| * Correspondence to: Gen Suwa, The University Museum, The University of Tokyo, Hongo 7-3-1, Bunkyo-ku, Tokyo 113-0033, Japan. E-mail: suwa@um.u-tokyo.ac.jp Published online 27 July 2011 in J-STAGE (www.jstage.jst.go.jp) DOI: 10.1537/ase.110617 |
Although dental size and morphology have contributed to population history studies in Japan (e.g. Brace and Nagai, 1982; Turner, 1987; Matsumura, 1994), little attention has been paid to root size and dimensions. This stems from the relative inaccessibility of tooth roots for measurement. Another potential limitation is the greater variability reported, for example, in root lengths (Ozaki et al., 1987).
We report here computer tomography (CT)-based metrics of mandibular tooth roots in the prehistoric Jomon and modern Japanese, and explore the utility of root system metrics in evaluating population history. We furthermore investigate root size and proportions of the 15000–20000 BP Late Pleistocene Minatogawa human remains of Okinawa Island (see Matsu’ura and Kondo, 2011; Kaifu and Fujita, 2011; and Suwa et al., n.d.) for recent chronological assessments). The Minatogawa Late Pleistocene remains were initially described by Suzuki and Hanihara (1982), and thereafter investigated primarily by Baba and colleagues (e.g. Baba and Narasaki, 1991; Baba et al., 1998; Baba, 2000). More recently, some newer studies have been conducted, either from a broader comparative basis and/or a fuller consideration of the more fragmentary evidence (Kaifu et al., 2011; and this study), by extending past observations via application of three-dimensional imaging methods (Kaifu et al., 2011; Kubo et al., 2011; Kubo and Kono, 2011; Saso et al., 2011; and this study), or from a refined statistical standpoint (Mizoguchi, 2011).
The landmark study of Brace and Nagai (1982) on dental size demonstrated that subtle distinctions in tooth crown size among the Japanese populations add up to an interpretable pattern of microevolutionary change. Specifically, they showed that the total sum of crown areas (buccolingual multiplied by mesiodistal diameters) differs substantially among the modern and prehistoric Japanese, with the indigenous Jomon having much smaller teeth than the immigrant Yayoi. The modern Japanese was shown to have slightly smaller teeth than the Yayoi, which was interpreted as reflecting a largely Yayoi ancestry with subsequent tooth size reduction.
Today, a diverse set of cranial, dental, and genetic evidence on Japanese population history suggests a dominant genetic influence of the Yayoi period (c. 2350–1650 BP) immigrants (e.g. Yamaguchi, 1985, 1987; Hanihara, 1987, 1991; Turner, 1987; Dodo and Ishida, 1990; Matsumura, 1994; Omoto and Saitou, 1997), but with substantial admixture with the indigenous Jomon people (e.g. Yamaguchi, 1982; Hanihara, 1991; Nakahashi, 1993; Kaifu, 1997). However, the timing, extent, and nature of admixture are regionally and temporally complex and the subject of continued study (e.g. Suzuki et al., 2004; Nakahashi and Iizuka, 2008; Adachi et al., 2009; Hanihara and Ishida, 2009; Ishida et al., 2009; Kawakubo et al., 2009; Matsumura and Dodo, 2009; Matsumura et al., 2009; Sato et al., 2009; Fukumoto and Kondo, 2010; Kaburagi et al., 2010).
Regarding the details of dental size, according to Brace and Nagai (1982), the Jomon had summed crown areas about 10% smaller than the Yayoi, while a 6.5% difference was observed between the Jomon and the larger-toothed modern Japanese. Since then, Matsumura (1994, 1995a) have compiled more extensive datasets of dental size through time and space, and largely confirmed the results of Brace and Nagai (1982). He also suggested a dichotomous difference of crown size proportions between the Jomon and the Yayoi/modern Japanese (Matsumura, 1994). He showed that the Jomon, Ainu, and some southern Japanese populations have relatively large first molars and smaller canines, premolars, and second molars, and the Yayoi, Kofun, and the modern Japanese the opposite condition.
Although Matsumura (1994) expressed tooth size and proportions predominantly by multivariate summary statistics, examination of his published data indicates a crown size difference between the Jomon and Yayoi/modern Japanese comparable to that formerly reported by Brace and Nagai (1982). His data suggests a close to 6% difference in total crown area between the Jomon and modern Japanese, with 4% stemming from the mesiodistal and 2% from the buccolingual diameters (inferred from Matsumura, 1994, 1995a). He also showed that a simple index between the first molar (M1) and canine, premolar, and/or M2 sizes can be useful in population distinctions. In these comparisons, mesiodistal diameters seem more diagnostic than buccolingual diameters. This contradicts the intuition that mesiodistal crown lengths are potentially less reliable because of wear-related reduction (see also Brace and Nagai, 1982). Recent genetic studies suggest considerable independence between the mesiodistal and buccolingual crown diameters (Hlusko et al., 2006), and enamel thickness also affects the two crown dimensions differentially (e.g. Kono et al., 2002). Presumably, omission of specimens with heavy wear (Brace and Nagai, 1982; Matsumura, 1994, 1995a) preserved the population signals.
In the present study, we first test the hypothesis that significant differences exist between the Jomon and the modern Japanese in mesiodistal dimensions of the root system (measured at the cervix). It is of some interest to confirm that mesiodistal cervical diameters faithfully track population differences in crown lengths, because substantial tooth wear often obscures actual crown dimensions especially in the fossil record.
