| Correspondence to: José Miguel Carretero, Laboratorio de Evolución Humana, Departamento Ciencias Históricas y Geografía, Universidad de Burgos, Edificio I + D + i, Plaza de Misael Bañuelos s/n, Burgos 09001, Spain. E-mail: jmcarre@ubu.es Published online 1 August 2008 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.070413 |
The Bodo partial distal humerus (BOD–VP–1/2), one of the very few postcranial remains from the African Middle Pleistocene, was recovered in 1990 from the surface of the upper Bodo Sand Unit (Clark et al., 1994). It was found approximately 100 m north of the original Bodo cranium discovery at approximately the same stratigraphic horizon. Ar/Ar ages, biostratigraphy, and tephrochronology indicate that the Pleistocene Bodo hominids are approximately 0.6 million years old (Renne, 2000). The age, stratigraphy, and archaeological assemblages of the site are discussed in Clark et al. (1994) and de Heinzelin et al. (2000).
We present here a brief report based on work on the original specimen housed in the Paleoanthropology Laboratory at the National Museum of Ethiopia, Addis Ababa. Despite its fragmentary status, several relevant variables can be directly measured on the specimen or can be reasonably estimated. Our comparisons of the Bodo humerus with African and European fossil hominid and modern human samples reveal some insights into the variation and affinities of the African hominids of the Middle Pleistocene.
We compared the Bodo distal humerus with the available humeral specimens of Homo from the Early, Middle, and Late Pleistocene of Africa and Europe. The Asian fossil evidence for this anatomical region (distal humerus) is limited to the recently published H. georgicus postcranial elements from Dmanisi, Georgia (Lordkipanidze et al., 2007). Nevertheless, the variables we use in this analysis are not available for the Dmanisi humeri, and our comparisons are based on the specimens and samples listed in Table 1.
Among the fossil samples in our comparisons we included some original specimens from the Sima de los Huesos Middle Pleistocene site, in the Sierra de Atapuerca (Burgos, Spain), with a very similar chronology to the Bodo hominids. New high–precision dates on the recently discovered speleothem overlaying the human bones within the Sima de los Huesos (SH) yielded finite dates ranging from 563 to 668 ka (average 600 ka) (Bischoff et al., 2007). A conservative approach by the authors (Bischoff et al., 2007) was to consider the lower limit ages as the minimum age of the speleothem, a range from 513 to 571 ka (average 530 ka). This minimum age is consistent with the faunal analysis of the site. The SH rodent fauna suggest that the site would correspond to about Marine Isotope Stage (MIS) 15/16 (500–600 ka). Among the larger fauna, the SH population of Ursus deningeri and Panthera leo fossilis resembles more closely the specimens from Mauer, Mosbach, or Arago (MIS 12–16) which approximately date to between 450 and 600 ka based on faunal and radiometric criteria (García et al., 1997; García, 2003). This compatibility between the new age estimates determined for the speleothem and the biostratigraphic indicators from the site provides strong evidence that the radiometric results should be considered conclusive for a minimum age of 530 ka (MIS 14) for the SH sediments (Bischoff et al., 2007).
The Bodo humeral fragment comprises approximately the distal third of the diaphysis (Figure 1). From a descriptive point of view, humeral length can be divided into percentages from distal (0%) to proximal (100%) following Trinkaus et al. (1994). By direct anatomical comparison with complete humeri, the Bodo specimen approximately represents the segment between the 35% level proximally (mid–distal shaft) and midway between the 10% and 5% levels distally (mid–olecranon fossa) (Figure 2).
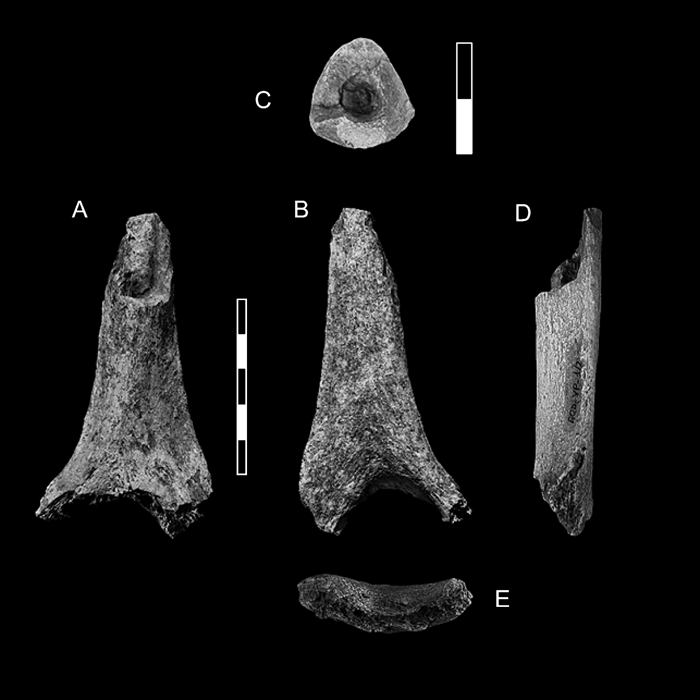 View Details | Figure 1. Anterior (A), posterior (B), superior (C), lateral (D) and distal (E) views of the BOD–VP–1/2 humerus. Anterior and posterior views taken on the original. Superior, lateral and distal views taken on cast. For the superior view (cross–section), anterior is up and lateral is to the left. Scales in cm. |
 View Details | Figure 2. (I) Sketch of the rough position of Bodo specimen in a silhouette of a complete humerus divided in percentages of length from distal to proximal following Trinkaus et al. (1994). (II) Sketch of Bodo humerus showing the position of the most proximally preserved point (1) and proximal section level (2) were the cross–section was studied. (III) Sketch of Bodo humerus showing the variables taken in the distal region: LPT, lateral pillar thickness; MPT, medial pillar thickness; OFB, olecranon fossa breatdth. |
The maximum preserved length of BOD–VP–1/2 is 94.0 mm and its maximum preserved breadth at the distal break is 51.0 mm. Although the distal articular surface is missing, the proximal half of the olecranon fossa and the medial and lateral pillars adjacent to the fossa are well preserved. Therefore, the olecranon fossa breadth and the thicknesses of the medial and lateral pillars can be measured with precision (Figure 2).
