| To whom correspondence should be addressed: Takao Arai, Department of Applied Biological Science, Faculty of Science and Technology, Tokyo University of Science, 2641 Yamazaki, Noda, Chiba 278-8510, Japan. Tel: +81–4–7122–9387, Fax: +81–4–7123–9767 E-mail: takarai@rs.noda.tus.ac.jp Abbreviations: HNE, 4-hydroxy-2-nonenal; ELISA, enzyme-linked immunosorbent assay. |
Oxidative stress is associated with various pathological conditions, including inflammation, diabetes, and neurodegenerative diseases. Excessive production of reactive oxygen and nitrogen species by infiltrating phagocytic cells in pathological lesions can cause damage to the cell membrane, initiating a chain reaction of lipid peroxidation and accumulation of lipid peroxides (Niki et al., 2005). Once formed, lipid peroxides are either reduced to their corresponding alcohols by cellular glutathione peroxidases, or undergo spontaneous decomposition yielding a number of aldehydic end products (Esterbauer et al., 1991; Uchida, 2003). 4-Hydroxynonenal (HNE) is one of the breakdown products of lipid peroxides generated from ω-6 polyunsaturated fatty acids such as arachidonic acid. HNE is an α,β-unsaturated alkenal and can react with various cellular nucleophiles, including cysteine, histidine, and lysine in proteins, and guanine in DNA molecules, forming Michael adducts (Esterbauer et al., 1991; Uchida, 2003). The toxicity of HNE has been ascribed to the covalent modification of vital cellular proteins, including Na+-K+ ATPase (Mark et al., 1997; Siems et al., 1996) and mitochondrial enzymes (Picklo et al., 1999) as well as DNA (Hu et al., 2002). Elevated levels of HNE production have been reported in several neurodegenerative diseases, such as Alzheimer’s and Parkinson’s diseases (Lovell et al., 1997; Sayre et al., 1997; Selley, 1998), suggesting a role in the pathogenesis of the diseases.
Microtubules play an important role in the physiological functions of the cell, ranging from the maintenance of cell morphology to cell proliferation and differentiation (Howard and Hyman, 2003). Microtubules are polymers of tubulin heterodimers composed of α and β subunits, and microtubule-associated proteins. In the cytoplasm, microtubules are in a state of dynamic equilibrium between polymerization and depolymerization of these components. Tubulin has 20 cysteine and 24 histidine residues per dimer. Studies on residue-selective alkylation or oxidative modification have demonstrated that some of these residues are important for polymerization (Hosono et al., 2005; Kuriyama and Sakai, 1974; Mellon and Rebhun, 1976; Oliver et al., 1976; Rai and Wolff, 1998).
Previous studies have demonstrated that exposure of cells to HNE disrupts the microtubule structure, and that this is followed by the death, hours later, of various types of cells such as fibroblasts, myocytes, and neurons (Gadoni et al., 1993; Neely et al., 1999; Olivero et al., 1990; VanWinkle et al., 1994). In vitro studies with purified microtubule fractions have shown that the modifications of tubulin by HNE impair the polymerizability (Neely et al., 2005; Olivero et al., 1990). Nevertheless, the precise mechanisms underlying the microtubule disappearance in cell caused by HNE exposure remain to be fully clarified. In this study, we addressed the mechanism both in vitro and in vivo. We show here using a couple of types of cells and a rat brain microtubule fraction that α-tubulin is a common major target modified by HNE. Moreover, our in vitro study demonstrates that HNE can modify unpolymerized tubulin and impair its polymerization but has a marginal effect on the integrity of assembled microtubules. Taken together, we propose that the disappearance of microtubule structures in cells exposed to HNE results from disruption of new microtubule assembly rather than destruction of assembled microtubules.
