| * Correspondence to: Aiko Saso, The University Museum, The University of Tokyo, Hongo 7-3-1, Bunkyo-ku, Tokyo 113-0033, Japan. E-mail: sasoyan@um.u-tokyo.ac.jp Published online 12 April 2011 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.100726 |
The Late Pleistocene human fossils recovered from the Minatogawa fissure deposits in southern Okinawa Island (Yaese-cho, formerly Gushikami-son) include four human partial skeletons (Minatogawa I–IV) (Suzuki and Hanihara, 1982). This human skeletal series (hereafter the Minatogawa series, or simply Minatogawa) represents the only substantial skeletal evidence of pre-Jomon prehistoric humans so far known from the Japanese archipelago. It also forms a part of the scarce and largely fragmentary Late Pleistocene Homo sapiens record of East Asia (e.g. Kamminga and Wright, 1988; Baba and Narasaki, 1991; Yamaguchi, 1992; and reviewed in Shang et al., 2007). Discovered in 1970–1971, Suzuki and collaborators have published an early monographic description and interpretations (Baba and Endo, 1982; Suzuki, 1982), followed by a series of evaluations by Baba (Baba and Narasaki, 1991; Baba et al., 1998; Baba, 2000a). Critical chronological evaluations suggest a possible age range of ~15–20 kyr (see Matsu’ura and Kondo, 2011, for details). More recently, additional morphological investigations of the Minatogawa series have been initiated (Kubo et al., 2008, 2011; Kaifu et al., 2011).
In the first descriptive and analytical account of the Minatogawa series, Suzuki (1982) considered Minatogawa to resemble the southern Chinese Liujiang more so than the northern Upper Cave crania. He interpreted the evidence to indicate that both Minatogawa and the Jomon were descendents of a southern Chinese ‘proto-Mongoloid’ population, and that Minatogawa and the Jomon had acquired their subsequent distinctiveness through isolation and gene flow patterns particular to each region (Suzuki, 1982). Yamaguchi (1992) made comparisons with a wider range of materials, and suggested that Minatogawa shows similarities not only with Liujiang, but also with Wadjak from Java. Wu (1992) found metric similarities with Liujiang, but not with Wadjak. Baba and colleagues expanded on previous studies and made systematic qualitative comparisons of the Minatogawa I skull with East Asian Late Pleistocene/early Holocene fossils (Baba and Narasaki, 1991; Baba et al., 1998). They considered Minatogawa to exhibit more similarities with Wadjak than with Liujiang, and suggested a Southeast Asian origin of Minatogawa.
These and other authorities (e.g. Hanihara, 1991) considered Minatogawa as broadly representative of, or having affinities with, populations ancestral to the Holocene Jomon. However, both genetic and paleolithic evidence suggests a contradictory continental northeastern source of the Jomon (Sagawa, 1995; Takamiya, 1996; Omoto and Saitou, 1997; Adachi et al., 2009; Hanihara and Ishida, 2009). Some have cautioned against a direct Minatogawa to Jomon relationship, particularly in Okinawa Island (e.g. Yamaguchi, 1992; Asato and Doi, 1999; Baba, 2000b, 2002; Takamiya and Obata, 2002). Recently, Kaifu et al. (2011), based on a broad comparative framework of mandibular morphology, suggested that the Minatogawa series probably derived from a Late Pleistocene southeastern Asian populational background with Australomelanesian affinities. They cautioned that the Japanese Holocene Jomon, lacking such morphological tendencies, probably represents a separate populational derivation stemming from northeastern Asian continental sources (Kaifu et al., 2011).