The proximal fracture of the diaphysis is irregular in shape along the first 25 mm where only the posterior wall and small portion of the medial wall of the shaft are preserved (Figure 1). No measurement can be taken along this short segment. Distal to this, the section of the diaphysis is complete and can be measured accurately. We have named this second position the proximal section level, to distinguish this position from the most proximal preserved point, 25 mm above (Figure 2). The cross–sectional shape at the proximal section level is subtriangular (Figure 1), the shaft’s anteroposterior diameter is 19.6 mm, the mediolateral diameter is 21.0 mm, and the perimeter is 65 mm. The shaft index is 93.3%.
For our comparative analysis, it is important to exactly locate the position of the proximal section level of BOD–VP–1/2 following the terminology of Trinkaus et al. (1994), and more specifically to determine the minimum shaft perimeter (MSP) of the specimen. Anatomically, the proximal section level is for certain included between the 25% and 35% levels. In our experience with measuring complete humeri, the MSP is always included between the 30% and 40% levels, and in the majority of specimens the MSP position coincides with the 35% level. Nevertheless, along this segment, and in general along the distal shaft, the diameters and perimeter vary very little given the absence of significant muscular markings. Normally, an evenly tapering diaphysis is seen distal to the deltoid tuberosity. For example, it was assumed that the 43% values for areas and second moments of area are the average of the 35% and 50% values (Trinkaus and Churchill, 1999).
Following these morphological observations, we can consider the perimeter of the Bodo proximal fracture level (65 mm) as a maximum value for the MSP, since the real value would be equal or slightly smaller, and would be located somewhat higher in the shaft. The lack of shaft variation between the 25% and 40% levels in normal modern humeri suggests that in no case would the real MSP of the Bodo specimen be much less than 65 mm, perhaps 64 or 63 mm at the least. Therefore, in our analysis, we adopted this range of variation (63–65 mm) for the MSP of the Bodo humerus.
Another important variable for our comparative purposes is the biepicondylar breadth (BB). As the distal end is missing, we estimated the BB breadth of BOD–VP–1/2 based on preserved shaft dimensions, specifically on the MSP taken at the proximal section level (63–65 mm). Humeral shaft dimensions are to some degree developmentally and environmentally plastic, so they may not be the best dimensions to estimate distal epiphyseal size or to standardize for overall bone size. Nevertheless, in the Bodo specimen, the shaft is the only preserved region. Due to the importance of this estimation for our comparative purposes, linear regression equations of BB (dependent variable) on MSP (independent variable) were computed using two chronologically different recent human samples of balanced sex: the San Pablo Monastery and the Coimbra modern human collections (Table 1). Furthermore, we computed a linear regression from the seven distal humeri of SH (Table 2).
Based on the three different regression equations, we estimated the BB of BOD–VP–1/2 using our accepted minimum (63 mm) and maximum (65 mm) values of MSP (Table 3). The two modern human–based regression lines produced close results for both shaft perimeter estimates (Table 3). The average of all predicted values was 60.2 mm and the range of 95% confidence limits for all estimations was 58.2–62.4 mm. On the other hand, the SH regression line predicted 61.9 and 63.0 mm for the minimum and maximum MSP values, respectively (mean 62.5 mm), as well as wider confidence limits that taken together range between 59.6 and 65.6 mm (Table 3). The average of the modern (60.2) and SH (62.5) predicted means was 61.4 mm.