2-(N-morpholino)ethanesulfonic acid (MES), piperazine-1,4-bis (2-ethanesulfonic acid) (PIPES), and ethylene glycol-bis(2-aminoethylether)-N,N,N,N-tetraacetic acid (EGTA) were purchased from Dojindo (Kumamoto, Japan). HNE was purchased from Cayman Chemical Co. (Ann Arbor, MI). Triton X-100 was obtained from Sigma-Aldrich (St Louis, MO). Other chemicals, salts, etc. used were of analytical grade.
Monoclonal antibodies against α-tubulin (E3B8) (Arai and Matsumoto, 1988) and β-tubulin (E1) (Ohuchi, 2000) were produced in our laboratory. A monoclonal anti-HNE antibody (11B11) was established using HNE-modified rat microtubules as an antigen according to a procedure reported previously (Arai and Matsumoto, 1988). The antibody also reacted with HNE-modified BSA, indicating that the epitope is an HNE moiety (data not shown).
Primary cerebral neurons were isolated from the brains of 17-day-old rat fetuses according to a procedure described by Suzumura et al. (Suzumura et al., 1996). Briefly, brain cells were dissociated from fetal brain by pipetting in DF medium (DMEM F-12 HAM, 15.8 mM glucose, 25 μg/ml insulin, 2 μM progesterone, 9.6 mg/ml ptoresin, 0.03 μM sodium selenite, 0.1 mg/ml apotransferrin, 100 U/ml penicillin G, 100 μg/ml streptomycin, 5 mM HEPES) and plated in polylysine-coated culture dishes at a density of 1.5×105 cells/cm2. The cells were cultured in 5% CO2 at 37°C for one week and used for experiments. Rat fibroblastoma 3Y1 cells were maintained in Dulbecco’s Modified Eagle Medium supplemented with 10% FCS, 100 U/ml penicillin, and 100 μg/ml streptomycin under a humidified atmosphere of 5% CO2 (Ohuchi, 2000). For experiments with HNE, cells were plated in polylysine-coated 96-well plates, 8-well plates, or 60-mm dishes (BD Biosciences, San Jose, CA) at a density of 6.25×103 cells/cm2 48 h prior to the assays.
Total cell lysates were obtained by sonication of the cells in PBS containing 1% Triton X-100, 1 mM phenylmethanesulfonyl fluoride, 10 μg/ml leupeptin, and 10 μg/ml pepstatin.
The cell lysates and microtubule proteins were resolved by SDS-PAGE (4 M urea-8% gel), transferred to polyvinylidene difluoride membranes (Immobilon). Tubulin and HNE-modified proteins were detected by their respective monoclonal antibodies, followed by horseradish peroxidase-labeled secondary antibody and chemiluminescence visualization using ECL Plus (GE Healthcare).
A 96-well microplate was coated with anti-β-tubulin antibody (E1) and blocked with BSA. Following sample application, HNE moieties on the bound tubulin were detected with biotin-labeled anti-HNE antibody (11B11), followed by horseradish peroxidase-coupled streptavidin. For color development, we used o-phenylenediamine, and absorbance was read on a microplate reader at 450 nm.
Cells were cultured on eight-well culture slides (BD Biosciences) at a density of 6.25×103 cells/cm2. Following HNE treatment, cells were washed with PBS and fixed with 4% paraformaldehyde in PBS. Tubulin and HNE-modified proteins were detected by their respective monoclonal antibodies followed by Alexa Fluor 488-labeled secondary antibodies. Nuclei and actin fibers were visualized with Hoechst 33258 and phalloidin-rhodamine, respectively, according to the instructions supplied by the respective manufacturers. Images were acquired using a fluorescence microscope (BXC51, Olympus, Japan) equipped with a computer image analyzer (SIT camera C2400 and Argus 50; Hamamatsu Photonics Co., Japan).
