| To whom correspondence should be addressed: Kohji Takei or Kenji Tanabe, Department of Neuroscience, Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences, 2-5-1, Shikata-cho, Kita-ku, Okayama 700-8558, Japan. Tel: +81–86–235–7125, Fax: +81–86–235–7126 E-mail: kohji@md.okayama-u.ac.jp (K. Takei) ktanabe@md.okayama-u.ac.jp (K. Tanabe) Nobuhisa Ishida, Yuichi Nakamura, and Kenji Tanabe contributed equally to this work. Abbreviations: BSA, bovine serum albumin; DAPI, 4',6-diamino-2-phenylindole; DIC, differential interference contrast; GED, GTPase-effector domain; GFP, green fluorescence protein; PB, phosphate buffer; PRD, proline-rich domain; SH3, Src-homology domain-3. |
Dynamin is known as a key player of clathrin-mediated endocytosis, and the majority of studies have sought to reveal its molecular mechanism (Kruchten and McNiven, 2006; Praefcke and McMahon, 2004). Dynamin polymerizes around the neck of newly formed endocytic pits and segregates them from the donor membrane by changing its conformation. There are three dynamin genes in humans: brain-specific dynamin 1, ubiquitously-expressed dynamin 2, and brain- and testis-enriched dynamin 3. Each gene has several splicing variants, and they each show distinct cellular localizations (Cao et al., 1998). Dynamin comprises several functional domains including a GTPase domain, middle domain, GTPase effector domain (GED), PH domain, and proline-rich domain (PRD). The middle domain binds to γ-tubulin, and is required for cohesion of centrosomes (Thompson et al., 2004). The PRD interacts with SH3 domain of amphiphysin, endophilin, syndapin, and others, and stimulates the GTPase activity to induce dynamin’s conformational changes, which are required for membrane fission (Cao et al., 1998; Qualmann et al., 2000; Witke et al., 1998).
Although dynamin is known as a fission protein for membrane transport, it was originally identified as a microtubule-binding protein (Shpetner and Vallee, 1989). Recently, we identified dynamin as a regulator of dynamic instability of microtubules, and showed that this function was impaired in a certain form of neuropathy (Tanabe and Takei, 2009). Dynamin has other functions independent of its membrane fission activity, including the formation of centrosomes (Thompson et al., 2004) and actin reorganization (Schafer et al., 2002), although these molecular mechanisms are still unclear.
Both microtubule dynamics and membrane transport are required for signal transduction, bacterial/virus infection, immunoresponse and so on (Burgdorf et al., 2007; Burgdorf and Kurts, 2008; Greene and Gao, 2009; Miaczynska et al., 2004; Miyauchi et al., 2009; Sadowski et al., 2009; Watanabe et al., 2005). Of these, cell cycle progression also requires both microtubule dynamics and membrane transport for the entry into mitosis, segregation of chromosomes, alteration of cell shape, and abscission between daughter cells. Both microtubules and actin filaments play central roles in mitosis (Doxsey et al., 2005; Glotzer, 2003; Peris et al., 2006; Rieder and Cole, 2000; Zimmerman et al., 2004). Membrane traffic associated proteins are also indispensable for cytokinesis, probably through membrane deformation for abscission of daughter cells (Conner and Wessel, 2000; Schweitzer et al., 2005; Schweitzer and D’Souza-Schorey, 2005; Strickland and Burgess, 2004). Previously, dynamin was found to be required for cell cycle progression, especially during cytokinesis (Konopka et al., 2006; Thompson et al., 2002; Wienke et al., 1999). On the other hand, clathrin, a coat protein for transport vesicles, was localized on the mitotic spindle, and required for the formation of the mitotic spindle (Royle et al., 2005). Thus, components required for membrane traffic may have other roles for cell cycle progression independent of membrane traffic.
As dynamin regulates microtubule dynamics (Tanabe and Takei, 2009) and cell cycle progression requires dynamic instability of microtubules (D’Avino et al., 2005; Galjart, 2005), we investigated the role of dynamin at cell cycle progression, especially focused on its interaction with microtubules. We found that dynamin was localized on the mitotic spindle, and depended on its PRD to do so. The deletion of PRD resulted in the impairment of both the localization and the abscission of daughter cells. Interestingly, this deletion mutant had no significant effect on endocytic function regulated by dynamin. On the other hand, another mutant of dynamin, which blocks dynamin-dependent endocytosis, also had a profound effect on abscission, but no alteration on the localization of the midzone. These results suggest that dynamin may regulate cytokinesis through both membrane traffic and microtubule dynamics.