To further evaluate to some degree the validity of the two modern human–based equations, we estimated the known BBs of seven humeri from the SH collection that include male and female specimens. As seen by the residuals in Table 4, the two modern linear regressions underestimated virtually all the SH specimens, and the mean residual of both was 2.6 mm (1.6 mm for San Pablo and 3.6 mm for Coimbra). Therefore, compared with the Coimbra and San Pablo specimens, the seven distal humeri from SH have a larger BB than expected for a modern human with the same mid–distal shaft perimeters. The question arises as to whether, by using the Coimbra and San Pablo regression lines, we are also underestimating the BB of the Bodo specimen (2.6 mm in average), and whether the predicted values based on the fossil sample (SH regression line) are better than the predictions based on recent European samples. The Bodo hominids could be much closer in body size and skeletal robusticity to other Middle Pleistocene humans than to recent Europeans (Grine et al., 1995; Arsuaga et al., 1999; Pearson, 2000), as is the case, for example, with the Bodo cranium, which exceeds modern humans in facial size and vault robusticity. If we accept this possibility, probably 62 or 63 mm rather than 60 mm is a better estimate for the BB of the Bodo humerus.
Finally, we made comparisons of overall size and shape by superimposing the Bodo humerus outline with that of the SH specimens. This indicates that the shaft dimensions at the proximal section level, the olecranon fossa breadth, and the medial and lateral pillar thicknesses of the Bodo humerus are slightly larger than in H–VI and H–XV (BB is 61 and 61.1 mm, respectively) and slightly smaller than in H–III, H–II and H–X (BB is 66, 65 and 63 mm, respectively) (Figure 3). The mean BB of these five SH specimens is 63.3 mm with a range of 61–66 mm (Table 4). We excluded H–XI and H–XIV from the SH mean calculation because they are clearly smaller than Bodo. Here, we have to remember that distally, the maximum preserved breadth of the Bodo specimen is 51 mm, and in no case is the BB of the specimen going to be lower than 60 mm (virtually the mean predicted value with the two modern regression lines).
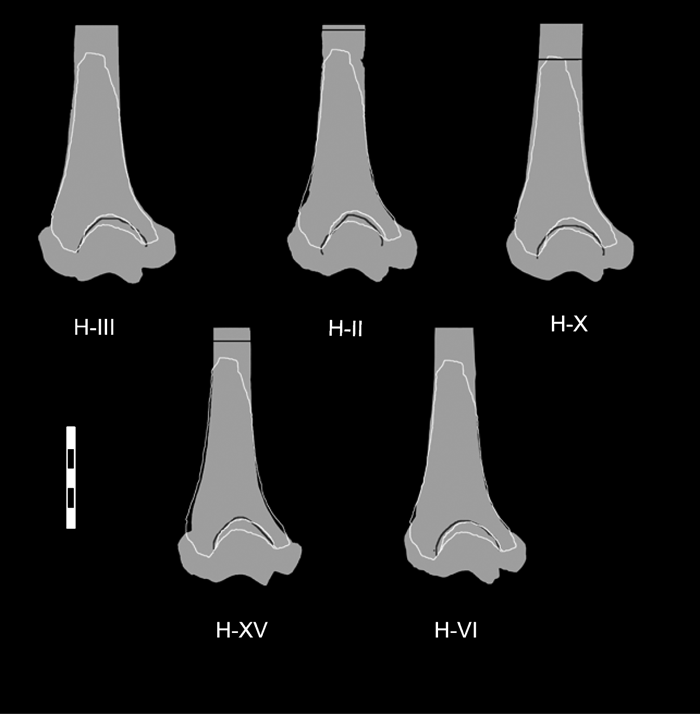 View Details | Figure 3. Outline of the Bodo humerus silhouetted over the humeri from Sima de los Huesos (SH) to compare its general dimensions. Dark line indicates the 35% level in the three SH complete specimens. Biepicondylar breadths of the SH specimens are as given in Table 4. Scale in cm. |
In conclusion, due to the difficulty in adopting a single value for the BB of Bodo, and taking into account the possible variation in the MSP of the specimen, as well as the results of the three regression lines and the metric comparisons with the SH humeri, a range of 60–66 mm for the BB of the Bodo humerus is considered reasonable. We strongly suspect that the BB of BOD–VP–1/2 was probably between 62 and 63 mm. Nevertheless, to be cautious, we adopted the broader range of 60–66 mm in our comparative analysis.
Finally, to aid in the evaluation of the sex of the fossil specimen, we compared the MSP and estimated BB of the Bodo humerus with large European and American samples of known sex—most of them measured by us (Table 5). Both the MSP and the accepted range of BB are clearly coincident with the male and well above the female values (Table 5). This sexual diagnosis is also suggested by the cross–sectional geometrical parameters, all of them well within the male ranges (see below). Therefore, based on our comparative analysis, BOD–VP–1/2 is more likely attributed to a male than to a female individual. Of course, we do not know the humeral size distribution as it relates to sex in a 600,000–year–old human population in the Horn of Africa, so these results must be interpreted carefully. This is because, as we noted above, some contemporary individuals presumably from the same population, such as the Bodo cranium which is unassociated with postcranial elements, exceed modern humans in facial size and vault robusticity.