Microtubules were prepared from rat brains by repeated assembly-disassembly cycles according to the procedure of Shelanski et al. (Shelanski et al., 1973) with some modifications (Arai, 1983; Arai and Kaziro, 1977). Microtubule fractions (2 mg/ml) in MES assay buffer (0.1 M MES, 1 mM EGTA, 0.75 mM MgCl2, 50 μM CaCl2, 25% glycerol, and 1 mM GTP; pH 6.8) were incubated with HNE on ice for 1 h, and unreacted HNE was quenched by adding 1 mM N-acetylcysteine. Polymerization was induced by warming the fractions at 37°C for 1 h, and the assembled microtubules were collected by centrifugation at 60,000×g for 1 h at 25°C. Because some of the microtubules became insoluble as a result of HNE modification, the microtubule precipitants were disassembled on ice for 1 h. The intact, disassembled microtubule proteins and insoluble materials were then separated by centrifugation at 30,000 × g for 1 h at 4°C. The protein concentration in the supernatant was used as an index of polymerization.
In some experiment, microtubule proteins were pre-treated with HNE in the MES assembly buffer without GTP at 37°C for 1 h in a 96-well plate with a total volume of 150 μl, and polymerization was carried out by adding 1 mM GTP, and the turbidity was monitored on a plate reader (SpectraMax, Molecular Devices) at 37°C.
Cell viability was measured by using Cell Counting Kit-8 (Dojindo) according to the manufacturer’s instructions. Protein concentrations were determined by the Bradford method using BSA as a standard (Bradford, 1976).
Rat primary neuronal cells were incubated with increasing concentrations of HNE for 1 h, and HNE-modified proteins were analyzed by Western blotting using anti-HNE antibodies. As shown in Fig. 1A, the levels of HNE-modified proteins increased in a concentration-dependent manner up to at least 125 μM. Two proteins with molecular masses of 50 and 55 kDa were labeled by HNE even at concentrations as low as 30 μM, the 55 kDa species being more intensely labeled than the 50 kDa. Above 60 μM, several proteins of higher molecular weight (100–250 kDa) were also labeled by HNE. The predominant HNE targets, the 55 and 50 kDa bands, coincided with the same migration positions as those of α- and β-tubulins, respectively, as revealed by anti-α- and anti-β-tubulin monoclonal antibodies on the same blot (Fig. 1A). The results suggest that tubulin is one of the major targets of HNE in primary neuronal cells.
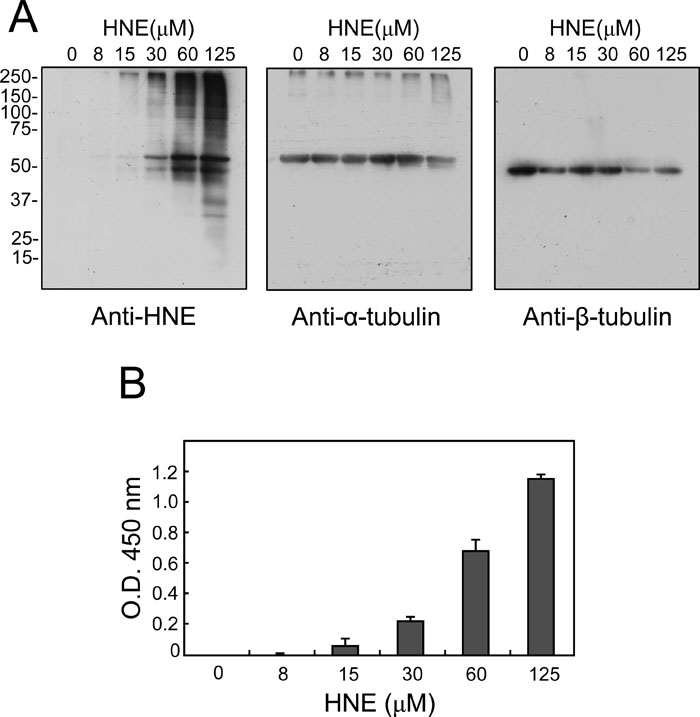 View Details | Fig. 1. Immunochemical detection of HNE-modified proteins in HNE-treated rat primary neuronal cells. Primary neuronal cells were treated with the indicated concentrations of HNE for 1 h, and HNE-protein adducts were detected by immunoblotting with antibody to HNE (A). The same membranes were also probed with anti-α-tubulin antibody and anti-β-tubulin antibody as noted. In B, the levels of HNE-tubulin adducts in the total cell extracts from HNE-treated cells were measured by sandwich ELISA. Values are mean±SD (n=3). |
To ensure that tubulin was indeed modified by HNE, the cell lysates were also analyzed by sandwich ELISA, in which anti-β-tubulin and anti-HNE antibodies were used as the capturing and detecting antibodies, respectively (Fig. 1B). Since tubulin exists as a stable heterodimer of α- and β-subunits, the ELISA was expected to detect NHE-modified tubulin regardless of the subunit being modified. HNE-modified tubulin could be detected at levels as low as 15 μM, and the levels increased with HNE concentration.