Next, tooth size of the Minatogawa fossils, whose mesial and distal crown faces are considerably worn, are evaluated in terms of the mesiodistal cervical diameters. Previously, Hanihara and Ueda (1982) evaluated the crown size of the Minatogawa I individual by multivariate statistics of mesiodistal crown dimensions combining both upper and lower teeth. They concluded that Minatogawa I is characterized by large teeth, especially the molars, and by relatively small incisor, canines, and premolars. However, most of the anterior teeth (incisor, canine, and premolar) included in their analysis are particularly worn (see Plate 3.1 of Hanihara and Ueda, 1982), and hence their results on relative size and proportions (superficially similar to ‘robust’ Australopithecus) are probably biased. Matsumura (2000) also published an analysis (albeit brief) of the mesiodistal and buccolingual crown dimensions of Minatogawa I, but cautioned that these mesiodistal lengths are heavily reduced by wear. Based on buccolingual dimensions, he noted the large size of the Minatogawa I teeth (relative to the Jomon condition), as well as its relatively small canines and premolars, which he described as Jomon-like.
Finally, we examine root length (cervix to apex) and proportions of the modern Japanese, Jomon, and Minatogawa fossils. Our aim is to examine if and how root size differences mirror crown size patterns among populations, and to interpret the microevolutionary significance of the modern Japanese, Jomon, and Minatogawa conditions.
There is some support for the hypothesis that root surface area (via periodontal support capacity) relates to occlusal load (e.g. Spencer, 2003; Kupczik and Dean, 2008), and hence for a given root shape, root length potentially qualifies as a proxy for relative occlusal load. Furthermore, inasmuch as crown and root sizes are structurally linked through the cervix, covariance between the two via selection for tooth support capacity is possible. However, within-group correlations of crown size and root lengths are generally weak (Garn et al., 1978; Ozaki et al., 1988), reflecting considerable morphogenetic independence of the two (Ikuta and Shimizu, 2005). This and the high individual variation of root lengths (coefficients of variation can be close to twice as large as those of crown dimensions (Rosing, 1983; Ozaki et al., 1987)) caution against supposing a simple relationship between root lengths and occlusal load in population comparisons, especially when sample sizes are small.
Nevertheless, although comparative assessments are scarce in modern humans, Pleistocene Homo sapiens, or archaic Homo, recent studies do consistently suggest that pre-modern populations are characterized by larger roots (e.g. Bailey, 2005; Walker et al., 2008; Kupczik and Hublin, 2010; Hershkovitz et al., 2011). Accumulation of such data may eventually enable us to establish baseline root size conditions for population groups/lineages and/or broad adaptive (occlusal loading) conditions, in turn enabling interpretations of regional and/or local microevolutionary trajectories of root size patterns.
The mandibular skeletal remains of a total of 31 ‘modern’ Japanese and 26 prehistoric Jomon individuals housed in The University Museum, The University of Tokyo were examined. In this study, we confined our investigations to males, which are better represented in the collections. The Jomon materials span a chronological age of approximately 5000–2350 BP, and the ‘modern’ Japanese are from the anatomy department collections formed at the end of the 19th/early 20th century. Further details of the samples are available in Fukase and Suwa (2010).
The Minatogawa human fossils that we examined include the mandible of the Minatogawa I individual and mandibular specimens A, B, C, and D (Suzuki and Hanihara, 1982; Kaifu et al., 2011) (Figure 1). The Minatogawa I mandible preserves a damaged right C root and the right P3–M3 and left C–M3 crowns and roots. Kaifu et al. (2011) suggested that the left canine, which is glued into the alveolus, may belong to a different individual, based on preservation differences (darker colored) and space between the root apex and alveolar bone (observed in the CT imagery). However, a comprehensive look at the imagery indicates highly congruent root and alveolar surfaces, except at the possibly broken root apex. We (at least one of the authors, G.S.) therefore think it probable that the left canine in fact belongs to the Minatogawa I individual, despite its slightly different state of fossilization. The analysis below considers the canine as belonging to Minatogawa I, but its treatment as an isolated canine does not require any significant change to the results and discussion.
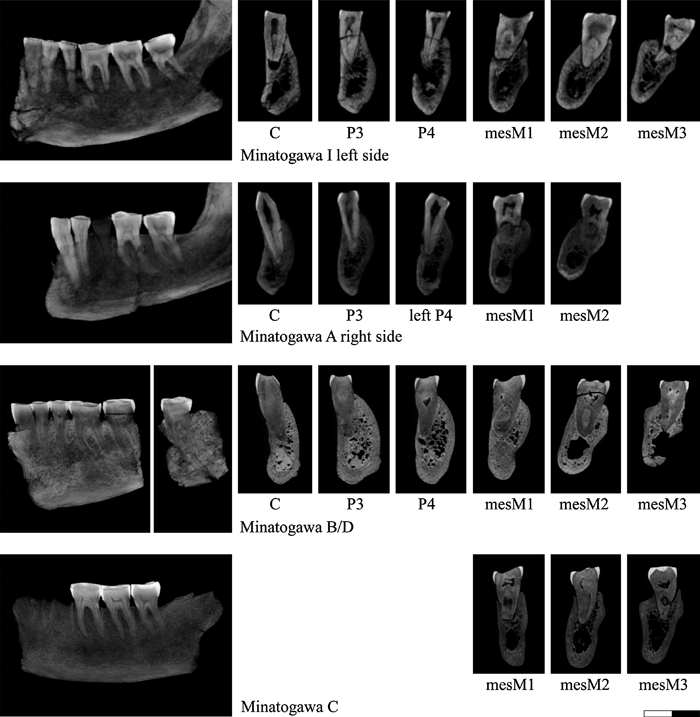 View Details | Figure 1. Subalveolar morphology of the Late Pleistocene Minatogawa fossil humans. The left column images are the summed-voxel projections of the serial CT stacks, rotated so that canine to molar rows are approximately in line. The right-hand side cross-section images of each tooth root are the multiplanar sections running approximately through mid-buccal cervix and root apex. Note that the root metrics were not taken from these images, but by defining landmarks using the volume dataset (see the text for details). All sections are shown with buccal to the left, regardless of whether the sections are of the right or left side corpus. Note that in mandible A, the right P4 is missing, so that the opposite side P4 section is shown. |
Mandible A preserves the right C–M2 and left P4–M2 crowns and roots. Its right P4 has been extracted and was not examined in the present study. Mandible B, a partial right corpus, preserves the C–M2 crowns and roots, and the fragmentary mandible D preserves the left M3 crown and root. Kaifu et al. (2011) suggested that mandibles B and D belong to the same individual, a proposition that we follow here. Mandible C preserves the right M1–M3 crowns and roots.