Glabellar morphology has been discussed by both Suzuki (1982) and Baba (Baba and Narasaki, 1991; Baba et al., 1998). Suzuki (1982) noted that a forward swelling of the glabellar region and a laterally flaring zygomatic process of the frontal bone (resulting in a broad upper face and narrow forehead) are among some of the most distinctive facial features of Minatogawa. Suzuki (1982) described the Minatogawa I and IV glabellar regions as formed by fusion of the developed glabella with the superciliary arches into a rhomboidal swelling that overhangs the nasofrontal suture. This produces a deeply receded nasal root especially in Minatogawa I (Suzuki, 1982). He apparently called the entire swollen area the “supraorbital ridge,” causing some confusion with the supraorbital torus (e.g. Baba et al., 1998). Suzuki (1982) also noted that Minatogawa I and IV’s glabellar area continues superiorly into a frontal “sagittal crest” (in actuality, an ill-defined weak keel), analogous to varying expressions of sagittal keeling observed in Holocene Jomon crania.
Baba and colleagues (Baba et al., 1998; Baba, 2002) described Minatogawa I’s glabellar region as swollen and projecting anteriorly, but not as extreme as in the Jomon males. Combined with a receding frontal squama, deeply recessed nasal root, and prominent nasal profile, this produces Minatogawa’s characteristic ‘rugged’ or ‘vertically flexed’ lateral facial profile. They interpreted these and other Minatogawa morphologies as a primitive retention from an Asian population prior to the emergence of cold-weather adaptations. They also categorized glabellar prominence into types 1 and 2 (Baba et al., 1998), in relation to superciliary arch projection. They classified both Minatogawa and Jomon glabellar regions as type 1 (glabella projecting more anteriorly than the superciliary arch).
Aside from its significance in evaluating the populational affinities of Minatogawa and other Japanese skeletal series (e.g. Suzuki, 1969, and citations above), supraorbital and glabellar morphology has featured prominently in studies that aimed to clarify the emergence and evolutionary patterns of anatomically modern Homo sapiens (e.g. Lieberman, 1995; Lahr, 1996; Lahr and Wright, 1996). A developed supraorbital ridge or prominence is often considered a primitive feature indicative of either mechanical or non-functional general robustness, and has been discussed from diverse perspectives including masticatory, spatiodevelopmental, and neutral genetic vs. environmental models (Russell, 1985; Lieberman, 1995; Lahr and Wright, 1996; Baab et al., 2010).
Supraorbital and glabellar morphology is typically evaluated by visual assessments and categorization of morphological expressions (e.g. Cunningham, 1909; Lahr, 1996; Rightmire, 1998; Lockwood, 1999). In the present paper, we provide an alternative three-dimensional (3-D) topographic evaluation, based on high-resolution laser scanning of the entire glabellar and superciliary regions. We apply this methodology to a comparison of the Minatogawa and Jomon glabellosuperciliary morphology. We consider the present study to be a preliminary one that serves two purposes. One of our aims is to objectively describe the Minatogawa glabellosuperciliary region using high-resolution 3-D data. The other purpose is to explore the potentials of a 3-D morphometric method in a Minatogawa–Jomon comparison. The major question we address is whether the protuberant glabellar morphology of Minatogawa can be seen as a possible ancestral condition to the Jomon, or whether it more likely represents a distinct morphological pattern.
The Minatogawa Late Pleistocene human skeletal series includes four partial skeletons, Minatogawa I–IV (Suzuki and Hanihara, 1982). The Minatogawa I skull (considered male) is nearly complete, while the Minatogawa II and IV partial crania (considered females) are less complete (Suzuki, 1982). In the present study, we describe the glabellar region of all three Minatogawa crania by 3-D topographic visualization. We then evaluate the best preserved and most-discussed male Minatogawa I by quantitative analyses of the 3-D morphometric data.
The comparative Jomon sample was selected from The University Museum, The University of Tokyo (UMUT) collections (Endo and Endo, 1979). The Jomon crania included in our examination comprised 129 male individuals from 36 sites. These range geographically from southern Hokkaido to northern Hiroshima, and chronologically from the Early to the Latest Jomon period (4000–500 BC). First, we conducted a visual evaluation of glabellar prominence. We ordered the 129 individuals in terms of glabellar prominence and then categorized these into three grades of development (Table 1). This revealed possible geographic variation, which needs to be confirmed/evaluated by further site samples and larger sample sizes. Because our sample was predominantly Middle to Latest Jomon (only three Early Jomon crania were included, from Hanazumi and Koyaba), the obtained pattern is suggestive of a north–south cline of glabellar robusticity during Middle to Latest Jomon times, and not a reflection of temporal differences within the Jomon period (the Earliest Jomon is considered to be more gracile, e.g. Ogata, 1981).