To find out whether tubulin modification by HNE could also be observed in other cells, we used rat neuron-like PC12 cells and fibroblast 3Y1 cells. Immunoblot analysis of HNE-modified proteins in PC12 cells (Fig. 2A) and 3Y1 cells (Fig. 2B) revealed that HNE also preferentially targeted α-tubulin; In either cells, HNE-modified α-tubulin appeared at an HNE concentration of 30 μM, and HNE-modified β-tubulin at 125 μM. Sandwich ELISA also verified the HNE modification of tubulin in this cell line (Fig. 2A and B). Thus, it is concluded that HNE can modify tubulin, preferentially α-tubulin, in primary neurons, PC12 cells and 3Y1 cells.
 View Details | Fig. 2. Immunochemical detection of HNE-modified proteins in HNE-treated PC12 cells and 3Y1 cells. As in the legend to Fig. 1, PC12 cells (A) or 3Y1 cells (B) were treated with the indicated concentrations of HNE for 1 h, and HNE-protein adducts were detected by immunoblotting with antibody to HNE. The positions of α- and β-tubulin are shown by arrowheads. The band around 65 kDa is an unknown protein. The levels of HNE-tubulin adducts in the total cell extracts from HNE-treated cells were measured by sandwich ELISA. Values are mean±SD (n=3). |
To assess the effect of HNE modification on microtubule integrity in cells, we next conducted immunofluorescence analyses. Since immunocytochemical visualization of the microtubule networks of the primary neuronal cells and PC12 cells was technically difficult, we used 3Y1 cells. 3Y1 cells were exposed to increasing concentrations of HNE for 1 h, and microtubule structures were stained with anti-β-tubulin antibody. Essentially, no alteration could be observed in cells exposed to HNE at concentrations below 15 μM (data not shown). When cells were exposed to HNE at 30 μM, however, the cells assumed a round shape, and the filamentous microtubule structures were shrunken and reduced in length (Fig. 3B). The effect of HNE at 30 μM was comparable to that of a low concentration (100 nM) of nocodazole, an inhibitor of microtubule polymerization (data not shown). The microtubule alterations with this HNE concentration proceeded progressively, and the filamentous structures disappeared almost completely by 12 h after treatment (data not shown). Exposure to 60 μM HNE for 1 h resulted in more severe microtubule destruction with cell shrinkage (Fig. 3C). With 125 μM HNE, the microtubule structures vanished completely within 1 h (data not shown). The effect of HNE at this concentration on microtubules was comparable to that of a high concentration (10 μM) of nocodazole (data not shown). These acute effects of HNE on microtubule filamentous structures were consistent with a report by Neely et al., who used Neuro 2A cells (Neely et al., 1999).
 View Details | Fig. 3. Immunofluorescence microscopic analysis of microtubule structures after treatment with HNE in 3Y1 cells. 3Y1 cells were untreated (A, D, G) or treated with HNE at 30 μM (B, E, H) or 60 μM (C, F, I) for 1 h, and were double stained with anti-β-tubulin antibody for visualization of microtubule structures (A–C) and with phalloidin for actin fibers (D–F). In parallel, HNE-modified proteins were also visualized by anti-HNE antibody using another set of cells (G–I). Bar=50 μm. |
The HNE treatment was also associated with the alterations of the actin fibers (Fig. 3E and F). Notably, immunostaining of the cells with anti-HNE antibody revealed that HNE-modified components did not co-localize with filamentous microtubule structures but were distributed throughout the cytoplasm and occasionally the nucleus (Fig. 3H and I).