Following Suzuki (1982) and Kaifu et al. (2011), we consider Minatogawa I and mandible B/D males and mandibles A and C females.
Serial cross-sectional images of the mandibles were obtained for each specimen by use of the microfocal X-ray CT system (TXS225-ACTIS, Tesco) of The University Museum, The University of Tokyo. The CT imagery was taken at 130 kV, 0.24 mA. Slice thickness and interval were set at either 320 or 450 μm (18 of the modern Japanese were taken at the latter resolution) for the Jomon/modern Japanese specimens. Cross-sectional images were reconstructed in a 512 × 512 matrix with pixel size equivalent to the slice interval/thickness, producing isotropic voxels. The Minatogawa mandibles were similarly scanned at isotropic voxel sizes of 380 μm (Minatogawa I), 320 μm (mandible A), or 140 μm (mandibles B–D).
CT image processing and analysis were carried out using the softwares Analyze 7.0 (Mayo Clinic) and CT-Rugle (Medic Engineering). Both mesiodistal cervical diameter and root length (cervix to apex) were taken by (1) determining the three-dimensional coordinates of landmarks (in the stacked serial volume dataset), and (2) calculating the linear distances between landmark pairs that define the metrics. Mesiodistal cervical diameters were taken between mid-mesial and mid-distal cervical points (P3–M3) or at maximum cervical line concavity (I1–C). These points were determined by first extracting the horizontal slices that each pass through the mid-mesial and mid-distal face cervix, and then applying the half-maximum-height subpixel interpolation routine (CT-Rugle) to determine the tooth–air interface. This procedure results in metrics equivalent to the mesiodistal cervical diameters defined in Hillson et al. (2005). I1–P4 root lengths were defined as the distance from the root apex to the lowest point of the labial/buccal cervical line. Molar root lengths were taken from the mesial root apex to the buccal cervical line at a position below the middle of the mesiobuccal cusp (protoconid). For these points, the closest pixel of the root–air interface was visually determined in Analyze (in horizontal slices), and taken to represent the cervical or apical points. To lessen measurement error effects, the right and left sides were averaged when possible.
Jomon and modern Japanese
The basic statistics of the mesiodistal cervical diameters are given in Table 1, and the means depicted in Figure 2. As with mesiodistal crown diameters (Matsumura, 1995a), the Jomon and the modern Japanese differ significantly in mesiodistal cervical diameters of most tooth elements. The means of the Jomon were smaller than the modern Japanese by 3–9% (except for the M1 and M3), and add up to a summed I1–M2 or I1–M3 difference of about 4% (Jomon smaller), comparable in magnitude to the crown size differences.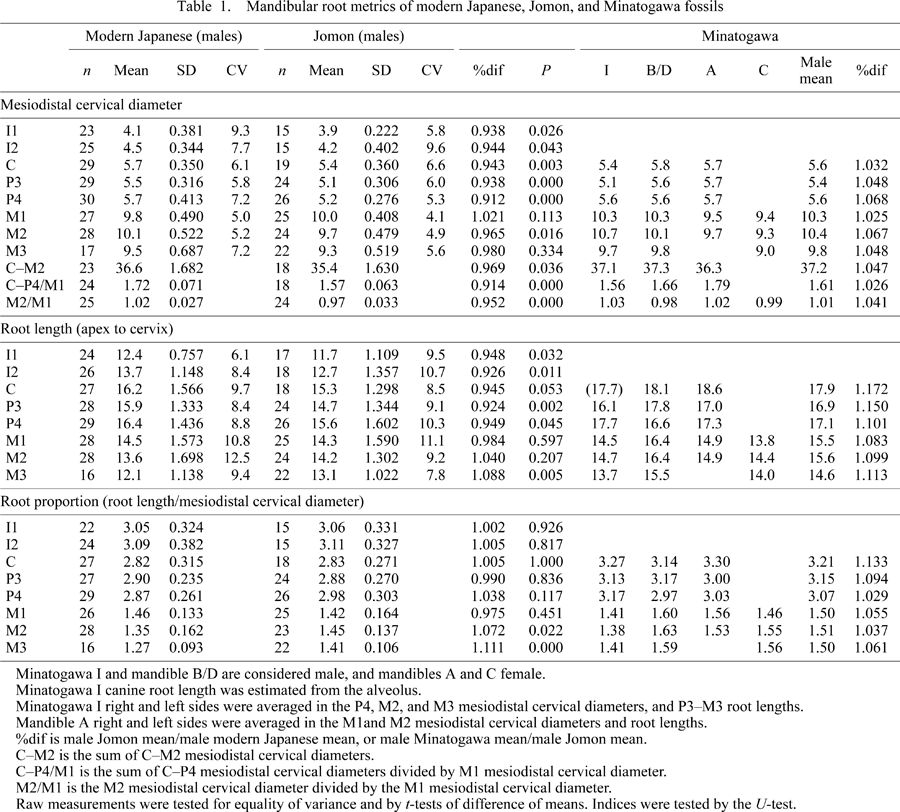
 View Details | Figure 2. Mean mandibular crown and root size in the modern Japanese and Jomon. Top: cervicoapical root lengths, mesiodistal crown lengths (middle, from Matsumura, 1995a), and mesiodistal cervical diameters; filled circles, Jomon; lines only, modern Japanese. Bottom: Jomon means divided by modern Japanese means; filled circles, mesiodistal cervical diameters; lines only, mesiodistal crown lengths (from Matsumura, 1995a); square, root lengths. |
