| * Correspondence to: Yukio Dodo, Department of Anatomy and Anthropology, Tohoku University School of Medicine, 2–1 Seiryo-cho, Aoba-ku, Sendai 980–8575, Japan. E-mail: ydodo@plum.plala.or.jp Published online 19 September 2009 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase090622 |
In the first author’s earlier work (Dodo, 1987), incidence patterns of two cranial nonmetric traits, supraorbital foramen (SOF) and hypoglossal canal bridging (HGCB), were proven to be fairly effective for discriminating between major human population groups in the world. In that work, however, a certain amount of interobserver error might have been involved, because much of the frequency data of SOF and HGCB was cited from literature, in some of which the criterion for scoring SOF was rather ambiguous. Recently, Japanese researchers have reported extensive frequency data of cranial nonmetric traits in nearly 80 samples from all over the world (Hanihara and Ishida, 2001b, c, d, e). Conveniently, SOF and HGCB are included in their battery of traits, and the presence or absence of both traits is judged following the criteria proposed by Dodo (1974, 1987). This indicates minimal interobserver errors between their data and ours.
In the present study, we reexamine the frequency distribution of SOF and HGCB in cranial samples from various regions of the world, including a Jomon sample in Japan. The Jomon are a prehistoric foraging, hunting, and fishing people, who inhabited the Japanese archipelago from about 12000 to 2500 years BP, and are widely accepted as the main ancestral population of the present-day Ainu (e.g. Howells, 1966; Turner, 1976; Yamaguchi, 1982; Brace and Nagai, 1982; Ossenberg, 1986; Hanihara, 1991; Dodo and Kawakubo, 2002).
The objectives of this study are (i) to inquire whether the frequency distribution of SOF and HGCB is actually effective in distinguishing between major human population groups in the world; and (ii) to discuss how the observed worldwide frequency variations of SOF and HGCB can be interpreted from an evolutionary perspective.
Table 1 lists the regionally representative 71 cranial samples from various areas of the world, which consist of more than 50 individuals. Hanihara and Ishida (2001d, e, and personal communications) reported the frequency data of SOF and HGCB in all but two samples—those of modern Thai and prehistoric Jomon. Detailed information concerning the dates, ethnic groups, and countries of origin of these samples is presented in Hanihara and Ishida (2001a). Although several prehistoric and early historic samples are included, most of them are of recent times. The frequency data of the Thai sample were cited from Ishida and Dodo (1997), and those in the Jomon sample, extending from the early to the final stages (c. 6000–2500 years BP), were gathered for this study, in adding the western Japan Jomon series to those of eastern Japan previously reported (Dodo and Ishida, 1990, 1992).
In calculating the frequencies of SOF and HGCB, an individual counting method was used; if a trait was present on one or both sides, it was recorded as being present. Moreover, since, in both SOF and HGCB, the frequencies were not significantly different between sexes for most of the individual and regionally aggregated samples (Ossenberg, 1969; Mouri, 1976; Dodo and Ishida, 1987; Hanihara and Ishida, 2001d, e), their frequencies were based on sex-pooled samples.
The presence or absence of SOF and HGCB was recorded according to the criteria of Dodo (1974, 1987) and Dodo and Ishida (1990). Any foramen on the supraorbital margin of the frontal bone opening to the orbital cavity was scored as SOF present. Thus, the single foramen, lateral foramen, medial foramen, and a few cases of the supratrochlear foramen were included in this category. With respect to HGCB morphology, only a complete bridging was recorded as present.
Table 1 lists the number of crania and frequencies of SOF and HGCB in 71 cranial samples representative of 13 major regions of the world. Figure 1 depicts the frequency distribution of SOF and HGCB for these samples.
 View Details | Figure 1. Frequency distribution of the supraorbital foramen (SOF) and hypoglossal canal bridging (HGCB) in 71 cranial samples from various regions of the world. Symbols denoting 13 major regions of the world are: closed stars, Sub-Saharan Africa; open diamonds, North Africa; open squares, Europe; open triangles, South Asia; closed inverted triangles, Southeast Asia; closed squares, East Asia; closed triangles, Central Asia; closed diamonds, East Siberia; #, Arctic; *, Americas; open circles, Melanesia; double circles, Australia; open stars, Polynesia. |
The variation range of frequencies is large in SOF (18.1–79.7%) and, with the exception of Australians and Jomon/Ainu, the frequency tends to increase eastward from Africa, reaching a peak at Chúkchi Península. The frequency of HGCB varies from 5.5% to 41.5% and tends to be higher in samples in the Northern Hemisphere, excluding Peru and Fuego/Patagonia.
