuff 10 dated to 1.66 Ma (Swisher et al., 1994; Swisher, 1997). Despite frequent discussions regarding the accuracy of the provenance of the Sangiran fossil hominins (e.g. Larick et al., 2001), the taphonomic and diagenetic influences on the fossils differ dramatically between Bapang and Sangiran Formation faunas; those recovered from the Bapang sandstones are undeformed and well preserved, whereas many of those recovered from the black clays of the Sangiran Formation are highly deformed and telescoped within their surrounding matrix. These differences and the preservation of the Sangiran crania suggest that fossil hominins were present in Java during the time of the Sangiran Formation (see Antón and Swisher, 2004; Indriati, 2004).
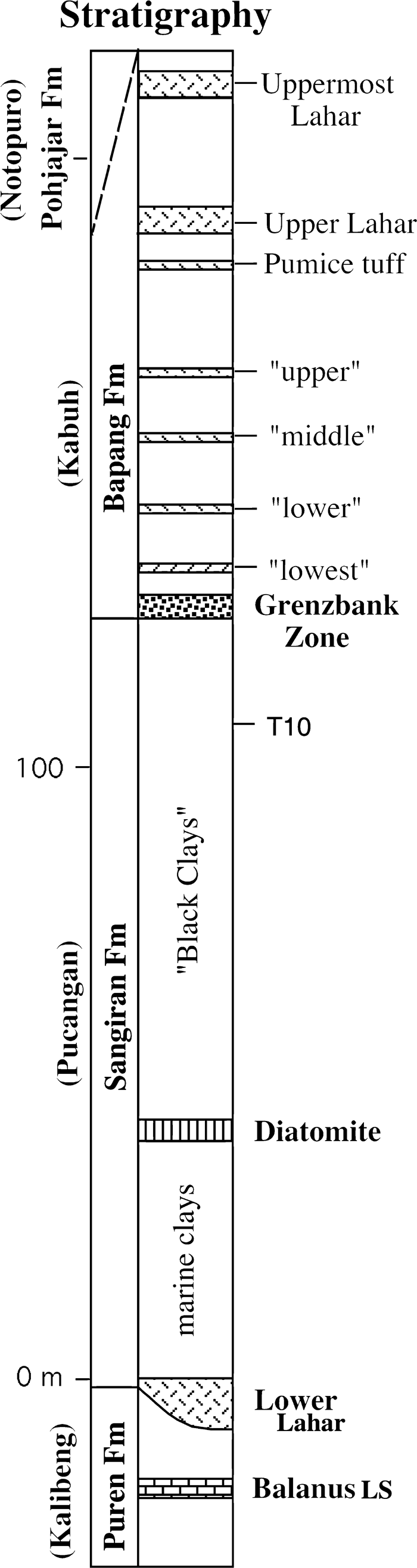 View Details | Figure 1. Schematic stratigraphic section of the Sangiran Dome region including four formations and their subdivisions. Sangiran 27 was probably recovered from above Tuff T10 and below the Grenzbank Zone. Upper, middle, lower, and lowest refer to tuffs in the section. Figure courtesy of Dr. C.C. Swisher, III. |
The absolute age of the Sangiran Formation remains a source of some argument (Langbroek and Roebroks, 2000). Although two independent groups have published a series of 40Ar/39Ar ages on tuffs situated throughout the four formations (Swisher et al., 1994; Swisher, 1997; Larick et al., 2001) that are in agreement with paleomagnetic and microfossil age estimates, faunal correlations provide a younger age of about 1.2 Ma for the upper portion of the Sangiran Formation associated with the hominin remains (e.g. Sondaar et al., 1996), and the paleomagnetic evidence has also been interpreted to be consistent with this younger chronological scale (e.g. Hyodo, 2001). However, as argued more extensively elsewhere, the calibration points for these faunas are not well established and where they do rest on radiometric techniques it is on fission-track ages for tektites, the provenience of which cannot be established (see Larick et al., 2001; Antón and Swisher, 2004). With increasing confidence, we consider that the age of the Sangiran Formation is most conservatively placed in the earliest part of the Early Pleistocene.
Of the many adult partial crania from Sangiran, Sangiran 4, 27 and 31 probably derive from the Sangiran Formation. Of these, both Sangiran 4 and 31 preserve portions of the posterior cranial vault, whereas only Sangiran 27 retains the anterior vault and face. In fact, Sangiran 27 is one of only three specimens with substantial facial portions known in the Indonesian fossil hominin record, the others being the Bapang Formation hominins Sangiran 17 and Sangiran IX (Sartono, 1972; Sartono and Tyler, 1993; Arif et al., 2001, 2002). Sangiran 27 was recovered in 1978 during construction of a dam in the black clays of the Sangiran Formation. The cranium has been mentioned previously, but not fully described or compared (Jacob, 1983; Antón and Indriati, 2002; Schwartz and Tattersall, 2003; Antón, 2003; Indriati, 2004; Kaifu et al., 2005).