To elucidate the effect of HNE modification at the protein level, purified rat brain microtubule proteins were modified with HNE in the presence of GTP at 0°C, and their polymerizability at 37°C was evaluated. A preliminary study revealed that significant portions of HNE-modified microtubule proteins become insoluble (data not shown). Therefore, polymerized microtubules were first collected by centrifugation, then depolymerized on ice, and finally centrifuged again to separate insolubilized materials into the pellet and depolymerizable active microtubule proteins into the supernatant. The degree of microtubule polymerization was expressed as the amount of protein in the supernatant.
HNE treatment concentration-dependently impaired microtubule polymerization upon increasing the temperature to 37°C in the presence of GTP (Fig. 4A). Approximately 300 μM HNE was required to inhibit microtubule polymerization by 50%, and nearly 800 μM was needed for maximum inhibition. Coomassie staining of SDS-PAGE gels for the two fractions demonstrated that with increasing HNE concentrations both α- and β-tubulins in the supernatant (assembled microtubules) disappeared proportionally, and concomitantly a novel broad band with an apparent molecular mass of 50 kDa in the insoluble pellet (denatured microtubule proteins) increased (Fig. 4B). Notably, the increase in the level of the 50 kDa band was reciprocally correlated with the disappearance of 55 kDa α-tubulin in the insoluble fraction. There was no apparent change in the mobility of 50 kDa β-tubulin on SDS-PAGE after treatment with HNE (Fig. 4B). Immunoblot analysis of the same samples revealed that most of the HNE-modified tubulin was recovered in the insoluble fraction at the same position as the 50 kDa band and β-tubulin. These results demonstrate that tubulin lost its polymerizability on HNE modification and that modified α-tubulin migrated faster and appeared as a 50 kDa band at the same position as β-tubulin on SDS-PAGE.
 View Details | Fig. 4. Effect of HNE on microtubule polymerization. A rat brain microtubule fraction (2 mg/ml) was treated with increasing concentrations of HNE for 1 h on ice, and then polymerized at 37°C. Polymerizable and denatured tubulins were separated by centrifugation as described in Materials and Methods. In A, microtubule polymerizability was expressed as a function of HNE concentration. Values are mean±SD (n=4). In B, polymerized (microtubules) and denatured tubulins (denatured) were subjected to SDS-PAGE followed by Coomassie staining (above) or anti-HNE immunoblotting (below). In (C) and (D), the microtubule proteins (1.25 mg/ml) in the MES assembly buffer without GTP was treated with HNE for 1 h at 37°C, and then polymerized by GTP. The turbidity was recorded on a plate reader, and polymerized (microtubules) and denatured tubulins (denatured) were analyzed for HNE adducts as above. |
Although significant parts of microtubules were destructed by HNE in live cells even at concentrations below 60 μM (Fig. 3), relatively high concentrations were required to observe similar destructive effects in the purified microtubule proteins (IC50=300 μM; Fig. 4A). As the differences in reaction temperature of HNE treatments, 37°C vs. 0°C, were considered as a factor responsible for the different effective concentration ranges of HNE between cells and cell-free assays, microtubule proteins were treated with HNE at 37°C in the absence of GTP, and then induced polymerization by adding GTP. In a turbidity assay, the HNE treatment attenuated microtubule formation at a concentration as low as 6 μM and in a concentration-dependent manner at least up to 50 μM (Fig. 4C). However, at 100 μM, the turbidity was apparently increased. Fractionation of intact and denatured microtubule proteins followed by SDS-PAGE analysis confirmed the concentration-dependent inhibition of microtubule assembly, which was associated with proportional increases in HNE-modified denatured tubulin (Fig. 4D). Immunoblotting analysis also revealed that small portions of HNE-modified tubulin can still be incorporated in microtubules, and that the bands for HNE-modified tubulin were completely superimposable to those for α-tubulin (data not shown). Thus, α-tubulin is also a selective target of HNE in cell-free conditions.