Few standard measurements can be measured on this specimen. Apart from the estimated BB, three more variables can be directly measured in the distal region, namely olecranon fossa breadth and medial and lateral pillar thicknesses (Figure 2, Table 6, Table 7). These three variables are of some evolutionary interest. In the distal humeral samples of SH (H. heidelbergensis) and the Neandertals (H. neanderthalensis), a high frequency of a relatively wide olecranon fossa and thin lateral and medial pillars adjacent to the fossa were found (Carretero et al., 1997). In our observation, although these morphological characters are also found in modern human humeri, in all the samples that we have studied they appear in a significantly low frequency (Table 6).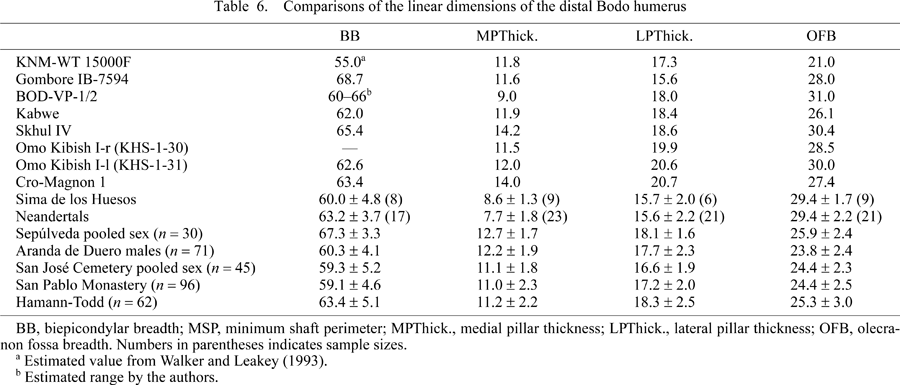

With caution, these two traits can be examined in the Bodo humeral fragment. Using our estimated range for the BB of the Bodo humerus (60–66 mm), the minimum and maximum olecranon fossa index values of this specimen are high (47.0–51.0). This range is very similar to the SH and Neandertal sample means (Table 7). At the same time, compared to our modern human samples, the medial pillar is narrow both as a proportion of the olecranon fossa breadth (Figure 4) and relative to the estimated BB (Figure 5, Table 7). Therefore, in these features, the Bodo distal humerus is more similar to the SH hominids and the Neandertals than to our modern human samples. On the contrary, the Kabwe distal humerus displays the same olecranon fossa and pillar proportions that characterize the majority of modern humans (Carretero et al., 1997) (Table 7). The KNM–WT 15000F West Turkana juvenile humerus displays a small olecranon fossa relative to BB, as well as modern human–like medial and lateral pillar proportions. Finally, the distal humerus from Melka Kunture, Gombore IB, attributed to Homo (Chavallion et al., 1977; Senut, 1981) also displays modern human proportions in these two traits.
 View Details | Figure 4. Medial pillar thickness vs. olecranon fossa breadth in several fossil specimens. The 90% equiprobability ellipse for a pooled modern human sample that includes the Aranda, San Pablo, and Hamann–Todd individuals (n = 275) is outlined. Dark triangles, Neanderthal; dark squares, Sima de los Huesos specimens. |
 View Details | Figure 5. Medial pillar index (medial pillar thickness/biepicondylar breadth) × 100 calculated for the fossil humeri, two human fossil samples, and five recent human samples. Neanderthal sample as in Carretero et al. (1997; Table 18, p. 400). SH, Sima de los Huesos; SJ, San José; SP, San Pablo; HTH, Hamann–Todd, Numbers in parentheses indicate sample sizes. Vertical bars for recent human samples represent 1.5 standard deviations around the sample mean. Vertical bar for Bodo indicates the maximum, mid–range and minimum values of the pillar index for biepicondylar breadths of 60, 63, and 66 mm, respectively. |
As we are working with trait frequencies in Middle Pleistocene populations, the significance of the morphology of an isolated African specimen is not obvious relative to the European samples. Three out of the four African Homo distal humeri (KNM–WT 15000, Gomboré IB, and Kabwe) ranging in age from 1.5 to 0.2 Ma (Table 1) display humeral distal proportions more common in modern humans, while only Bodo shows the pattern more common in the European Middle Pleistocene hominids and the Neandertals.
Besides the AP and ML shaft diameters and the perimeter (see above), cross–sectional parameters can also be studied at the proximal section level (Table 8, Table 9). Generalized thickening of cortical bone is a well–known cranial and postcranial characteristic of all representatives of Pleistocene Homo, except H. sapiens, in which this trait is not reflected on the entire skeleton. The great cortical thickness of the Bodo humerus was noted by Clark et al. (1994) as one of its remarkable characteristics. As stated above, the proximal section level of BOD–VP–1/2 approximately corresponds with the 35% position (or somewhat below this) from the distal end of a complete humerus, so we can compare the cortical area of the cross–section at this level. The Bodo humerus was found to be very similar to the SH and Neandertal humeri in its cortical bone proportion, with a cortical area that is 80% of total cross–sectional area. This value is well above the means of some recent human samples studied by Trinkaus et al. (1994) (Table 8). The relatively thick cortical bone demonstrates the greater resistance of all these archaic humeri to axial loading in comparison with some recent humans (Trinkaus et al., 1994, 1999; Trinkaus and Churchill, 1999). According to Trinkaus et al. (1999), at both the 35% and 65% levels, Neandertals generally have a higher relative cortical area than that of early modern humans.