We use the descriptive format of Wood (1991) and the terminology of Weidenreich (1943) and Wood (1991). Occlusal wear and accessory cusp development is scored following the Arizona State University Dental Anthropology System (ASUDAS; Turner et al., 1991). Because of the poor preservation of much of the preserved cranial vault and face, we provide comparative evaluations primarily on the relatively well preserved supraorbital torus and dental morphology.
Sangiran 27 is a highly mineralized partial cranium that includes portions of the face, anterior vault, and maxillary dentition (Figure 2). The fossil has undergone substantial postmortem crushing and deformation, including the telescoping of superior structures onto the more inferior structures, and substantial dorsoventral shear that affects the right side more than the left. The crushed elements are cemented together into four sections, two that are primarily composed of left and right temporal bones, and two fine-grained blocks of matrix, each containing a number of cranial elements. These two blocks conjoin at the medial portion of the left supraorbital torus and at the zygomatic process of the left maxilla. Secondary mineral formation within the bones has caused additional separation along sutures and postmortem fracture lines. The close approximation of the densities of the fossil and its matrix has, so far, precluded the use of computed tomography to generate a three-dimensional reconstruction/restoration (J.J. Hublin, personal communication).
 View Details | Figure 2. Sangiran 27 cranium. (a) Facial view; (b) right lateral view, note where the two major blocks of fossilized bone join at the medial end of the left supraorbital torus and along the left maxilla; (c). left lateral view; and (d) mastoid process. |
Sangiran 27 retains portions of the frontal area, including most of the supraorbital torus and the frontal squama on both sides, portions of both zygomatics, including nearly the entire left zygomatic, portions of both maxilla, including substantial right and left alveolar portions, the left horizontal shelf of the hard palate, the inferior lateral edges of the pyriform aperture on both sides, the right sphenoid and palatine at the inferiormost pterygoid plate (where they join the posterior alveolar process of the maxilla), and portions of the left parietal and temporal including the mastoid process. Most of the central face is absent although portions of the nasals and frontal processes of the maxilla are preserved but substantially deformed. In contrast, the postcanine maxillary dentition is well preserved, retaining P3 to M3 on the right side and P4 to M3 on the left side, although the third molars are badly damaged and/or displaced (Figure 3). Portions of both canines, including the roots are present.
 View Details | Figure 3. Palate of S27 showing details of upper dentition. Note that the second fossil block holds the left third molar. Also note that palate shape is distorted by postdepositional deformation. The crushed enamel of the RM3 is visible approximately 15 mm posterior of the M2 and just posterior of a large mediolateral crack in the block. The crack is indicated by a dark arrow-head. |
Frontal
From the right side to the left side near the midline, a portion of the frontal squama, some 40 mm long, is preserved and it is largely undeformed (Figure 2). The frontal squama is thick; it measures 12 mm in thickness at the posterior end of the medial supratoral sulcus, and 9.25 mm further superiorly. Laterally, the frontal squama is 14.3 mm thick posterior to the supratoral sulcus, a value that may be slightly low due to an incomplete endocranial surface. Overall the ectocranial table is thicker than the endocranial table. At the left lateral supraorbital torus the ectocranial table is 3 mm thick, the endocranial table is 1.3 mm thick, and the remainder of thickness (11 mm) is diploic bone. The left temporal line is preserved as a moderately raised ridge that extends from the back of the lateral supraorbital torus and runs medially and then posteriorly. The extent of the incurvature of this line from the lateral supraorbital torus suggests moderate postorbital constriction, probably similar to that seen in Sangiran 17.
Nearly the entire supraorbital torus (SOT) is preserved (Figure 2). The left side is virtually undeformed with only several short, superoinferiorly oriented cracks that do not show evidence of secondary mineralization, whereas the right SOT experienced substantial postmortem crushing superoinferiorly and anteroposteriorly. The rim of the left orbit is more complete and less deformed than the right side, whereas more of the orbital roof and posterior aspects of the right orbit are present but crushed and enclosed in matrix. The left supraorbital torus forms the basis of this description.
The torus is continuous and thick (Table 1) being negligibly thicker at midorbit than laterally. The internal structure can be observed at its medial broken edge, where only diploe and no sinus is visible; however, the location of the break is likely to be lateral to the possible extent of a frontal sinus, and therefore we cannot determine whether or not a confined frontal sinus existed.