HeLa cells were maintained in DMEM containing 10% FBS, penicillin-streptomycin, and fungizone at 37°C, 5% CO2. Antibodies were as follows: goat antibody against dynamin 2 (C-18, Santa Cruz), mouse antibodies against α-tubulin and γ-tubulin (Sigma), rabbit antibody against V5 (Millipore). Secondary antibodies were as follows: Cy3-conjugated donkey anti-goat IgG (KPL), Alexa488-conjugated goat anti-mouse IgG and Rhodamine Red-X-conjugated goat anti-rabbit IgG (Invitrogen). Alexa488-conjugated human transferrin was from Invitrogen.
Rat dynamin 2aa wild type and K44A fused to GFP were generous gifts from M. A. McNiven (Mayo Clinic, Rochester, MN, USA). Dynamin 2 wild type and K44A tagged with V5/His were created as previously described (Tanabe and Takei, 2009). Deletion constructs were made by PCR, checked by sequencing, and inserted into pcDNA4-V5/His (Invitrogen). ΔPRD and Δmiddle constructs contain 1–740 aa, and 1–300 aa conjugated to 526–870 aa, respectively. DNAs were purified for transfection experiments by using a Plasmid Maxi Kit (QIAGEN).
Cells were synchronized at the G1/S boundary by thymidine-hydroxyurea block, as described by Krek and DeCaprio (Krek and DeCaprio, 1995). Briefly, cells were plated onto multiple 100 mm dishes and culture in the presence of thymidine (2.5 mM; Sigma) for 24 h. Then, they were washed with normal growth medium, incubated for an additional 10 h under normal growth conditions, and finally arrested by the addition of hydroxyurea (1 mM; Wako) for 16 h. At time zero, cells were washed three times with phosphate buffered saline (PBS) and placed under normal growth conditions. Mitotic cells were collected and analyzed at 12 h.
Cells were fixed with 4% paraformaldehyde in PBS for 15 min at RT, washed three times by PBS (Tanabe and Takei, 2009). Endogenous dynamin 2 was detected by anti-dynamin 2 antibody, and exogenous dynamin 2 was detected by anti-V5 antibody. For cytosol extraction, cells were incubated in 1% TX100, BRB80 {80 mM PIPES/KOH (pH 6.8), 4% PEG8000 (W/V), 1 mM MgCl2, 1 mM EGTA} for 5 min at 37°C, and washed by BRB80 without TX100 and fixed as described above (Tanabe and Takei, 2009). For drug treatment, cells were treated with nocodazole (1 μg/ml) for 30 min, or 2.5 μM Taxol (Sigma) overnight. For internalization assay, cells were incubated with 5 μg/ml rhodamine-conjugated transferrin for 30 min, and then fixed and processed for immunofluorescence. Cells were processed as previously described (Tanabe and Takei, 2009), and analyzed by the confocal spinning disk microscope system (CSU10; Yokogawa Electric Corp.) on an inverted microscope (IX-71; Olympus) equipped with an Ar/Kr laser. Images were acquired using a UPlan-Apochromat 100 NA 1.35 oil immersion objective (Olympus) and an electron-multiplying charge-coupled device camera (iXon; Andor Technology). Image capturing and acquisition were performed using MetaMorph software (MDS Analytical Technologies).
Cells were plated onto glass-bottomed culture dishes (Iwaki), transfected with GFP-dynamin 2 and synchronized as described above. The cells were incubated in culture medium and kept at 37°C and 5% CO2 using a stage incubator (MI-IBC; Olympus) and digital gas mixer (GM-2000; Tokai Hit) and imaged using the aforementioned confocal system. Images were taken every 5 min for 1 h.