Despite the generally smaller Jomon teeth, the mean mesiodistal cervical diameter of the M1 was slightly larger (~2%) in the Jomon than the modern Japanese. This is opposite to the condition documented for crown length by Matsumura (1994, 1995a), whose data show the modern Japanese M1 crown to be marginally larger (by <1%). However, neither mesiodistal cervical diameter nor crown length of the M1 differed significantly between the two populations. This reflects the relative conservatism of the M1, the mean crown length of which differs only slightly even in the Yayoi (only ~1% larger than the modern Japanese, Matsumura (1994, 1995a)). Thus, the entire Jomon dentition is not necessarily small, but rather, the I1–P4 and M2 are small absolutely and relative to M1 size (Table 1) (see also the univariate comparisons provided by Matsumura (1995a, 2000)).
Although heavily worn teeth are generally omitted in analyses of crown size (e.g. Brace and Nagai, 1982; Matsumura, 1994, 1995a), such criteria are variably applied, usually allowing inclusion of some degree of worn teeth to secure sample sizes. Therefore, the crown length of the M1, an early erupting tooth, would be affected (shortened) by interproximal (and occlusal) wear more so than the other teeth, especially in populations with high wear rate. In a meticulous study that aimed to ascertain the effects of this problem, Suzuki (2001) assembled crown dimension datasets of carefully chosen unworn specimens (or especially minimally worn teeth) whose dimensions were not affected by interproximal wear. His comparisons showed that the Jomon (and Yayoi) unworn molars can be larger by ~0.6 mm in mean mesiodistal crown lengths than in samples of worn teeth. Most interestingly, his wear-controlled samples show that the mean crown length of the Jomon M1 is in fact larger than that of the modern Japanese, and equivalent to the Yayoi (Suzuki et al., 2004). Results of our analysis of mesiodistal cervical diameters are concordant with the Suzuki et al. (2004) metrics, and suggest that the Jomon has a slightly larger mean M1 size than the modern Japanese.
Hillson et al. (2005) investigated the relationship of crown and cervical diameters in a sizable archeological sample of extracted unworn teeth. They reported only moderate correlations between mesiodistal crown and cervical diameters (ranging from 0.39 in the C to 0.77 in M2), but suggested that metrics alternative to the usual crown dimensions should be of use. Thus, it is of some significance to confirm that population differences in crown and cervical dimensions do actually correspond. In the present analysis, when the probable effects of dimensional reduction of the Jomon M1 crown are taken into account, whether measured by crown or cervical diameters, mean differences between the Jomon and the modern Japanese are almost identical in both magnitude and pattern along the mandibular tooth row.