In overall form the supraorbital torus appears to have been bar-like, but this observation is complicated by crushing on the right side. Several observations related to the supraorbital torus, such as the depressio glabellaris, an inferior indentation at glabella, cannot be made due to the poor state of preservation. Approximately 35 mm from the left lateral edge of the orbit (just medial of midorbit) there is a small supraorbital notch that is nearly enclosed into a foramen. Lateral to this on the anterior face of the supraorbital torus there is a small, probably pathological, ovoid depression that is bilaterally expressed and broader than it is tall (approximately 8.5 mm × 5.0 mm). Posterior to the lateral supraorbital torus and medial to the temporal line, a shallow, wide, continuous depression (supratoral sulcus/postoral gutter) is present. This sulcus narrows and deepens towards the midline.
Temporal
On the left side, posterior and superior to the maxilla, a portion of the anterior temporal squama, articular plane, anterior articular eminence, and root of the zygomatic process of the temporal are preserved (Figure 2c). The entire piece measures approximately 22 mm anteroposteriorly and 23 mm superoinferiorly. Remnants of the anterior sphenotemporal suture are present. The glenoid fossa is incomplete medially but appears to have been mediolaterally short. The anterior wall of the glenoid fossa faces posteriorly and is steeply angled from the articular plane. The fragment can be seen in the coronal section posteriorly. Matrix infillings of the endocranial cavity suggests that about half of the glenoid fossa was positioned lateral to the endocranial cavity. The thickness of the ectocranial table of the temporal squama varies between about 2.5 and 2.8 mm. Endocranial thickness is obscured by matrix.
Two separate pieces preserve portions of the deformed left and right posterior temporal bones. The preservation of the mastoid process is better than that of the rest of the skull and face. The right temporal fragment measures 73 mm anteroposteriorly and 65 mm superoinferiorly, and the left temporal measures 68 mm × 45 mm. The total outline of the mastoid processes is observable (Figure 2d). The right mastoid process is more intact than the left, because the left side is eroded on the inferior third of the mastoid tip. The mastoid process is robust, and is oriented anteromedially. They are taller superoinferiorly (~30 mm) than long anteroposteriorly (~20 mm). The apex of the mastoid process is well developed and individualized. The digastric groove occurs medially, and is wide mediolaterally (this is clearly observable on the left side, but partially covered by matrix on the right side, giving an impression of narrowness). The left digastric groove is 6.2 mm wide and 5.7 mm deep. Above the position of the left digastric groove, and about 12 mm medially, a drill hole is present, with a diameter of 5.3 mm and a depth of 7.8 mm (this hole is approximately at the level of the sigmoid sinus and its origin is unknown). The right temporal retains a segment of the petrous portion and a centrally shrunken sigmoid sulcus. The petrous lacks an arcuate eminence and the superior body is flat (see also Schwartz and Tattersall, 2003). Air cells are present just above the petrous pyramid. A bit of the posterior portion of root of the zygomatic process is also present. Bone thickness at the parietomastoid suture is 11.9 mm and 12.5 mm on right and left sides, respectively.
Parietal
On the large, multi-element block from the left side, there is a small piece of parietal visible in posterior view, located medial and somewhat superior to the temporal piece described above. The position of the parietal piece suggests that it is likely a portion near the squamosal suture that was telescoped inferiorly toward the temporal. Except for its existence, the piece is unremarkable. Postmortem crushing precludes thickness measurements.
Nasoalveolar clivus and pyriform aperture of the maxilla
Both right and left maxillae preserve the inferior lateral margins of the nasal aperture, to a position about 7 mm superior to the nasal floor. The subnasal area is preserved for approximately 15 mm inferior to the nasal aperture. Neither side preserves the anterior subnasal midline. Although the nasal aperture is largely filled with matrix, the lateral and inferior aperture margin is clean of matrix anteriorly (particularly on the left side), revealing a smooth transition between the clivus and nasal floor. Below this, the lateral aspect of the nasoalveolar clivus appears slightly concave to flat. However, in the absence of an intact midline region, we cannot exclude the possibility that the nasoalveolar clivus might have been more transversely convex in overall profile (see below).
Lateral to the nasal aperture there is no evidence of the flattened pillars seen in both KNM-WT 15000 and Dmanisi 2282 (sensu Rightmire, 1998). Rather, the better-preserved right side suggests that the canine root is angled posteriorly and away from the lateral margin of the nasal aperture (the position of the right canine root does not appear to be affected by postmortem deformation since the buccal alveolar surface is largely undeformed). The position of the canine suggests that subnasal prognathism was stronger than it seems as preserved, and suggests the likelihood of a convex nasoalveolar clivus.