Other cross–sectional geometric properties measure the distribution of skeletal tissue in bone sections that are related to biomechanically relevant measurements, and reflect loading modes, structural adaptation, and activity patterns (e.g. Larsen, 1997; Ruff, 2000). At this point, it is necessary to remember that the Bodo humerus is from the left arm, which tends to be the non–dominant limb. Given the plasticity of long bone diaphyses and the high degree of humeral bilateral asymmetry in Pleistocene humans (Trinkaus et al., 1994; Churchill and Formicola, 1997), a considerably greater level of variation in cross–sectional measures exists in the left humerus, and most individuals exhibit greater cross–sectional properties on the right side (Trinkaus and Churchill, 1999).
The polar moment of area (J) was examined as an overall measure of diaphyseal rigidity, since it approximates torsional rigidity and sums the perpendicular measures of bending rigidity (Trinkaus et al., 1999). In order to compare the biomechanical properties of long bones of different individuals or samples, it is necessary to incorporate some kind of size standardization. This is usually done by dividing cross–sectional properties by powers of bone length (Ruff, 2000), a variable that is not available for the Bodo specimen. Although we could attempt to estimate the maximum and biomechanical lengths of the Bodo humerus, based on the already estimated BB, a simple and less speculative procedure for at least partially controlling for differences in body size (not limb length) is to compare cross–sectional diaphyseal properties relative to articular size (Ruff, 2000), in our case with the estimated BB (60–66 mm). We remember again that Bodo humerus is from the left side, hence may reflect base–line rather than dominant activity levels.
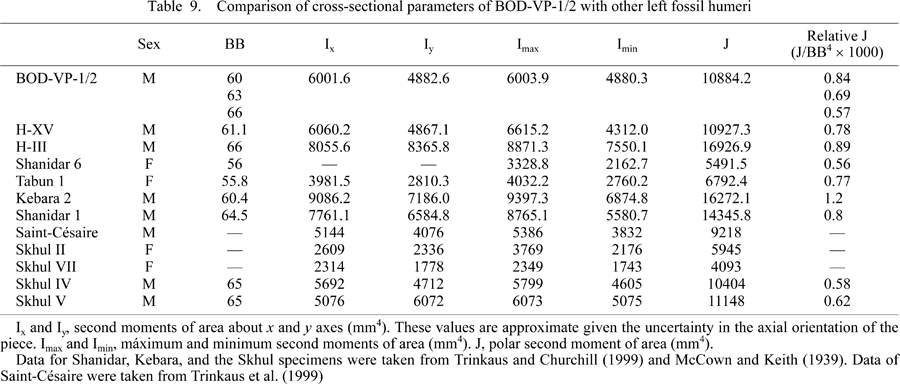
In absolute values, the polar moment of area (J) and the rest of the cross–sectional parameters of the Bodo humerus at the proximal section level are very similar to those of the SH H–XV, Saint–Césaire Neandertal, and the Skhul IV and V early modern humans. In the Bodo humerus, the J value is clearly larger than in Tabún 1, Shanidar 6, and Skhul II females, giving support to the sexual diagnosis mentioned above, but it is much lower than in Kebara 2, SH H–III, and Shanidar 1 males (Table 9, Figure 6a).
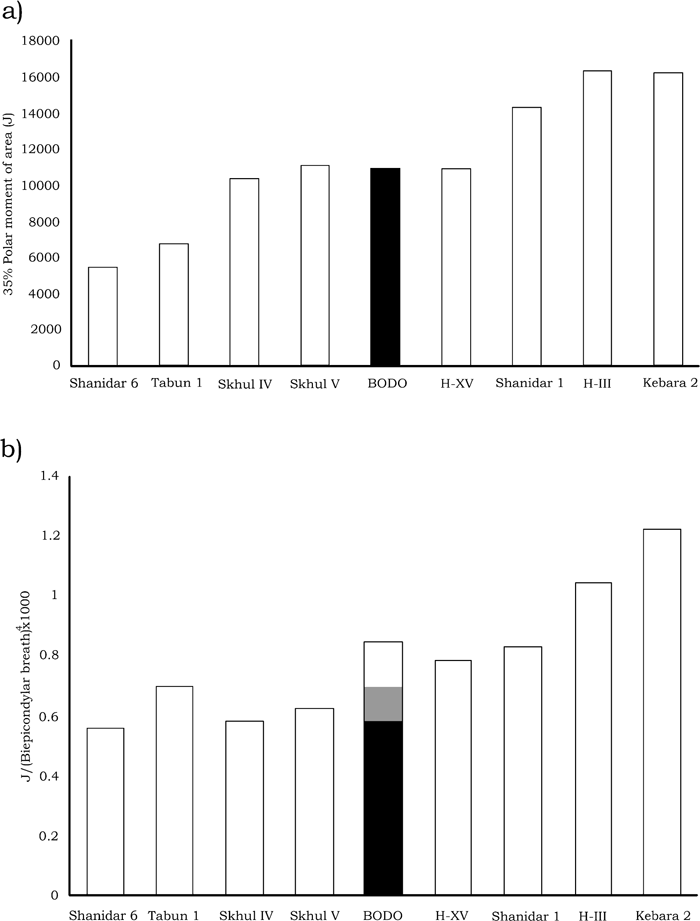 View Details | Figure 6. (a) Absolute polar moment of area for Bodo and other left fossil humeri at the 35% level calculated as in Table 9. (b) Relative polar moment of area of the same specimens. Vertical bar for Bodo indicates the maximum, mid–range, and minimum values of the index for biepicondylar breadths of 60, 63, and 66 mm, respectively. |
Relative to the BB used as a proxy of body size (Table 9, Figure 6b), the Bodo humerus does not show a very robust diaphyseal rigidity either, if we assume the maximum BB value of the proposed range (66 mm). Using the mid–range value of the BB of 63 mm, Bodo has the least robust section of the four archaic males in the sample (Table 9). In this comparison, Bodo was similar to the Tabun 1 female but far from the robustness exhibited by Shanidar 1, SH H–III, and Kebara 2 males. Interestingly, in this case the shaft rigidity of the Bodo humerus is above that of the Skhul IV and V modern human males. Using a BB value of 60 mm, the shaft rigidity of the Bodo humerus reaches the values exhibited by other archaic male left humeri such as those of Shanidar 1 and SH H–XV (Figure 6b).