We then chose a subset of crania for 3-D evaluation, based on the following criteria: (1) minimal/no post-burial deformation; (2) minimal/no surface bone damage, so that interpolation of missing data would not be necessary; and (3) only one specimen from each site, so as not to bias the small comparative samples by site-specific tendencies. This confined the sample of grade 3 (strong glabellar protrusion) crania to only five measurable specimens (Table 2). As outlined above, because our major focus of enquiry was morphological comparisons between Minatogawa I and Jomon individuals with strong glabellar protrusion, we limited the grade 1 and 2 crania to five specimens each so that statistical analyses would not be biased by the more numerous grade 1/2 specimens.
High-resolution 3-D surface data of the entire upper facial region were taken by a laser scanning device (Vivid 910, Konica-Minolta). This was done with the cranium in the Frankfurt horizontal orientation. An individual scan records data points with a resolution of approximately 300 micron. Scans of the Minatogawa or Jomon face were taken from three directions (three scans per specimen, in anterior view and 30° oblique views from each side) and these were merged using the Polygon Edging Tool version 2. This software includes an automated routine that fine-tunes scan alignments by 3-D best-fit algorithms, as well as choosing the more accurate (better angled) points in areas of overlapping scans. Specifications of the resulting aligned and merged scan data sets are given as ±0.03 mm precision and approximately 0.05 mm accuracy. Typically, a scan covering most of the frontal squama and both maxillas would be represented by approximately 240000 points (Figure 1).
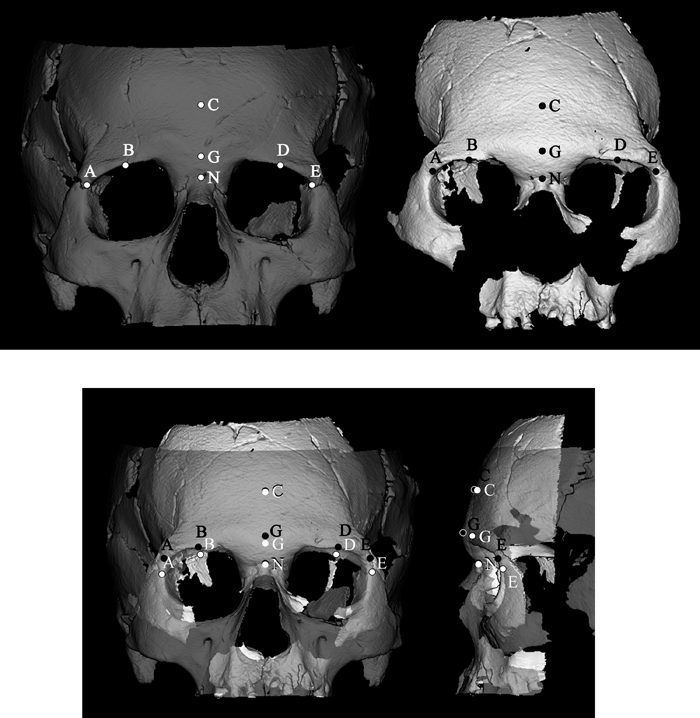 View Details | Figure 1. Landmarks used for superimposing each cranium with an operational reference cranium (UMUT 131377). Left (grey), UMUT 131377; right (white), Minatogawa I. Points A and E, frontmalare orbitale; B and D, superior orbital margin points at mid-orbit; C, median point above nasion at a distance equivalent to 1/3 of bi-frontmalare orbitale breadth; G, glabella; N, nasion. Using these seven points, the affine transformation that minimizes the summed squared distances between each cranium and the reference cranium (UMUT 131377) was determined. The entire data set of each cranium was then rotated around the x-axis by the derived amount, and then translated in three axes so that nasion coincides. |
We then extracted height values of positionally homologous 3-D data points of the glabellar and surrounding regions for morphometric comparisons among specimens. This was done by first scaling each specimen to a biorbital breadth (between frontomalare orbitale) equivalent to that of an operational reference cranium (UMUT 131377). Next, because height values of these data points are influenced by frontal squama orientation, we took the