Maxillary incisure region and maxillary sinus
Both sides preserve a portion of the root of the zygomatic process of the maxilla still attached to the buccal alveolar surface. Preservation is more extensive on the right side than the left side. Both sides show evidence of a maxillary incisure, based on the position of the zygomatic root relative to the alveolar process. In addition, the right side provides evidence of the form of this incisure. Although the bulk of the right zygomatic process of the maxilla is separated superiorly and laterally from the zygomatic root area by about 20 mm (postmortem sediment infill and secondary mineralization at the fractures), little plastic deformation is apparent and none to the maxillary incisure region itself. As a result, the maxillary incisure can be relatively accurately visualized to form a strong, nearly 90° angle curvature.
In lateral view, on both sides, the center of the zygomatic root is located over M1. The anterior surface of the zygomatic root is not preserved but could have extended slightly more anteriorly to a position above P4–M1. On the right side, the undeformed distance from the alveolus to the beginning of the zygomatic root is slightly more than 8 mm. On both sides, the buccal surface of the alveolar process is slightly depressed, this weak concavity extending anteroposteriroly from canine root to zygomatic root positions. This concavity is clearer on the right side than on the left side. The maxillary sinus is matrix-filled, which shows that it extended inferiorly below the level of the nasal floor. The posterior walls of the maxillae are present but are fractured into several pieces and flattened postmortem. Extensive secondary mineralization exists in between these fractured pieces. The orbital surfaces of the maxillae are not visibly preserved.
Zygomatic
The left zygomatic is more complete than the right side, retaining the zygomatic body and parts of all major processes. The right side retains only the body and a portion of the inferior orbital margin. On both the left and right sides, the inferior orbital margin is not sharp but rounded. Because of deformation, orbital height and breadth cannot be measured on either side. The better-preserved left zygomatic is displaced laterally and inferiorly relative to the superior orbital margin. Both sides exhibit a single, large zygomaticofacial foramen.
The body of the left zygomatic has a measured height of 25.7 mm as preserved, but this may be enlarged by as much as 4 mm due to superoinferior expansion by a series of horizontal cracks spread apart by secondary mineralization. The breadth of the frontal process of the left zygomatic is about 19.5 mm (inferior portion). The left zygomatic also preserves the anterior masseter attachment area, which is at least 12 mm thick and shows no sign of a maxillary tubercle. The left posterior zygomatic surface (temporal fossa surface) is largely undeformed and unfractured. Much of this surface is covered by a skin of matrix that prevents full observation on the existence and number of zygomaticotemporal foramina. No foramina are visible on the currently cleaned areas.
Palate and alveolar process
Anteriorly, the palate is ill preserved; the incisors and their alveoli are absent. Posteriorly, the alveolar processes are relatively complete. On the right side, the buccal alveolar surface of the postcanine region is intact, and there appears to have been little postmortem movement from C–M2. The lingual alveolar surfaces are less intact. There is obvious postmortem rotation of the right (R) P3 crown, accompanied by secondary mineralization between tooth roots and alveolar bone, resulting in increased interdental spacing. Distal to the RM2, the RM3 and its alveolus have been separated from the C–M2 portion. As preserved, the RM3 is placed approximately 15 mm directly posterior to RM2. It has fallen forward on its mesial face and then been subsequently crushed. Its distal surface enamel is broken, exposing dentine. The RM3 alveolar bone, including the posterior surface of the alveolar process, and adjacent portions of the sphenoid pterygoid plate are located another 5 mm posterior to the crushed M3 crown.
Unlike the right side, on the left side the C–M2 alveolus is cracked into multiple segments. Buccally, a fissure anterior to the left (L) M1 separates the C–M2 area into two main segments. Lingually, there are four segments that each hold a tooth crown. The segments (and teeth) are in line with one another but clearly separated by secondary mineralization within the fissures. Separation between segments ranges from 1.5 to 3.5 mm. Distal to LM2, the LM3 is displaced approximately 16 mm posteriorly. The LM3 is located on a second, smaller block of bone and matrix, and its crown is rotated distobucally from the main axis of remaining postcanine tooth row.
The horizontal shelf of the hard palate is absent (or covered by matrix) on the right side, but is represented by three pieces on the left side. These pieces are detached from the lingual alveolar segments. The palatal roof is rough and pitted. Alveolar depth to the palatal roof is approximately 13 mm as preserved at the LM2 position.
As a result of postmortem deformation, the left tooth row is shifted just slightly anterior and inferior relative to the right side, so that the left and right tooth rows are no longer in the same occlusal plane. The palate is narrower than it would have been prior to deformation. As preserved, the projected internal breath is approximately 34 mm between P4 and 41 mm between M1 or M2. Although postmortem deformation complicates observation of dental arcade shape, the right side is sufficiently intact to suggest the presence of a minor lingual concavity from P3–M2. Orientation of P3–M2 relative to the damaged subnasal area suggests that the tooth rows were slightly diverging, perhaps similar to the possibly contemporaneous Sangiran 4 palate.