Next, we measured the effect of HNE on the stability of preassembled microtubules. GTP-triggered polymerized microtubules were incubated with various concentrations of HNE for 1 h at 37°C, and the amount of tubulins in the soluble and insoluble fractions was assessed as above. In contrast to the effect of HNE on polymerization, the degree of HNE-dependent disassembly of polymerized microtubules was less than 30% of the control even at 1000 μM, as judged by the protein concentration of the supernatant (Fig. 5A). In Coomassie staining, HNE-modified tubulin also appeared as a 50 kDa band and was recovered exclusively from the insoluble fraction (Fig. 5B). However, the levels were far less than those of unpolymerized microtubule proteins (compare with Fig. 4B). Similar results were obtained in the turbidity assay (data not shown). The results indicate that the assembled microtubules were less susceptible to HNE modification than free tubulin, and that HNE modification renders microtubules insoluble.
 View Details | Fig. 5. Effect of HNE on polymerized microtubules. A rat brain microtubule fraction (2 mg/ml) was polymerized at 37°C and then treated with the indicated concentrations of HNE for 1 h at 37°C. Polymerizable and denatured tubulins were separated by centrifugation as described in Materials and Methods. In A, levels of polymerized microtubules were expressed as a function of HNE concentration. Values are mean±SD (n=4). In B, polymerized (microtubules) and denatured tubulins (denatured) were subjected to SDS-PAGE followed by Coomassie staining (above) or anti-HNE immunoblotting (below). |
A number of target molecules and mechanisms have been suggested to account for the cytotoxicity of HNE (Gadoni et al., 1993; Hu et al., 2002; Mark et al., 1997; Neely et al., 2005; Picklo et al., 1999; Siems et al., 1996). The present study revealed that tubulin, preferentially α-tubulin, undergoes HNE modification in 3Y1 cells, PC12 cells, and primary neurons, although the relative sensitivity to modification of α-tubulin to β-tubulin by HNE differed slightly among cell types (Fig. 1 and 2). A previous study by Neely et al. showed that α-tubulin is one of the major targets of HNE in Neuro 2 neuroblastoma cells (Neely et al., 1999). Our results, in conjunction with theirs, indicate that α-tubulin is a common major target of HNE in various cells.
Morphologically, microtubule disruption has been reported to be associated with HNE toxicity in various cells (Gadoni et al., 1993; Neely et al., 1999; Olivero et al., 1990; VanWinkle et al., 1994). In agreement with the previous results and consistent with our data on tubulin modification, exposure of 3Y1 cells to HNE (>30 μM) caused rapid (<1 h) and irreversible destruction of microtubule structures (Fig. 3). The HNE concentrations employed in these experiments were cytotoxic to the cells; at 30 μM approximately 50% of cells were killed within 12 h, and at 60 and 125 μM HNE almost all the cells were killed within 6 and 3 h, respectively (data not shown). Although actin filaments were also destroyed by HNE treatment (Fig. 3), no protein corresponding β-actin (45 kDa) was detected as HNE adducts in immunoblotting (Fig. 1 and 2), suggesting the collapse of the actin filaments was presumably through an indirect mechanism such as a consequence of the microtubule disruption. The relatively selective modifications of α-tubulin and the disappearance of microtubule structures by HNE treatment observed in these cells prompted us to elucidate the mechanism at the protein level.