As seen in Figure 1, individual samples belonging to each of the 13 major regions are distributed within a limited area, and the three major population groups, Sub-Saharan Africans and the so-called ‘Caucasoids’ and ‘Mongoloids’ are fairly well distinguished. Sub-Saharan Africans and Australians are located in the lower left part; ‘Caucasoids’, consisting of North Africans, Europeans, and South Asians, occupy the middle position; and ‘Mongoloids’ in the widest sense, including Southeast Asians, East Asians, Central Asians, East Siberians, Arctic peoples, Native Americans, and Polynesians, shift to the right half of the figure. The Jomon and Ainu in Japan are outliers of the major human populations in the world.
The findings of the present study are roughly consistent with those of the previous study (Dodo, 1987) and with the results of multivariate statistical analyses using 20 cranial nonmetric traits (Hanihara et al., 2003). A couple of the SOF and HGCB morphs fairly well discriminate between the three major human population groups in the world. The exceptions are the Australo-Melanesians and Jomon/Ainu in Japan.
Consulting the originals, casts, color atlases, and literature of Plio-Pleistocene hominid fossils to examine the occurrence of SOF, it seems most likely that SOF first appeared in late Pleistocene early modern humans, with some possible exceptions, e.g. Stw 265 australopithecine (Lockwood and Tobias, 2002), Dederiyeh 2 infant Neanderthal (Ishida and Kondo, 2003), and La Qiuna 5 Neanderthal (cast). A list of the presence or absence of SOF in Plio-Pleistocene hominids is given in Table 2, in which a small number of juvenile fossils are included and, when one side of the supraorbital margin is missing, scoring of the trait is based only on the other side. Though its frequency in early modern humans is not high (16% or so), the SOF morph may be included in a series of derived modern human characteristics.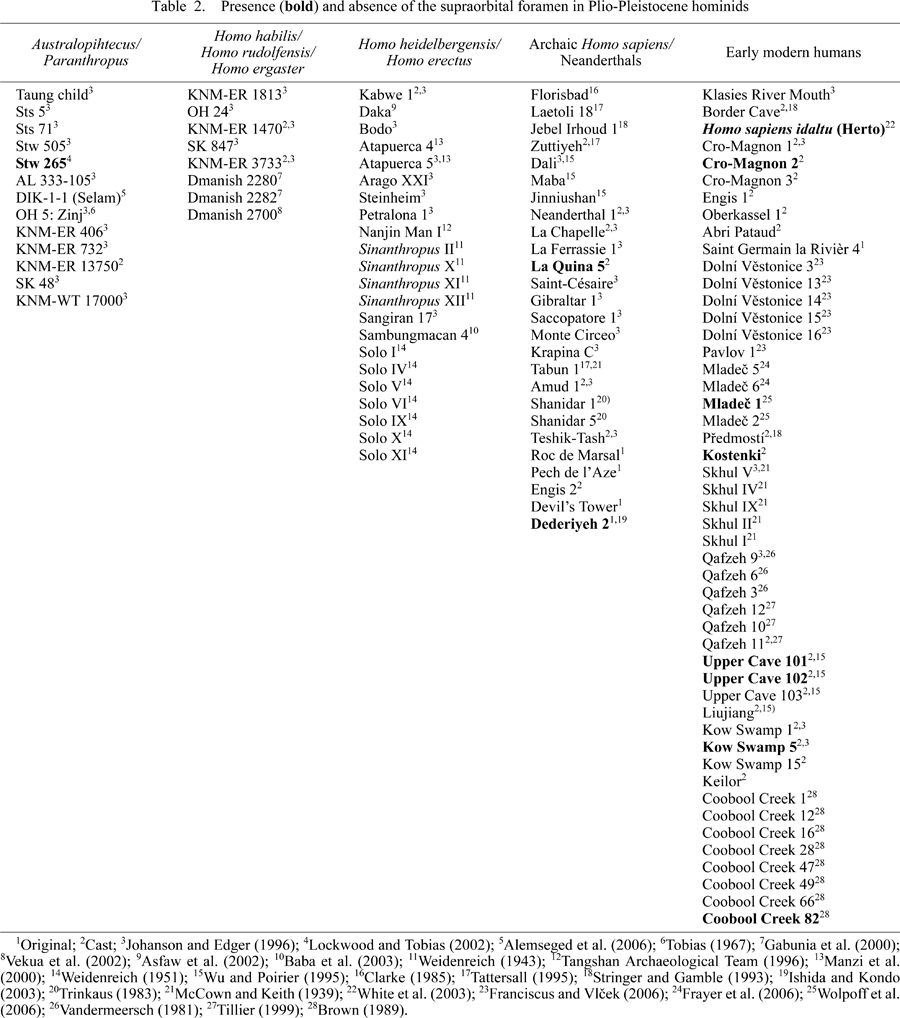
Lower frequencies of SOF are observed in Sub-Saharan Africans (25.5–44.1%), Australians (23.0–23.3%), and Jomon/Ainu (18.1–26.4%). The SOF frequency of the Jomon is the world’s lowest. The low SOF frequency in these populations may be the retention of an early modern human physical condition. Such an archaic condition appears to have been retained in the Jomon, who lived in the Japanese archipelago, considerably isolated from the Asian mainland, for about 10000 years (Imamura, 1996; Hanihara and Ishida, 2009), and in the Hokkaido Ainu, undoubtedly descended from the Jomon (Dodo and Kawakubo, 2002). A similar process must have occurred in Australia, though the term of isolation is much longer, 30000 years or more (Mulvaney and Kamminga, 1999). Sub-Saharan Africa may be the homeland of early modern humans, as many adherents of the single African origin hypothesis for modern Homo sapiens have claimed (see reviews in Stringer, 2002; Mellars, 2006).