Relatively complete maxillary dental rows of H. erectus are rare. We therefore describe the Sangiran 27 (S27) postcanine dentition in some detail, with reference to two other early Pleistocene dentitions: Sangiran 4 (S4) and Dmanisi 2282 (D2282) (Figure 4). The crown dimensions of the Sangiran 27 postcanine teeth are given in Table 1, together with those of selected comparative materials.
 View Details | Figure 4. Palates of Sangiran 27 (original with scale), Sangiran 4 (cast), and Dmanisi 2282 (original) showing details of upper dentition. Note incomplete distal marginal ridges in S4. |
Third premolar
The RP3 of S27 was rotated postmortem, resulting in an exaggerated distance between the distobuccal surface of P3 and the mesiobuccal edge of P4. The RP3 crown is damaged on its buccal crown portion, but is well preserved lingually. The P3 crown is in moderate occlusal wear (ASUDAS grade 1) and the occlusal wear bevels distolingually. There is a small dentine patch exposed on what remains of the buccal cusp. No lingual patch is evident. The wear direction of S27 differs from that of S4. The S4 P3 is more strongly worn towards the mesial margin of the tooth.
In occlusal view, the S27 RP3 is oval in outline with nearly parallel mesial and distal borders. The buccal and lingual cusps are nearly equal in buccolingual breadth, but the lingual crown is substantially shorter mesiodistally. The main mass of the lingual cusp is placed somewhat distal to the buccal cusp. The S4 P3 is similar in form, with slightly greater difference in mesiodistal length between its lingual and buccal cusps than the condition seen in S27. Although damage to the buccal cusp of S27 obscures observations, the buccal and lingual cusps appear to have been completely separated by a deep longitudinal fissure. In S4, this is also the case in the LP3; however, on the right side the two cusps are only incompletely separated by the longitudinal fissure. In S27, a small longitudinal ridge runs mesiodistally along this fissure. Mesially, the longitudinal fissure runs into an anterior fovea, where it deflects a short distance lingually. The buccal course, if any, of the mesial end of the longitudinal fissure is unobservable due to postmortem damage. Distally, the longitudinal fissure bifurcates into the posterior fovea. The lingual deflection is short; the buccal deflection is more substantial, passes bucally and then slightly mesially towards the buccal cusp tip.
The mesial marginal ridge is damaged but where preserved is thick lingually. The distal marginal ridge, which is thicker buccally than lingually, is thinner than the mesial marginal ridge and weakly incised by the longitudinal fissure. In contrast, in S4, the longitudinal groove of the LP3 has only slight buccal deflections at its mesial and distal ends. Additionally, the longitudinal fissure deeply incises the distal marginal ridge. Centrally, the longitudinal fissure is crossed by two short, transverse ridges that have the effect of forming a small central fovea. The longitudinal fissure of the RP3 is strongly distal to the transverse ridge. As in the LP3, it deeply incises the distal marginal ridge. Mesial to the transverse ridge, the longitudinal fissure forms a part of a fovea-like configuration, placed distal to the anterior fovea.
Fourth premolar
The S27 RP4 crown is missing the mesiobuccal quarter of its occlusal enamel but is otherwise intact. The buccolingual axis of the S27 P4 crown is rotated slightly diagonally. Like the RP3, the RP4 exhibits moderate wear (ASUDAS score 1), which bevels distolingually. A small dentine patch is exposed bucally and a tiny pinpoint exposure of dentine occurs lingually. The LP4 lacks most of its enamel except for the central portion of the lingual cusp and longitudinal fissure. The general occlusal outline and longitudinal fissure appear similar to the right side, but no other morphology is observable. Therefore, we describe the right side below.
The P4 of S27 is slightly shorter mesiodistally than the P3, as is the case in S4. This differential is greater in S4 than it is in S27. The S27 P4 is ovoid in occlusal view outline, with nearly parallel mesial and distal borders. This shape is the result of buccal and lingual cusps that are nearly equal in size, although the lingual cusp is both slightly narrower buccolingually and shorter mesiodistally than the buccal cusp. The S4 P4 shows a similar overall outline. However, the D2282 P4 has a lingual cusp that is mesiodistally longer than the buccal cusp.
In S27, the buccal and lingual cusps are separated by a short but deep (not wide) longitudinal fissure. Both mesially and distally, this fissure ends in discrete circular pits (the anterior and posterior fovea). Due to postmortem damage the anterior fovea cannot be observed further. The posterior fovea has an arm (fissure) that is deflected buccally. The mesial marginal ridge, where preserved lingually, is thick. The distal marginal ridge is complete, and is thinner than the mesial marginal ridge.