additional step of fine-tuning orientation by superimposing seven landmarks (or positionally defined points). The points we used were glabella, nasion, both frontmalare orbitales, bilateral superior orbital margin points at mid-orbit, and a median point on the frontal squama above nasion at a distance equivalent to 1/3 distance of bifrontmalare orbitale breadth (Figure 1). Each cranium was realigned by a rotation factor determined from least-squares best fit against the reference cranium (UMUT 131377), involving rotation around the x-axis, and thereafter translation so that nasion corresponds. The nasion of the reference cranium (UMUT 131377) was set to a height value of 15 mm. Finally, a 21 × 13 point grid (in 3 mm intervals in the reference cranium) was defined in frontal plane orientation (Figure 2), and the height (perpendicular to the frontal plane) was recorded for each of the up to 273 points that span the 60 mm × 36 mm area (in the reference cranium) in the frontal view. The measured area covers a region that includes approximately the medial halves of both orbits.
 View Details | Figure 2. The area sampled in principal components and cluster analyses. A 21 × 13 point grid system was defined in frontal view, and height values of approximately 240 points were used in the quantitative morphological analysis. The grid lines correspond to 3 mm intervals in the reference cranium (UMUT 131377). See text for further details. |
Quantitative morphological analysis of the 3-D data was done by principal components and cluster analyses of the heights of the approximately 240 points that fall on the cranial surface (of the total of 273 points of the grid system). Cluster analysis was done by UPGMA clustering of the Euclidean distance matrix derived from all points with pairwise deletion of missing values. 3-D data handling, measurement, and visualization were done using 3D-Rugle version 4, and most statistical calculations with Systat version 10. The principal component axes were morphologically expressed by deforming the 273-point grid by the eigenshape method (Nagata et al., 1996; Isono et al., 1999; Kuratate et al., 1999, 2003; MacLeod, 1999) using the routines Bone Restorer and Proc 3D written by one of us (S.M.).
All three Minatogawa and 15 Jomon crania are shown in Figure 3. In the Jomon crania, the glabellar and superciliary prominences are fused to varying degrees, but are usually bilaterally distinct and tend to form an inverted triangular to trapezoid region of protrusion. Some Jomon individuals exhibit a more rhomboidal eminence. However, these are still asymmetric (superoinferiorly) because of an inferolaterally concave outline (corresponding to the medial orbit margin and superciliary arch contours) and a variable transition to the frontal squama. In some specimens, a median keel-like prominence extends superiorly. Either the glabella or the medial superciliary arch projects anteriorly the most.