The Minatogawa fossils
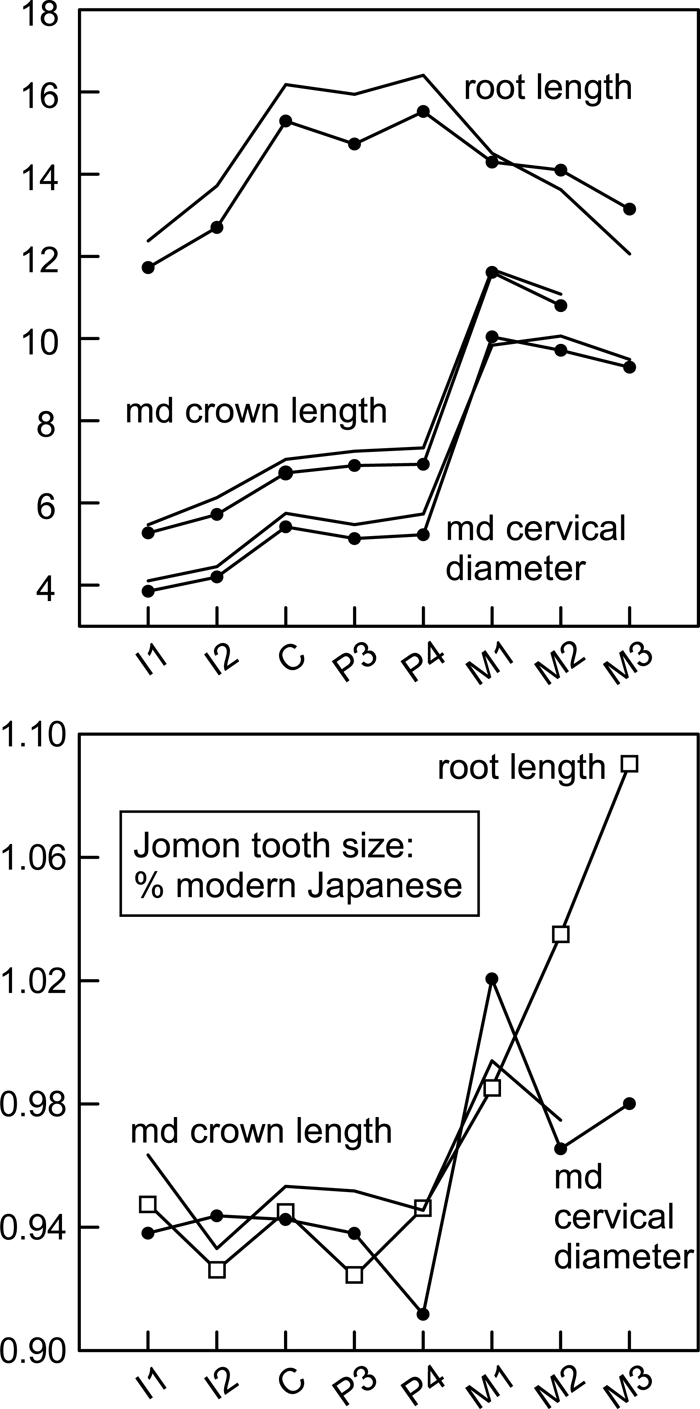
The mesiodistal cervical diameters of the Minatogawa human fossils are summarized in Table 1 and Figure 3. Evaluations are hindered by small sample sizes and possible sample bias. The mesiodistal cervical diameters of the Minatogawa male molars (M1–M3) are distinctly large. Those of the females are smaller, showing the expected pattern of sexual dimorphism (male > female). In contrast, the average canine and premolar mesiodistal cervical diameters of the two Minatogawa males are slightly less than the modern Japanese means. However, the single Minatogawa female with canine and premolars has cervical diameters larger than the Minatogawa male means. Since sexual dimorphism predicts the opposite, it is probable that the available Minatogawa males represent some degree of sample bias. Finally, the summed C–M2 mesiodistal cervical diameters of the Minatogawa individuals are large, despite their somewhat small canines and premolars, especially in the Minatogawa I individual.
 View Details | Figure 3. Mesiodistal cervical diameters of the modern Japanese, Jomon, and Minatogawa fossils. Left column: top, canine; middle, P3; bottom, M1. Right column: top, summed C–M2 mesiodistal cervical diameters; middle, summed C–P4 divided by M1 mesiodistal cervical diameters; bottom, M2 divided by M1 mesiodistal cervical diameters. Box plots show central 50% range (box), range within inner fence (vertical line) and outliers. Each cross mark represents one specimen. The Minatogawa fossils are shown with the males to the right (each individual represented by cross mark), and females to the left (each individual represented by open circle). |
It is difficult, at present, to interpret the relative sizes of the Minatogawa canines and premolars. Whereas the two males (Minatogawa I and mandible B/D) exhibit a condition more comparable to the Jomon (small canine and premolar cervical dimensions) than the modern Japanese (Figure 3), the single female (mandible A) exhibits large relative canine and premolar cervical diameters. Therefore, it is possible that the Minatogawa population as a whole had larger canine and premolar sizes than is apparent from the two known males.
Overall, we interpret the available evidence to support a characterization of the Minatogawa tooth size as large. Most likely, the Minatogawa humans combined large molars with somewhat large premolars and anterior teeth, the probable ancestral condition. However, this needs to be confirmed by future fossil finds.
Although advanced wear precludes precise mesiodistal crown measurement of the Minatogawa molars, one of us (G.S.) estimates the Minatogawa I M1 and M2 crown lengths as 12.2 and 12.0 mm, respectively (correction for interproximal wear as in White et al. (2000) and Suwa et al. (2009)). These are larger than the values reported by Hanihara and Ueda (1982) or Matsumura (2000) and approach the Australian means (Matsumura and Hudson, 2005). The large-toothed Australians are matched by the Yayoi in comparably-sized C–P4 crowns, but the Australians have larger molars (Matsumura, 1995a, b; Matsumura and Hudson, 2005). This suggests different patterns of dental reduction characteristic of population lineages. For example, the Yayoi (and modern Japanese) can be considered to represent a population lineage with somewhat reduced molars (relative to Australians), but with retention of large premolars and anterior teeth. The Jomon appears to combine a conservative (for Asian) Yayoi/modern Japanese-sized M1 and advanced reduction of all other teeth. This results in canine or premolar to M1 size ratios superficially resembling the megadont Australians (Matsumura, 1995b). Interpretations of tooth size proportions need to consider such possibilities of convergence.
In the future, it is of considerable interest to establish (1) the average molar size condition, and (2) the degree of size reduction (if any) of the premolar and anterior teeth of the Late Pleistocene Okinawa populations relative to the presumed ancestral condition, e.g. the Australo-Melanesian condition. Recently excavated skeletal materials suggest that populations with Australo-Melanesian affinities might have been distributed widely in southeastern continental/insular Asia from the Late Pleistocene to early–middle Holocene times (e.g. Matsumura and Hudson, 2005; Matsumura et al., 2008a, b; Kaifu and Fujita, 2011).