The D2282 P4 seems to have a similar morphology, with complete distal and mesial marginal ridges and a continuous longitudinal fissure; however, the longitudinal fissure only just moderately incises the transverse ridge joining the buccal and lingual cusps. In contrast, in the S4 P4, the longitudinal fissure is interrupted entirely by the centrally located transverse ridge. Distally, the longitudinal fissure crosses the distal marginal ridge; on the right side, a second, more buccally placed fissure also incises the distal marginal ridge. On the S4 LP4, two buccal fissures arise just distal to the mesial transverse ridge. They diverge from one another and define a central V-shaped portion of the buccal cusp. Anteriorly, the buccolingually oriented anterior fovea (or fissure) is completely separated from the longitudinal fissure and runs parallel to the complete mesial marginal ridge. The RP4 seems to retain similar features, although the greater degree of wear obscures the details.
First molar
The S27 RM1 lacks the middle third of the mesial marginal ridge and the adjacent portion of the occlusal surface between the paracone and protocone, but is otherwise intact. Occlusal wear is moderate to strong (ASUDAS grade 1), with a distolingual bevel. Dentine is exposed on all four main cusps with the protocone exhibiting the largest dentine patch. LM1 is similar to RM1 except it is slightly more worn. Because occlusal morphology is better observed on the right side, we describe the RM1 below.
The S27 M1 crown is rhomboidal in occlusal view outline and substantially broader buccolingually at its mesial than distal crown portions. The S4 M1 shows a similar crown outline, whereas in D2282, while the M1 is rhomboidal in outline, it is somewhat more mesiodistally elongated than it is in S27 or S4.
The paracone of S27 is separated from the other cusps by an L shaped groove with two accessory fissures. The shorter comes off the buccal limb and partially defines an accessory cusp between the paracone and metacone. The longer comes from the mesial limb. In the same region of S4, a full accessory cusp is formed between the metacone and paracone, which is further reduced in size by formation of a cusp between it and the protocone. In both S27 and S4, the buccal limb of the L-shaped groove is continuous with the buccal groove. In S27, the mesial limb extends to the preserved mesial break, but its course is unobservable from there. In S4, this limb is continuous with the longitudinal fissure. In neither specimen does an anterior fovea appear to be formed, whereas it is present in D2282. In S27, the buccal limb of the L groove is lingually continuous with a groove that extends to the protocone wear facet. As a result, the protocone joins the metacone by a crista obliqua located distal to this short groove. In S27, this small groove, the L-shaped groove defining the paracone, and the longitudinal fissure meet in a small central fovea. A similar configuration is seen on the LM1 of D2282, whereas the crista oblique is not incised in the LM1 of D2282 or in S4.
The hypocone of S27 is incompletely separated by an obliquely oriented, semilunar groove that continues to and includes the posterior fovea. The posterior fovea on the left side is more restricted buccolingually. As a result, the hypocone is continuous with the protocone mesially (i.e. the oblique groove is discontinuous with the lingual groove) and with the metacone buccally (via the distal marginal ridge). The distal marginal ridge is complete and thicker than the mesial marginal ridge. An apparent incision of the distal marginal ridge in the S27 M1 is a postmortem crack in the enamel. Similarly, D2282 also possesses an intact distal marginal ridge that is thicker than the mesial marginal ridge. In contrast, in S4, the oblique, semilunar fissure that separates the hypocone is lingually continuous with the lingual groove and distally incises the distal marginal ridge. A branch from the center of this groove divides off a 5th cusp (ASUDAS grade 5); another branch forms a small posterior fovea parallel to the distal marginal ridge. A similar cusp is developed in the D2282 M1.
On the buccal face of the S27 M1 a weak parastyle is defined by a vertical groove (ASUDAS grade 1) placed close to its mesial crown border. This is seen bilaterally. Neither S4 nor D2282 M1s have parastyles. However, the D2282 M1 shows evidence of a Carabelli’s complex on its lingual face, and at least the RM1 may show evidence of an enamel ridge that extends occlusally and distally.
Second molar
The RM2 of S27 is missing the mesiobuccal quarter of the occlusal rim enamel, but is otherwise complete. Occlusal wear is moderate and bevels distolingually. Dentine patches are preserved on the paracone, protocone, and a very small one on the metacone. LM2 is similar in morphology to its antimere, but due to postmortem damage lacks the central portion of its mesial marginal ridge. Its mesiobuccal paracone enamel has been restored, but has slipped slightly below the occlusal surface. Therefore, the description below is of the right side.
In occlusal view outline, the S27 M2 is rhomboidal in shape, and slightly more asymmetrical than M1 due to the relatively smaller size of the metacone. The asymmetry of this outline is typical of H. erectus (Spoor et al., 2007). The S27 M2 is absolutely larger than M1, as is the case in both D2282 and S4, but the size differential is not as great as in S4. As with M1, the D2282 M2 is somewhat more mesiodistally elongated than in the M2s of S27 and S4.