 View Details | Figure 3. Glabellar region topography of the three Minatogawa and 15 Jomon crania. The Jomon and Minatogawa I crania are shown in standard orientation with color grading of distance above the coronal plane (set at 15 mm below the nasion in the reference cranium, UMUT 131377) (see the text for further details). Each cranium was scaled to the biorbital breadth (between frontomalare orbitale) of the reference cranium, so that the color gradings represent size-adjusted relative values of topography. Minatogawa II and IV exhibit only weak absolute protuberance, and are shown here in enhanced color grading, with maximum glabellar protuberance adjusted to match that of Minatogawa I. Specimen codes are as follows: HNZ, Hanazumi; HOB5, Hobi (1941) 5 (secondary individual); HSU1, Hosoura (1919) 1; KSM6, Kasori-Minami (1964) 6; KSK, Kosaku (1926); KYB2, Koyaba 2; MTD, Matsudo; MTW7, Motowanishi 7; NKZ3, Nakazawahama (1926) 3; SNG22, Sanganchi (1952) 22; SNG117, Sanganchi117; TKM21, Tsukumo (1919) 21; UBY8, Ubayama (1962) 8; UBY9, Ubayama (1962) 9; UBY16, Ubayama (1950) 16. The overlap in maximum glabellar projection between the grade 2 and 3 specimens stems from the combined effects of biorbital size standardization (i.e. the depicted maximum protuberance is not the actual absolute value) and differences between grade criteria and 3-D measurement protocol. |
The Minatogawa glabellar region appears distinct from that of the Jomon. In Minatogawa I, the superciliary region is devoid of any arch-like expression (weakly retained in Minatogawa IV). The median swelling extends superiorly and laterally in a diffused manner, completely obliterating (Minatogawa I) or only weakly preserving (Minatogawa IV) the superciliary arch-like structure. Suzuki (1982) described this morphology common to both Minatogawa I and IV as glabella and superciliary arches that are “fused” into a “rhomboidal supraorbital ridge.”
The frontal bone nasal process of Minatogawa I is superoinferiorly short and swollen across its anteroinferior surface. This swelling is conspicuous and extends laterally, contributing to the oval to rounded rhomboid pattern of the entire glabellosuperciliary prominence. 3-D enhancement of the weaker glabellosuperciliary prominence in the female Minatogawa II and IV demonstrates the same pattern as seen in Minatogawa I (Figure 3). However, Minatogawa IV retains a faint superciliary arch expression. We interpret this as due to the weaker overall swelling of the Minatogawa IV central eminence compared with Minatogawa I. Minatogawa I is shown as having a moderately strong anterior protrusion of the glabella, but its degree of protrusion is not particularly extreme compared with the Jomon.
Suzuki (1982) also noted that both the Minatogawa I and IV glabellar regions “stretches upward as the sagittal crest to the bregma.” We agree that a median keel-like extension runs superiorly onto the frontal squama, but this is faint in Minatogawa I and only slightly better defined in Minatogawa II and IV, somewhat in contradistinction to Suzuki’s (1982) description of a “crest” extending to bregma. In Jomon crania, the median frontal eminence often takes the form of a ‘pinched’ or ‘keeled’ structure flanked by slight bilateral concavities. This is not seen in the Minatogawa crania.
Results of the PCA of the height values (covariance matrix) are shown in Figure 4. The first five principal components (PCs) account for a cumulative 93.9% of the variance. The pattern of shape variations shown in Figure 4 indicates that PC1 (53.5% of variance) contrasts bilaterally prominent superciliary arches (with a horizontally flat lower frontal squama) against a prominent midline region with lack of superciliary arch differentiation. In the PC1 score, Minatogawa I lies in the lower (weak or lack of bilateral separation) Jomon range of variation.