Jomon and modern Japanese
The basic statistics of root lengths (cervix to apex length) and proportions (mesiodistal cervical diameter vs. root length) are given in Table 1, and the root length means are depicted in Figure 2. From I1 through M1, differences between the modern Japanese and the Jomon parallel the situation seen in mesiodistal cervical and crown diameters. That is, mean root lengths of I1–P4 are significantly shorter in the Jomon than the modern Japanese (by 5–8%). In these teeth, root proportions (root length relative to mesiodistal cervical diameter) are, however, comparable in the two populations. In the M1, both root length and proportion do not differ significantly between the modern Japanese and the Jomon, again showing conservatism in the first molar. The two distal molars appear to exhibit a significant root length reduction in the modern Japanese, resulting in absolutely and proportionately (relative to mesiodistal cervical diameter) shorter M2 and M3 roots compared to the M1 or the Jomon conditions.
Correlation between mesiodistal cervical diameter and root length in the modern Japanese and Jomon samples was weak, with most of the significant correlations seen in the modern Japanese premolars and molars (Table 2). However, this differs from previous reports of even weaker association between root length and crown dimensions (correlation coefficients generally lower than 0.1–0.2) (Garn et al., 1978; Ozaki et al., 1988). Kupczik and Hublin (2010) measured cervical plane area and root length and found greater degrees of correlation between the two (coefficients of ~0.6) in both Neanderthal and modern human M1s (but not the M2 and M3). Furthermore, significant correlations were observed in some of the distal molars when root surface area was analyzed. This suggests some degree of morphogenetic association between cervical size and root length/shape, perhaps from structural relationship and/or selection.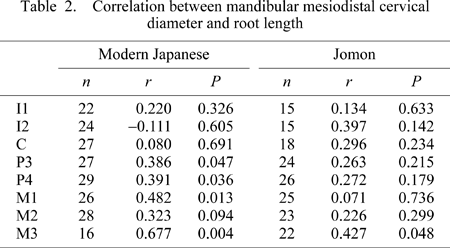
If root size patterns potentially relate to occlusal load (e.g. Spencer, 2003), the above-described modern Japanese–Jomon patterns of crown and root sizes must reflect loading conditions that are relatively consistent across populations in the M1, but not in the other teeth. Compared to the modern Japanese, the Jomon condition can be interpreted as indicating reduced load anterior to the M1 and retention of larger load potential posteriorly. Furthermore, the similar cervical diameter–root length ratios observed in the homologous teeth of the Jomon and modern Japanese (with significantly different crown sizes) suggest that occlusal load probably relates with crown size (i.e. larger teeth tend to be loaded more, or vice versa).
When root length and crown size patterns are considered together, the Jomon is characterized by isometric reduction of crown and root sizes in all tooth elements except the M1, suggesting a masticatory system with increased reliance on the first molar region. The modern Japanese pattern is characterized by a combination of retention of large premolar and canines (both crown and roots) and reduced posterior molar roots. This can be interpreted as reflecting relaxed selection for posterior molar root support, perhaps related to increasingly sophisticated food processing, while loading conditions remained proportional to crown size in the premolars and anterior teeth. These interpretations need to be cross-checked by comparable assessments across a wider range of modern human and prehistoric population samples, a data source currently not available.
The Minatogawa fossils
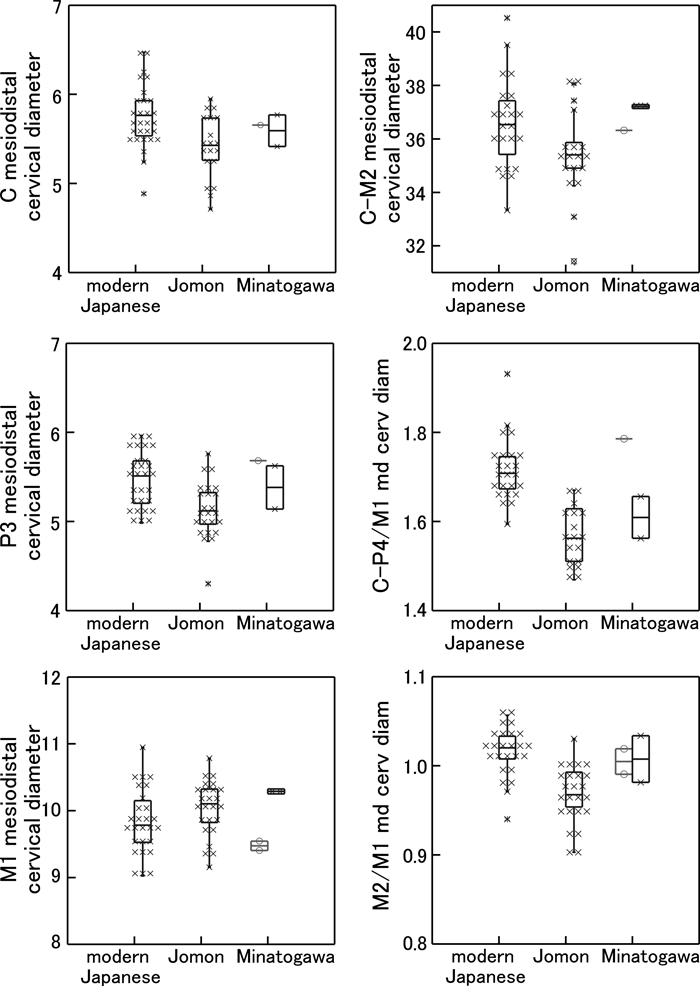
The Minatogawa teeth exhibit absolute and proportional root lengths considerably larger than the mean Jomon and modern Japanese conditions (Table 1, Figure 4). This suggests a more ‘primitive’ or conservative/ancestral masticatory apparatus with large teeth and potential for withstanding greater occlusal loads. This is in concert with the craniofacial morphologies interpreted as reflecting enhanced masticatory capacities (e.g. Suzuki, 1982; Baba and Narasaki, 1991; Baba et al., 1998; Baba, 2000).