The taxonomy and evolutionary relationships of Middle Pleistocene African and European populations are currently a matter of debate (Smith et al., 1989; Arsuaga et al., 1997; Hublin, 1998, 2000; Rightmire, 1998; Stringer and Hublin, 1999; Tattersall, 2000; Hawks and Wolpoff, 2001; Bermúdez de Castro et al., 2004). Whatever the taxonomic and phylogenetic frameworks adopted, comparison of the Bodo humerus with the European samples reveals some insights into the African fossil hominids of the Middle Pleistocene.
In our view, H. heidelbergensis is an exclusively European species and is ancestral only to the Neandertals (Arsuaga et al., 1991, 1993, 1995, 1997; Rosas and Bermúdez de Castro, 1998). For the African Middle Pleistocene specimens, the specific name H. rhodesiensis is preferred and is generally held to represent the ancestral lineage that eventually gave rise to modern humans. Finally, the recent definition of the species H. antecessor in the European terminal Early Pleistocene (c. 800 ka) (Carbonell et al., 1995; Bermúdez de Castro et al., 1997) raises the question of its evolutionary relationship with the near contemporaneous African specimens (Carbonell et al., 2005), including the Bodo remains.
As mentioned previously, a relatively wide olecranon fossa and the relatively thin medial and lateral pillars adjacent to the fossa (or the high frequency of this pattern) represent a distinctive or typical morphological pattern of the Middle and Late Pleistocene European Neandertal lineage (Carretero et al., 1997). The admittedly scarce African evidence, including the new Bodo specimen, highlights the morphological variation of these traits. Nevertheless, as a group, the modern human pattern of a narrower olecranon fossa and relatively thicker medial and lateral pillars is the most common condition of the available Early and Middle Pleistocene African distal humeri.
Some Neandertal postcranial traits are primitive features within the genus Homo, but other aspects appear to be derived, or specialized, rather than merely being primitive retentions (Trinkaus, 1977, 1983, 1995; Ruff, 1994; Villemeur, 1994; Carretero et al., 1997; Holliday, 1997; Arsuaga et al., 1999). Of these derived Neandertal features in the 500–600 ka fossils of H. heidelbergensis from the Sima de los Huesos in Atapuerca, some are present in a mosaic fashion and in lower frequencies (Carretero et al., 1997, 2005). What we find in Europe during the Middle Pleistocene are morphological traits that are variable within the H. heidelbegensis populations, until the final fixation (very high frequency) of one morphotype in the Late Pleistocene Neandertals that was well established by 200 ka. This model has been formally named the ‘accretion model of Neandertal evolution’ (Arsuaga et al., 1997; Martínez and Arsuaga, 1997; Dean et al., 1998; Hublin, 1998, 2000; Stringer and Hublin, 1999).
Although the Neandertals show numerous derived characters in their postcranial skeleton, this is not the case for early modern humans. Unfortunately, the Early and Middle Pleistocene African fossil record is characterized by a paucity of postcranial specimens and this complicates our understanding of when and how the modern human morphology emerged.