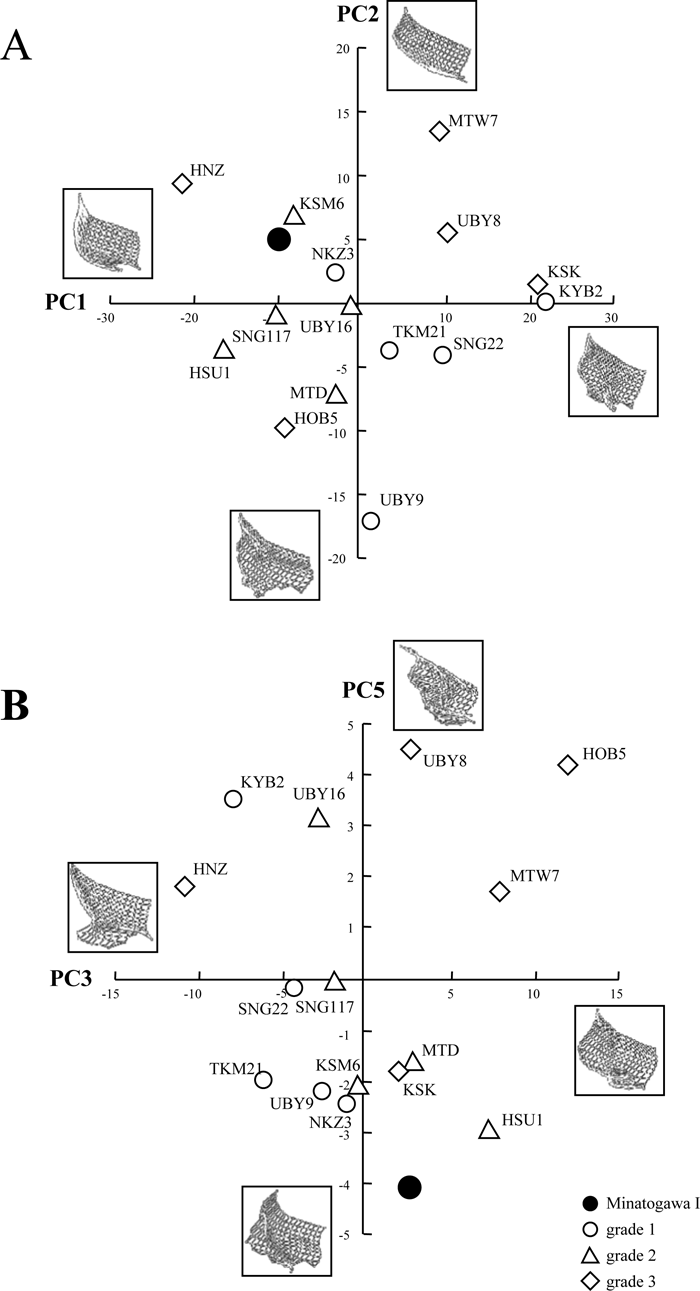 View Details | Figure 4. Results of the principal component analysis (PCA) performed on Minatogawa I and the Jomon specimens. (A) PC1 vs. PC2; (B) PC3 vs. PC5. Shape variation is visualized by wireframe deformation of grid points calculated from two standard deviation positive or negative scores. Specimen labels as in Figure 3. |
PC2 (20.5% of variance) appears to reflect some aspect of glabella and superciliary projection, but is not necessarily a general protrusion factor. PC3 (12.3% of variance) appears to reflect the degree or abruptness of nasal root recession, and Minatogawa I lies at the positive (root receding) side of the Jomon range. PC4 (4.9% of variance) seems to reflect artifacts of registration methods and is not considered here. PC5 (2.7% of variance) appears to reflect development of a circumscribed median zone of glabellar protrusion, in which Minatogawa I exhibits an extreme score. We also performed PCA without Minatogawa I and confirmed that the results were similar for PC1–PC4, but an equivalent of PC5 was not found. This indicates that PC5 of the initial analysis reflects the distinctiveness of Minatogawa I, albeit of small magnitude in proportion to the entire 3-D topographical variance of the current Jomon sample.
The results of the cluster analysis are shown in Figure 5, which appears to reflect the shape/pattern rather than the degree of prominence. Although there seems to be some tendency to group the grades 1 and 2 (weaker glabellar region) specimens together, individuals of different grades with similar morphologies are also joined. The latter example includes the Motowanishi 7/Kosaku/Koyaba 2 cluster which are characterized by distinct bilateral arches and weak superior crest/eminence extensions, and Nakazawahama 3/Kasori-minami 6 which clusters with Minatogawa I. The Jomon crania of the latter cluster exhibit ill- or non-separated superciliary arches that are only weakly distinct and tend to blend with the median prominence. This results in a relatively symmetric rhomboidal prominence, but differs from Minatogawa I in their weaker superior swelling. Kasori-minami 6 (and less so Nakazawahama 3) also exhibits an inferolaterally concave eminence contour, typical of Jomon crania (associated with a superoinferiorly tall nasal process of the frontal) and differing from Minatogawa.