 View Details | Figure 4. Root lengths and proportions of the modern Japanese, Jomon, and Minatogawa fossils. Left column, root length, from top to bottom: canine, P3, M1, and M3. Right column, root proportion (root length divided by mesiodistal cervical diameter), from top to bottom, canine, P3, M1, and M3. Box plots show central 50% range (box), range within inner fence (vertical line) and outliers. Each cross mark represents one specimen. The Minatogawa fossils are shown with the males to the right, and females to the left. |
A comparison of Minatogawa with other modern and prehistoric populations is of considerable interest. However, comparative root length data available in the literature is plagued by methodological inconsistency in both metric definition and measurement technique. Some of the more reliable and broadly comparable metrics are summarized in Table 3, but even these should be viewed with caution. With caliper-based measures, projected length (made by offset calipers) is expected to be systematically shorter than length measured between two points (e.g. from cervix to root apex). Radiograph-based measures are projected, but its direction is poorly controlled, and distortion factors make direct comparisons of details suspect. CT-based metrics can be taken analogous to projected measures or between two defined landmarks as in direct linear distances. In compiling Table 3, we limited the comparative samples to those with around 20 or more individuals, except for the Australian modern human and Neanderthal/Late Pleistocene samples.
With the above limitations in mind, the only modern human dataset listed in Table 3 that approaches the Minatogawa condition in terms of root length is the Australian dataset (the metric definition of which is unfortunately not available). Minatogawa’s canine and premolar root lengths are especially large, approaching the Neanderthal means. However, the Minatogawa molars tend to exhibit subequal M1 and M2 root lengths (Table 1, Figure 1), possibly a derived early modern human condition relative to Neanderthals and their contemporaries such as the Aterian H. sapiens, who appear to show distinctly longer M2 roots. Because posterior molar roots, often simpler in form, are expected to have smaller surface areas for a given root length, the Minatogawa condition of subequal M1 and M2 root lengths most likely indicates slightly reduced posterior molar loads. Such posterior root length reductions are greater in the Jomon and extreme in the modern Japanese (Figure 4).
The present study conclusively establishes the Minatogawa dentition as considerably larger in overall size than either the Jomon or the modern Japanese conditions. Furthermore, we were able to document a coherent set of distinct dentomasticatory features of the Minatogawa Late Pleistocene people: (1) absolutely and relatively long canine and premolar roots (possibly close to the basal early modern human condition); (2) a conservative molar row retaining considerably sized M1 crown and roots (possibly close to the megadont Australo-Melanesian condition); and (3) only modest degrees of posterior molar reduction as reflected by the weak M3 root length reduction. This complex of features suggests a masticatory system designed to withstand considerable load throughout the dentition, more so than in either the Jomon or the modern Japanese. When considered in light of the apparent small body size of the Minatogawa people (Baba and Endo, 1982; Baba, 2000), the dental evidence fits nicely with Minatogawa’s highly distinct facial (frontozygomatic flare) and cranial (strong infratemporal constriction, medially placed temporal line, and strong supramastoid development) morphology described by Suzuki (1982) and emphasized by Baba in relation to masticatory development (Baba et al., 1998; Baba, 2000).
There are other cranial features of the Minatogawa series that are distinct from the Jomon, most of which have been previously described (Suzuki, 1982). We have recently attempted a more thorough characterization of some of these (and some new) features including: (1) a small cranial capacity (Kubo et al., 2008; Kubo and Kono, 2011); (2) a distinct endocranial shape (low and wide proportions with a broad temporal region, and weak parietal bossing) (Kubo et al., 2011); (3) a swollen median glabellar region with obscure superciliary arches and superoinferiorly short nasal process of the frontal (Saso et al., 2011); and (4) a suite of mandibular features including an ill-developed coronoid process, small ramus breadth, and anterior alveolar prognathism (Kaifu et al., 2011). Minatogawa’s glabellonasal morphology, described in detail by Saso et al. (2011), is associated with narrow/pinched nasals (Suzuki, 1982) and lack of a steep, sagittally facing frontal process of the maxilla. The latter (strong sagittal orientation of the nasal root) is often considered a prime characteristic of the Jomon (e.g. Yamaguchi, 1996), but which is not expressed in Minatogawa. The results of the present study on tooth roots can be added to the above suite of features that make the morphological package of the Minatogawa series considerably distinct from the Jomon condition.