The hypoglossal canal morphology can hardly be observed in cranial casts. It is also poorly represented in the fossil hominid anthropological literature, in which we could find descriptions of hypoglossal canal morphology for only 28 Plio-Pleistocene hominid skulls, as shown in Table 3. Again in this table, a few juvenile fossils are included and, when one side of the occipital bone including the hypoglossal canal is missing, scoring of HGCB is based only on the other side. Of the 28 skulls, HGCB is reported to be present in two: Paranthropus boisei (OH5-Jinj) and Dolní Vestonice 13. In contrast to the great apes (Pongo, Gorilla, and Pan), which have extremely high frequencies of HGCB (Braga, 1995), Plio-Pleistocene hominids, including early modern humans, seem to have a relatively low HGCB frequency (7.1% in the present survey).
If a low frequency of HGCB is the generalized condition in Plio-Pleistocene hominids, its lower frequencies in Sub-Saharan Africans (5.6–22.0%) and Australians (9.0–11.9%) may also be the retention of their ancestral fossil human condition. In this respect, the circumstances of the Jomon/Ainu were somewhat different from the case of SOF, with considerably high frequencies of HGCB in the Jomon (28.9%) and the Ainu (35.5%). The founder effect or local adaptation of the ancestors of Jomon may have been the main cause for this. In any case, however, a unique combination of archaic and derived conditions, i.e. a low frequency of SOF and a high frequency of HGCB, makes the Jomon/Ainu outliers of major human populations in the world, as seen in Figure 1.
The Jomon tendency to have an outlying position in the world has been sometimes interpreted based on the assumption that they retained a number of archaic characteristics shared with Eurasian Upper Palaeolithic hunter-gatherers (Yamaguchi, 1982; Ossenberg, 1986; Kamminga, 1992), and that they were in an incipient stage of modern human evolution before ‘racial differentiation’ (Kozintsev, 1992; Dodo, 2006). Such an assumption is partially supported in view of the world’s lowest SOF frequency in the Jomon, most likely the retention of an early modern human physical condition.
In this connection, if the lower frequencies of SOF and HGCB actually represent a late Pleistocene early modern human condition, it is emphasized that an incipient stage of modern Homo sapiens is reflected in Sub-Saharan Africans and Australians. For further discussion on this theme, we need much more information from human skeletal remains of the late Pleistocene and Early Holocene unearthed from the Old World.
We express our sincere thanks to H. Ishida of University of the Ryukyus and T. Hanihara of Kitasato University School of Medicine for personal communication and helpful advice. We are greatly indebted to Y. Mizoguchi and Y. Kaifu of the National Museum of Nature and Science for permission to study the cranial casts under their care and for useful comments. We also thank T. Nara of Nippon Dental University School of Life Dentistry at Niigata and Y. Nagura of Tohoku University School of Medicine for advice and technical support. This study is financially supported by Grants-in-Aid for Scientific Research (Nos. 20370096 and 20770198) from the Japan Society for the Promotion of Science.
|