 View Details | Figure 5. UPGMA cluster analysis of Euclidean distance matrix of the Minatogawa I and comparative Jomon specimens, based on height values of the grid points. Topographic images of each specimen are shown in color grading, adjusted to equivalent maximum and minimum values so as to enhance topographic patterns. White, maximum height in each specimen; blue, minimum height in each specimen. Specimen codes are shown in Figure 3. |
A “forward swelling” glabellar region has been described by Suzuki (1982) to characterize both Minatogawa I and IV. In our 3-D topographic analysis, we were able to visualize this morphology effectively, as a rhomboid-to-ovoid swollen area lacking (or only weakly retained in Minatogawa IV) expressions of differentiated superciliary eminences. In contrast, the Jomon patterns were variable but exhibit distinct expressions of bilateral superciliary structures. Even in cases when this was weak (KSM6, HNZ, HSU1), glabellar swelling is not developed superiorly, thereby differing from the Minatogawa condition. The Jomon crania also differ in their inferolaterally concave contour of the eminence (Figure 3). Inspection of the original specimens suggests that the latter is related to subtle distinctions seen in the nasal root region. In Minatogawa I, the nasal process of the frontal bone is short superoinferiorly, so that its swollen anterolateral surface forms a part of the ovoid outline of the entire glabellosuperciliary eminence. In contrast, in the Jomon crania, the nasal process is taller, leading to an inferolaterally concave eminence contour. Suzuki (1982) described the Minatogawa I glabellar swelling as overhanging the frontonasal suture and forming a deeply receding nasal root. In Minatogawa I, due to the above-noted nasal process morphology, the abrupt recession of the nasal root also extends laterally to the frontomaxillary suture. Such a morphology is not observed in the Jomon crania, in which the frontomaxillary suture often faces more laterally.
Minatogawa IV exhibits a similar pattern of glabellosuperciliary swelling as seen in Minatogawa I, but at a weaker level of expression. Minatogawa II, which has only a very faint glabellar topography—not characterized by Suzuki (1982)—is here shown by topographic enhancement to actually exhibit the same morphological pattern as found in Minatogawa I and IV (Figure 3).
With regards to Baba et al.’s (1998) categorization of relative projection of the glabella itself, our 3-D dataset confirms that all three Minatogawa crania belong to their type 1 (glabella projecting more anteriorly than the superciliary arch). The Jomon crania examined here mix types 1 and 2 (superciliary arch projecting anteriorly; KSK, MTW 7). We also confirmed Baba et al.’s (1998) observation that absolute glabellar projection of Minatogawa I is not necessarily strong. Of the 10 Jomon crania classified as grades 2 and 3 by our qualitative assessments, based on our 3-D analysis, four Jomon crania (HUS1, KSM6, grade 2 and MTW7, HOB5, grade 3) have a more projecting glabella region (relative to nasion) than Minatogawa I.
The results of the PCA were less clear-cut, probably because our PCA combines a single Minatogawa specimen with 15 Jomon crania. By necessity, the resulting components predominantly express within-Jomon variation. This was the case for PC1–PC3 for which Minatogawa I exhibited factor scores within the Jomon range of variation. In PC1, a factor related with differentiation of the superciliary arches, Minatogawa occupied a position at the lower Jomon range of variation. Only in PC5, which appears to express development of a locally swollen glabellar area, did Minatogawa I exhibit an extreme score.
Cluster analysis also shows that the distinctiveness of Minatogawa I is slight, the latter securely nested within Jomon variation. This indicates that, in overall glabellar region/lower frontal topography, the magnitude of within-Jomon variation is far greater than specific Minatogawa–Jomon differences. This is not unexpected, since the present method positively weighs features/patterns that are area-wise more extensive and prone to greater variation in magnitude, but does not contradict the qualitative Minatogawa–Jomon differences documented above by 3-D visualization. We note that the Jomon crania that formed a cluster with Minatogawa I are all characterized by ill-differentiated bilateral superciliary arches and weak/diffused superior extensions onto the frontal squama. Because the distinctive Minatogawa morphologies concern a small area around glabella and the medial orbital region, its impact on both the PCA and the Euclidean distance matrix-based cluster analysis was probably de-emphasized. This also suggests that further sampling of the Minatogawa series population is needed to confirm the strength of the distinct morphological tendency suggested by the available specimens.