In S27, the paracone is separated by a semicircular to L-shaped groove arching buccal to mesial, and having multiple accessory fissures. The buccal and distal accessory fissure defines an accessory cusp between the paracone and metacone; the mesial and longitudinal accessory fissures define accessory cusps centrally and mesially. This is similar to the case in S4. The L-shaped groove forms part of the central fovea, it is not (apparently) continuous with the buccal groove, and its mesial course is obscured by postmortem damage. The buccal limb of the groove is deflected mesially at the central fovea. There is a groove that continues lingually distal to the protocone as the transverse groove, but this does not reach the lingual face of the tooth. Thus, the metacone and protocone are joined by a crista obliqua that is incompletely incised by this fissure. As in the M1, the S4 M2 has a continuous, but narrow crista obliqua, whereas the D2282 M2 has an incised crista obliqua.
In S27, the hypocone is partially separated from the protocone by a short, oblique groove that is convex mesiobuccally. The groove is discontinuous with the lingual groove and is buccally continuous with the posterior fovea. A similar situation describes both S4 and D2282 M2s. However, unlike the M1, the S4 M2 does not appear to possess a 5th cusp, whereas the D2282 M2 may possess one (wear renders this observation unclear). In S27, the distal marginal ridge is continuous and thicker than in the M1. Both S4 and D2282 also show continuous distal marginal ridges. In S27, the mesial marginal ridge is observable lingually where it is thinner than the distal marginal ridge but thicker than the mesial marginal ridge of M1.
On the buccal face, the parastyle region is not observable. The distal interproximal contact facet for M3 is located on the distal face of the hypocone (lingually) and is wider than it is tall. S27 shows no Carabelli’s complex development on the M2. S4 shows a broad Carabelli’s complex lingually (ASUDAS grade 6, but also extending distally across the lingual groove). In D2282, the Carabelli’s complex development, which is limited to the mesial half of the lingual crown face, is greater in the M2 than in the M1, and contributes to the asymmetrical occlusal view outline.
Third molar
The S27 RM3 morphology is not observable due to extensive postmortem crushing. The LM3 is located on the second block of matrix/bone. The tooth lacks the mesial third of the crown including paracone and protocone portions mesial to the cusp tips. Wear is moderate to light (ASUDAS grade 0–1) with dentine exposed on the protocone. Wear bevels lingually. The occlusal view outline is not determinable due to postmortem damage. All four major cusps are present. The size of the S27 M3 cannot be clearly compared to the other molars given the postmortem damage, but its buccolingual dimension can be estimated and does not seem as reduced relative to that in M2 as is the case for S4. In S4, RM3 is the smallest of the three molars and is compressed mesiodistally as the result of significant reduction of the metacone and hypocone. In S27, as the crown is incomplete, mesiodistal reduction of M3 cannot be assessed; however, it gives the impression that it is not as mesiodistally compressed as is S4.
In the S27 M3, a groove occurs distal to the paracone from the central fovea, and is continuous with the buccal groove. Lingually, the groove continues as the transverse fissure located distal to the protocone. An accessory fissure mesial to the transverse fissure comes from the central fovea and extends towards the protocone tip. A mesial and buccal fissure extends from the central fovea towards the paracone. Another fissure extends distally from the central fovea toward the crista obliqua but does not entirely incise this crest. The metacone and protocone are joined by a narrow crista obliqua that is incompletely traversed by the fissure described above. In contrast, in the S4 M3, the crista oblique is not continuous.
The hypocone of S27 is incompletely separated by a mesially convex semilunar oblique fissure that widens distally into the posterior fovea. Neither the lingual nor buccal ends of the groove extend to the edge of the tooth. In the S27 M3, the distal marginal ridge is incised by a groove from the posterior fovea, whereas in S4 the distal marginal ridge is intact. In S27, a second smaller groove, lingual to that which incises the distal marginal ridge, incompletely defines a fifth cusp (ASUDAS grade 1). No fifth cusp is defined at the distal crown of the S4 M3, although a mesial and central cusp is defined between the paracone and protocone.
Craniofacial comparisons
Morphologically, the preserved Sangiran 27 face and vault compares favorably with both the relatively sparse early Indonesian record and with later Indonesian H. erectus. Overall vault shape cannot be assessed due to postmortem deformation. However, postorbital constriction was probably moderate to pronounced in Sangiran 27, similar to the Sangiran 17 condition. The supraorbital torus of Sangiran 27 is massive in vertical height (Table 1), apparently evenly thick from the central to the lateral portions of the orbit, and probably barlike. However, preservation is not sufficient to enable a description of glabellar region shape. Given its large size and development, we found no evidence of the apparent ‘feminine’ features of the torus ascribed by Wolpoff (1999: p. 457).