Although Kaifu et al. (2011), based on the mandibular evidence, suggested that the source population of the Minatogawa people probably had Australo-Melanesian morphological affinities, some other of the above summarized features, such as the endocranial (Kubo et al., 2011, Suwa et al., n.d.) and glabellar (Saso et al., 2011) morphologies are not particularly Australian-like. Multivariate evaluations of metric proportions also do not exhibit specific affinities with Australians, but show more general East or Southeast Asian similarities (Mizoguchi, 2011, and references reviewed therein). The present study of tooth roots is not clear on this issue, because of lack of relevant Australo-Melanesian and other datasets and the small Minatogawa sample size. At present, the dental evidence presented above is compatible with models that would derive Minatogawa either directly from a Late Pleistocene population with Australo-Melanesian affinities, or from a slightly evolved/differentiated Southeast to southern East Asian group of early modern humans, e.g. population groups that might also qualify as Sundadont sensu Turner (2006).
As recently presented by Hanihara and Ishida (2009) and Nakashima et al. (2010) on Jomon cranial metrics and non-metrics, Kaifu et al. (2011) on mandibular morphology, and Adachi et al. (2009) on the genetic evidence, these combined with the distinctiveness of the Minatogawa fossils make it increasingly unlikely that the Jomon had a predominantly Southeast Asian or southern Chinese origin via a Minatogawa-like ancestral population. The latter hypothesis, a southern Chinese origin of Minatogawa and the Jomon, was preferred by Suzuki (1982), while Baba inclined towards a Southeast Asian ancestry of the Minatogawa humans and Minatogawa–Jomon continuity (Baba et al., 1998). These hypotheses were based on the combination of primitive Minatogawa features and Minatogawa–Jomon (and Liujiang/Wadjak) similarities in conventional cranial metrics and selected qualitative features. Although the present study contributes only marginally to this issue, the tooth root evidence does aid in highlighting the considerable morphological gap between the Minatogawa fossils and the Jomon. For example, the modern Japanese condition, which combines a somewhat large tooth crown size with posterior molar root reduction, differs entirely from a Minatogawa to Jomon contrast. This suggests that relaxation of masticatory function (apparent in the modern Japanese) applied to Minatogawa’s dentition would not necessarily result in Jomon-like crown and root size/proportions.
Despite the above-described lack of support for a specific Minatogawa–Jomon relationship, gene flow between the geographically adjacent population lineages must have been possible, perhaps intermittently, and most likely complex in pattern. It is then probable that admixture occurred between Okinawa’s Minatogawa-derived populations and the more northerly based Jomon-culture-bearing populations. The insights gained from the present study on root size and proportions (as well as with some of the other distinctive features of the Minatogawa fossils summarized above) provide a tool with which one could test hypotheses of admixture by comparing time-equivalent Holocene population examples of the Okinawa Islands and the Jomon population of main-island Japan. Admixture would predict that the Okinawa Islands Jomon-period crania, even if exhibiting overall cranial proportions close to the Jomon condition, would show some degree of phenetic resemblance towards the Minatogawa condition in specific focal features such as in glabellonasal shape, some key mandibular features, and/or crown size and root length patterns.
We examined the mesiodistal cervical diameter and root length (cervix to apex) in modern Japanese, Holocene Jomon, and Late Pleistocene Minatogawa human fossils. We aimed to determine whether the Jomon and the modern Japanese differ significantly in root size and proportions. Our results confirmed that, in both mesiodistal cervical diameter and root lengths, the Jomon is (in most tooth elements) significantly smaller-sized, with total cervical lengths 4% smaller than in the modern Japanese. This is comparable to the known mesiodistal crown size differences between the two populations, showing that mesiodistal cervical diameters can be used in evaluating dental size. However, the cervical diameter of the first molar was not significantly different, and marginally larger in the Jomon, suggesting that dental reduction in the Jomon does not apply to the entire dentition, but only to elements other than the M1. This conclusion is supported by the root length metrics, which showed no significant difference in M1 root length between the Jomon and the modern Japanese, an isometric root length reduction (proportional to crown size) in the Jomon I1–P4, and an allometric root length reduction in the modern Japanese M2 and M3. These results reflect different patterns of dental reduction in the Jomon and the modern Japanese population lineages.
We then examined the mesiodistal cervical diameters and root lengths of Minatogawa Late Pleistocene human fossils. The Minatogawa fossils tend to have distinctly larger cervical diameters and root lengths than either the modern Japanese or the Jomon, in concordance with their cranial and facial morphology suggestive of enhanced mastication. The great root lengths of the Mintogawa fossils are possibly comparable (or close) to the large-toothed Australian condition. Their canine and premolar root lengths, furthermore, approximate the Neanderthal and apparently exceed at least some of the Late Pleistocene H. sapiens conditions. This indicates a conservative root system retained in the Minatogawa fossils, distinct from the reduced Jomon condition, providing little support for the hypothesis of a direct Minatogawa to Jomon transition. Rather, if Minatogawa-derived populations admixed with the more northerly based Jomon, this should be detectable in the Holocene of the Okinawa Islands in features such as root length patterns.
We are greatly honored to work on the Minatogawa human fossils, which were discovered in 1970 by the late Mr Seiho Oyama and a team of researchers including the late Professors Hisashi Suzuki and Naotune Watanabe of The University of Tokyo. We are indebted to their dedication and efforts in their pioneering research. We hope that our own modest studies on these fossils contribute to the continued efforts in unraveling the early population history of the Japanese archipelago. We thank Mr Seiko Oyama for research access to the Minatogawa mandibles B, C, and D, without which this study could not have been conducted. We thank Drs Osamu Kondo, Yuji Mizoguchi, and Eisaku Kanazawa for providing some of the literature that we cited. This work was supported in part by a Grant-in-Aid from the Japan Society for Promotion of Science (No. 17107006).
|