The above results and analyses lead to the tentative conclusion that Minatogawa’s glabellosuperciliary morphology is best considered as distinct from the Jomon condition. In particular, all three crania so far known were shown to exhibit a common pattern, regardless of the observed range of prominence (from strong in Minatogawa I to hardly expressed in Minatogawa II). In contrast, Jomon crania that tended to exhibit a Minatogawa-like rhomboid-shaped prominence were found to be a minority (less than a third), and furthermore differed in degree of superior swelling and inferolateral eminence contour.
Several prospects arise from these observations relating to the question of within-Jomon temporogeographic variation. Our current Jomon sample is confined to the post-4000 BC time period of predominantly eastern Japan (eastern Honshu and Hokkaido), with a more limited western Honshu representation. Although detailed evaluations of our larger sample of ~130 Jomon crania is outside the scope of the present study, cursory observations suggest the lack of a strictly Minatogawa-like morphological pattern as defined above. We therefore tentatively conclude that the glabellosuperciliary morphology of Minatogawa and the post-mid-Holocene Jomon were distinct.
Of particular significance would be to examine a fuller range of variation in Jomon assemblages, temporally and geographically more proximal to the Minatogawa remains. For northern and eastern Jomon, the question would be if a Minatogawa-like morphology would increase in frequency with chronological antiquity, as hypotheses of a Minatogawa-like ancestry to Honshu Jomon populations would predict (e.g. Suzuki, 1982; Baba et al., 1998). Of additional interest would be to document the temporogeographic occurrences (or absence) of a Minatogawa-like morphology in the southern Japanese Jomon populations of Kyushu and their time-equivalents in the Okinawa Islands. Accumulation of such comparative assessments would eventually contribute to a discussion of whether populations represented by the Minatogawa fossils were likely continuous at least with Holocene Okinawa populations as surmised by Suzuki (1982), and whether or not they had an influence in forming the more northern Jomon populations.
From 3-D-based topographic analysis, including qualitative visualization, PCA and cluster analysis, we conclude the following:
1. Minatogawa I exhibits a distinct rhomboid-to-ovoid glabellosuperciliary prominence, in the form of an undifferentiated swollen structure that encompasses both glabellar and superciliary arch regions. Most notably, contrary to relatively similar Jomon examples, the central swelling is diffuse and extends both superiorly and inferolaterally. Accompanied by a short nasal process of the frontal, this leads to a relatively ovoid swelling lacking a concave inferolateral contour (contra Jomon morphology) and the virtually complete obliteration of bilateral superciliary expression.
2. Minatogawa II and IV also exhibit the same morphological pattern at weaker absolute levels of expression (with weak superciliary expressions retained in Minatogawa IV). Together with the observation that absolute glabellar projection of Minatogawa I is not as great as the developed Jomon examples, this indicates that it is the pattern that is distinctive of Minatogawa, rather than development of glabellar region prominence itself.
3. The Minatogawa glabellosuperciliary morphological pattern, as characterized here, can be used as a tool to assess the possible relationships of the Late Pleistocene Minatogawa remains with Jomon-age equivalent populations of the Okinawa Islands, on the one hand, and Kyushu and eastern/northern Jomon populations, on the other.
The authors wish to sincerely thank Hisao Baba and other researchers at the Department of Anthropology, National Museum of Nature and Science, Tokyo, for their comments and suggestions over the years that this work was conducted. We are also grateful to Takashi Fukumoto, Kazuaki Kanei, Satoshi Tamura (Niigata University), and members of the paleoanthropology laboratory at The University Museum, The University of Tokyo, for their invaluable assistance to the present study. Toyohisa Tanijiri of Medic Engineering Corporation provided advice about 3-D data analysis.
|