In a rat brain microtubule fraction, HNE impaired microtubule assembly and covalently modified unpolymerized tubulin in a similar concentration range in which microtubule disappearance was observed in cells. The HNE modifications, which are also selective for α-tubulin at least at low HNE concentrations, caused insolubilization of both α- and β-tubulins to a similar extent (Fig. 4). Since tubulin exists as a α- and β-tubulin heterodimer, it may be possible that β-tubulin may have been precipitated without HNE attachment as a result of the insolubilization of HNE-modified α-tubulin. It has been demonstrated that a thiol-blocking reagent not only abrogates the polymerizability of free tubulin but also can destroy assembled microtubules (Kuriyama and Sakai, 1974). Surprisingly, however, HNE could not destroy assembled microtubules to such a degree, and the extent of modification was far less than that for unpolymerized microtubule proteins (Fig. 5). A recent report by Stewart et al. also demonstrated in a turbidity assay that polymerized tubulin was more resistant than unpolymerized molecules to modification by HNE (Stewart et al., 2007). At least three cysteine residues on or near the surface of the tubulin dimer have been identified as targets of HNE (Stewart et al., 2007). The difference in susceptibility to HNE between polymerized and unpolymerized tubulins may be attributable to the helical ring structure of microtubules. That is, the target amino acid(s) of HNE in the tubulin molecule may be faced inward the tubular structure of microtubules or buried between adjacent polymerized tubulin molecules, thereby being protected from reaction with HNE.
In purified rat brain microtubule fractions, HNE-modified α-tubulin migrated faster than its unmodified counterpart and appeared at the same position as β-tubulin (Figs. 4 and 5). The faster mobility of α-tubulin suggests that it may have been intramolecularly cross-linked and adopts a globular shape because HNE can act as a divalent cross-linking reagent (Uchida and Stadtman, 1993). Alternatively, this may be attributable to an intrinsic property of α-tubulin upon Cys modification; palmitoylation of Cys residues of tubulin dimers, which does not result in a cross-link, also causes selective mobility shift of α-tubulin to the same position as β-tubulin (Wolff et al., 2000).
Meanwhile, contrary to the result of the cell-free study, HNE modification of 3Y1 or primary neuronal cells did not affect the mobility of α-tubulin on SDS-PAGE. The difference in the mobility of HNE-modified α-tubulin on SDS-PAGE between the cell and cell-free systems may have been due to the difference in pH; a slightly acidic pH (~6.5) used for in vitro assays may favor intramolecular cross-linking via Schiff base formation over hemiacetal formation, compared with reactions at the cytosolic pH of 7.4. Alternatively, a possible greater extent of cross-linking of α-tubulin as a result of the higher HNE concentrations used in the cell-free system may be responsible for the difference. In-depth biochemical analyses for these issues are under way.
Based on the in vitro results, it is considered that HNE can modify mostly unpolymerized α-tubulin but not polymerized microtubules in intact cells as well. Consistent with the interpretation, HNE treatment of 3Y1 cells resulted in scattered HNE immunoreactivity throughout the cells without co-localization with the remaining microtubule filamentous networks (Fig. 3), suggesting HNE modify only unpolymerized tubulin. Given cellular microtubules are in a state of dynamic equilibrium between polymerization and depolymerization, the disappearance of microtubule structures by HNE in intact cells could result from the inhibition of polymerization of tubulin rather than destruction of polymerized microtubules. Increased HNE levels have been reported in several neurodegenerative disorders, such as Alzheimer’s and Parkinson’s diseases (Lovell et al., 1997; Sayre et al., 1997; Selley, 1998). The present results suggest that impaired tubulin polymerization caused by HNE may be in part responsible for the decline in neuronal activity in the early stages of these diseases.
In conclusion, the present study has demonstrated that modification of unpolymerized tubulins, predominantly α-tubulin, by HNE, which results in insolubilization and polymerization incompetence, is responsible for the rapid and irreversible disappearance of microtubule structures in cells exposed to HNE.
This work was supported by grants from the Promotion and Mutual Aid Corporation for Private Schools of Japan, and Research on Advanced Medical Technology from the Ministry of Health, Labor, and Welfare of Japan.
|