The postcranial morphology of H. ergaster is known from the remains of KNM–WT 15000 (Walker and Leakey, 1993) and numerous fragmentary postcranial remains from Koobi Fora and Olduvai (Day, 1986). Morphological diversity is present in H. ergaster in some features that are very frequent and phylogenetically relevant in Neandertals. For example, the morphology of the scapula is well known in Neandertals (Trinkaus, 1977, 1995) and the SH hominids (Carretero et al., 1997), and in both species there is a high frequency of an axillary sulcus dorsally placed along the lateral margin. Early anatomically modern humans (the Skhul–Qafzeh sample), Upper Palaeolithic people, and modern populations tend to have either a ventrally positioned sulcus (ventral sulcus) or an intermediate morphology (bisulcate pattern) (McCown and Keith, 1939; Trinkaus, 1977; Vandermeersch, 1981). The KNM–WT 15000 partial skeleton exhibits a ventral axillary sulcus on the scapulae (Walker and Leakey, 1993), but the specimen KNM–ER 1808 displays a clear axillary sulcus in a dorsal position (Trinkaus, 1995; Carretero et al., 1997 and personal observation). At least with regard to this postcranial trait H. ergaster seems to be very variable.
In Africa, the few human postcranial fossils postdating H. ergaster from the period between roughly 800 and 300 ka (early Middle Pleistocene) also exhibit a surprising amount of morphological diversity, which includes specimens that do not differ substantially from modern humans, and other bones that are distinctly not modern (Pycraft et al., 1928; Stringer, 1986; Solan and Day, 1992; Trinkaus, 1993; Grine et al., 1995; Churchill et al., 1996; Bräuer et al., 1997; Stringer et al., 1998; McBrearty et al., 1999; Pearson, 1999, 2000; Churchill et al., 2000; Yokley and Churchill, 2006). Morphological variation is also present in specimens dating to the late Middle Pleistocene (c. 200–120 ka) such as the radius of the Cave of Hearths, which display a radial tuberosity oriented anteriomedially as in modern humans as well as a long radial neck and small radial head as in the Neandertals (Tobias, 1971; Pearson and Grine, 1997).
Human postcranial fossils dating from the African Late Pleistocene also show that the earliest modern humans from Africa and the Levant displayed a considerable amount of morphological heterogeneity, and that the postcranial skeletons of these hominids differed from those of recent humans as well as from European Neandertals (McCown and Keith, 1939; Day, 1969; Vandermeersch, 1981; Day et al., 1991; Churchill et al., 1996; Pearson and Grine, 1996, 1997; Grine et al., 1998; Pearson et al., 1998; Pearson, 1999, 2000). For example, modern human pelvic shape constitutes a genuine novelty of our species (Arsuaga et al., 1999; Carretero et al., 2004), but some of the earliest modern humans, such as Skhul IX, retain plesiomorphic details of pelvic anatomy such as a relatively long pubic ramus (Rak and Arensburg, 1987). This is a primitive trait (Arsuaga and Carretero, 1994; Rosenberg, 1998; Arsuaga et al., 1999) retained in some of the Skhul–Qafzeh specimens, but subsequently lost in the modern human lineage (Pearson, 1999, 2000).
It seems that the anatomically modern form as represented by living humans was not entirely complete in the Skhul–Qafzeh hominids; many minor features had yet to change to their modern frequencies. Furthermore, different individuals from these African and Levantine samples often display different sets of features that distinguish them from studied collections of recent humans. A parallel situation is seen between European Middle Pleistocene humans and Neandertals. Furthermore, and perhaps more importantly, almost all of their distinctive features, when considered singly, can be found in one or more groups of recent humans, but the entire suite of distinctive features cannot be found in any single group of modern humans.
Regarding the cranial evidence, the Bodo cranium, for example, has an extraordinarily broad face but its morphology recalls that of the European Middle Pleistocene fossils such as Arago 21 and Petralona, although in the latter two specimens there could be some Neandertal traits (such as the raised and sharp lower rim of the nasal margin of Arago 21). The Sima de los Huesos site has an age which is probably similar to that of the Arago human sample (that of the Petralona skull is unknown) but it is more Neandertal–derived in the midfacial prognathism (but not in the lower nasal margin). The Bodo skull does not show any Neandertal character, and in fact could represent the primitive morphotype of the European and African Middle Pleistocene populations, just before the split between the European and African lineages. There are however some possible instances of polymorphism in African Middle Pleistocene specimens. In addition to the facial contrasts between Bodo and Broken Hill, the mastoid region of Laetoli hominid 18 is reminiscent of Neanderthals (Arsuaga et al., 1997). In his study of the Petralona skull, Stringer (1983) also stated that supposed Neandertal apomorphies do occur sporadically in specimens which are clearly not otherwise Neandertal (e.g. Laetoli hominid 18 has a large occipitomastoid crest; Broken Hill has a Neandertal–like forward position of the first upper molar alveolus).
The presence of Neandertal traits in some, but not all, specimens of H. ergaster, H. Rhodesiensis, and early H. sapiens, together with the greater similarity of only some African specimens, but not others, to modern humans highlights the problem of anatomical variation in African hominids of the early Middle Pleistocene. At least two possible explanations for this variation can be proposed (Pearson, 2000). Some of the more modern–like postcranial fossils may represent populations that were, in fact, either ancestral to modern humans or more closely related to our ancestors than the populations represented by the specimens such as Berg Aukas. Alternatively, populations of early Middle Pleistocene African humans may have simply been physically diverse and their skeletons may have contained mosaics of modern and non–modern traits (Pearson, 2000).