The posterior cranium cannot be assessed in Sangiran 27, but it is massive in the other early specimens Sangiran 4 and 31 (Antón and Indriati, 2002). Prognathism cannot be accurately assessed, but it was probably weak, similar to that seen in Sangiran 4. The infraorbital region is convex transversely, cheek height is relatively tall, and the nasal sill of Sangiran 27 is smooth. These facial features make Sangiran 27 similar in overall morphology to the later Sangiran 17 condition.
Sangiran 27 contrasts with the broadly contemporaneous specimens from Dmanisi, Georgia, and Turkana region, Kenya, spanning the 1.5–1.8 Ma time period, in much the same way that the other Indonesian H. erectus differ from early African H. erectus. Sangiran 27 is larger and more robust than specimens from Dmanisi and East and West Turkana in its greater development of the supraorbital torus and thicker cranial vault bones (Table 1, Table 2). In addition, unlike D2282 and KNM-WT 15000, Sangiran 27 lacks paranasal pillars. The presence of such pillars is probably a primitive condition.
Thus, little in the craniofacial anatomy of Sangiran 27 would support designation as H. habilis or H. rudolfensis rather than as H. erectus. Sangiran 27 shares with the other probable earliest Pleistocene Indonesian specimens a hyper-robust appearance; that is, they exhibit thicker vaults, larger superstructures, and are generally more massive than their Georgian or African conspecifics (see, for example, the size analysis of Antón et al., 2007 and descriptions in Rightmire et al., 2006). Based on the cranial capacity of Sangiran 4 (908 cm3), the earliest Indonesian crania may be somewhat larger in overall size compared with their Georgian and African contemporaries (see Antón, 2002, 2003).
Dental comparisons
Dental anatomy and size show strong similarities between Sangiran 27 and its probable contemporary Sangiran 4. Occlusal area is absolutely large in both fossils. The molars possess high mesial marginal ridges and accessory cusp complexes, as do the isolated teeth of Sangiran 7, presumably from the Sangiran Formation (Grine and Franzen, 1994). However, distal marginal ridge shape is variable, tending to be incomplete in Sangiran 4 (Antón and Indriati, 2002). The upper canine was probably large in Sangiran 27, as it was in Sangiran 4 (Wolpoff, 1999: p. 456; Antón and Indriati, 2002), although the mesial slope of the P3 wear would appear to indicate that the lower canines were larger in S4 than in S27.
The Sangiran 27 dentition, like the other early Indonesian postcanine dentitions (Kaifu et al., 2005; Kaifu, 2006), exhibits large occlusal areas, and differs in molar crown shape from the buccolingually narrow crowns of early African H. erectus (Table 1). Occlusal areas of the Indonesian molars are more similar to those of the adult dentitions of Dmanisi (Gabunia et al., 2000), although the Dmanisi molars are more elongated mesiodisatally as in the early African examples of H. erectus. The Sangiran 27 and Dmanisi molars are distinguished from the even larger H. rudolfensis molars; they are accompanied by development of accessory cusp complexes, whereas this is not the case in H. rudolfensis or H. habilis. The Sangiran 27 and Dmanisi dentitions also share a more bulbous cusp profile that again is partly the result of the presence of accessory cusp complexes. In particular, Sangiran 4 and Dmanisi 2282 share the presence of a 5th cusp and Carabelli’s complex development, while Sangiran 27 and Dmanisi 2282 differ in mesial marginal ridge height, which is high in Sangiran 27 (and Sangiran 4), but low in D2282 and early African H. erectus of the Turkana region (Antón and Indriati, 2002).
Although it exhibits significant postmortem distortion, Sangiran 27 suggests that the earliest Indonesian hominins had morphological features typical of later Indonesian H. erectus and, if anything, are hyper-robust compared with their successors (see also Kaifu et al., 2005). The anatomy of the earliest Pleistocene Indonesian fossils further suggests that, from its inception, the Indonesian hominin lineage was derived relative to the time-equivalent counterparts of Africa, particularly as relates to the dentition.
We are grateful to our many colleagues worldwide for access to specimens and intriguing discussion, particularly Dr. Y. Kaifu. The following individuals allowed access to specimens in their care: Dr. F. Aziz, Quaternary Research Laboratory, Geological Research and Development Center, Bandung, Indonesia; Dr. J.L. Franzen, Senckenberg Museum, Frankfurt; Dr. D. Lordkipanidze, Georgian State Museum, Tbilisi, Georgia; Dr. I. Tattersall and K. Mowbray, American Museum of Natural History, New York. This study was partially supported by NSF BCS-9804861.
|