The distal humerus from Bodo points out, once again, the presence of variability in the postcranial skeleton of African Middle Pleistocene hominids. We think that an analysis of frequency–based patterning can help us interpret this variation. Taking into account the within–species (sample) variation we see in the fossil record (see above), the explanation that there was polymorphism in several skeletal traits in the Middle Pleistocene of Europe and Africa is quite plausible. Some specimens on both continents would show the Neandertal condition. Through time this morphology became fixed in the classic European Neandertals but lost in Africa. We draw this conclusion with the support of corroborative observations. This is exactly the case in the Middle Pleistocene hominids from Europe in cranial (Arsuaga et al., 1997; Martínez and Arsuaga, 1997) and postcranial morphology (Carretero et al., 1997, 2005).
In the present work the Bodo partial distal humerus from Ethiopia with an approximate age of 0.6 million years is described and compared. Despite its fragmentary status, several relevant variables—such as the MSP, olecranon fossa breadth, and thicknesses of the medial and lateral pillars adjacent to the olecranon fossa—can be directly measured on the specimen. Furthermore, based on regression analysis in two large modern human samples and in the European Middle Pleistocene sample from the Sima de los Huesos (SH) site we estimated the BB of the Bodo specimen to be around 62–63 mm (with a range of 60–66 mm). Cross–sectional properties at the 35% level do not indicate an especially robust shaft compared with other fossil humeri, although the relative cortical area is large in comparison with some modern human samples. Sexual diagnosis is only approximate, but the Bodo humerus more probably comes from a male.
Comparisons of the Bodo humerus with African and European fossil and modern hominid samples revealed that the Bodo specimen displays distal humeral features very common in the European Middle Pleistocene humans from SH and the Late Pleistocene Neandertals, such as the relatively thin lateral and medial pillars and the relatively wide olecranon fossa (Carretero et al., 1997). Three out of the four African Homo distal humeri (KNM–WT 15000, Gomboré IB and Kabwe) ranging in age from 1.5 to 0.2 Ma (Table 1) display the distal humeral proportions more common in modern humans, while only Bodo shows the pattern more common in the European Middle Pleistocene hominids and the Neandertals.
In Africa, the postcranial fossils from the Middle and Late Pleistocene exhibit a surprising amount of morphological diversity (Pearson, 2000). The presence of Neandertal traits in some, but not all, specimens of H. ergaster, H. rhodesiensis and early H. sapiens, together with the greater similarity of only some African specimens, but not others, to modern humans, indicates that populations of early Middle Pleistocene African humans may have been physically diverse and that their skeletons may have contained a mosaic of modern and non–modern traits (Pearson, 2000). Moreover, different individuals from the African and Levantine samples often display different sets of features that distinguish them from studied collections of recent humans, a parallel situation seen in European Middle Pleistocene humans and Neandertals (Arsuaga et al., 1997). The Neandertal–like morphology of the Bodo distal humerus fits the explanation that there was polymorphism in several skeletal traits in the Middle Pleistocene of Africa, as was the case in European Middle Pleistocene humans. Some specimens on both continents would show the Neandertal condition. Through time this morphology became fixed in the classic European Neandertals but was lost in Africa.
The recurrent nature of most postcranial traits we study can help us understand the striking variation we encounter in the fossil record. More fossils are needed, particularly postcranial remains of Middle and early Late Pleistocene African hominids. The Bodo partial humerus is quite relevant in the context of these discussions.
We thank Middle Awash Research Project, funded by grants from the National Science Foundation, for allowing us to study the Bodo humerus. We thank the Ethiopian government and the National Museum of Ethiopia for access to original fossil materials. We thank the Human Evolution Research Center at the University of California at Berkeley for facilities and support. We derived much benefit from the discussions with our colleagues Tim White from the Human Evolution Research Center at the University of California at Berkeley, Nuria García, Rolf M. Quam, Ignacio Martínez and Ana Gracia from the Centro UCM–ISCIII de Investigación sobre Evolución y Comportamiento Humanos of Madrid, and Berhane Asfaw from the Rift Valley Research Service of Ethiopia. Tim White kindly supplied photographs and background on the discovery of the Bodo humeral specimen. We thank Bruce Latimer and Lyman Jellema from the CMNH for providing assistance and helpful access to the Hamann Todd skeletal collection. We especially thank Rebeca García, from the Laboratory of Human Evolution of the University of Burgos (LEH), who measured the Hamann Todd sample in the CMNH. Elena Santos and Ciaran Brewster (LEH) gave us general assistance with the preparation of manuscript. We are very grateful to the Anthropological Science editors, and especially to Dr Gen Suwa, for their help with the text. Laura Rodriguez was funded by a grant from Fundación Siglo para las Artes de Castilla y León. Financial support was obtained from the Dirección General de Investigación Científica y Técnicia of Spain (project CGL 2006–13532–C03–02) and from Junta de Castilla y León (project BU032A06).