Synchronized HeLa cells were permeabilized with 1% TX100 to extract the cytosolic proteins, then fixed with 4% paraformaldehyde, 0.05% glutaraldehyde and 4% sucrose in 0.1 M phosphate buffer (PB), pH 7.4. Fixed cells were scraped and centrifuged to make cell pellet. After washing the pellet with 0.1 M PB three times, the samples were dehydrated with ethanol, then embedded in LR White (London Resin Company Ltd., England). Ultrathin sections picked up on nickel grids were incubated with 6 M urea in 0.1 M glycine-HCl buffer (pH 3.5) for 5 min to etch the surface of sections. The grids were washed three times with 0.1 M PB containing 0.1% bovine serum albumin (BSA), and then blocked for 15 min with 10% normal rabbit or goat serum and 1% BSA in 0.1 M PB. The grids were incubated with primary antibodies at 4°C for overnight. After washing with 0.1% BSA in 0.1 M PB five times, the grids were incubated for 1.5 h with the secondary antibodies conjugated with 10-nm colloidal gold. The grids were washed with 0.1% BSA in 0.1 M PB, postfixed with 2% glutaraldehyde, rinsed with H2O, and dried. The sections were stained with 2% uranyl acetate and 3% lead citrate, and observed with a Hitachi H-7100 transmission electron microscope at the Central Research Laboratory of Okayama University Medical School.
In order to examine the localization of dynamin 2 during mitosis, cell cycle-synchronized HeLa cells were double stained by immunofluorescence for dynamin 2 and α-tubulin. Dynamin immunoreactivity, which was observed as fine puncta present throughout the cytoplasm at interphase (Fig. 1A), was evident at centrosomes as the cell entered prophase (Fig. 1B). During prometaphase and metaphase, dynamin was concentrated in the proximity of kinetochore microtubules as well as polar microtubules (Fig. 1C and D). In addition, dynamin appeared at midzone during anaphase and telophase (Fig. 1E and F). At cytokinesis, dynamin was highly concentrated at midbody (Fig. 1G). Thus, dynamin 2 was highly concentrated at apparent microtubule structures formed during mitosis.
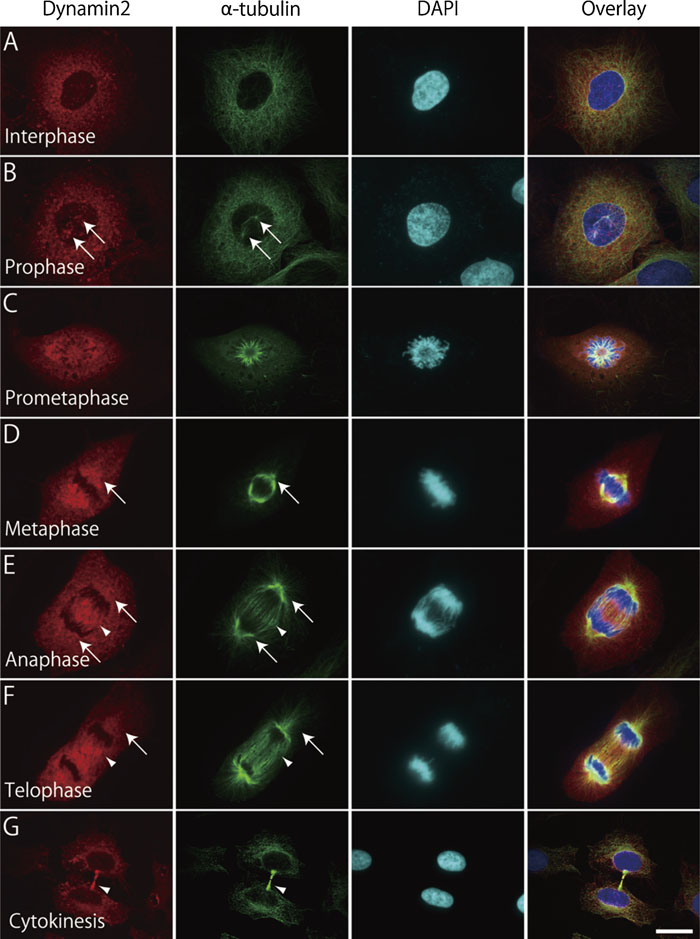 View Details | Fig. 1. Dynamin 2 localizes on microtubules during mitosis. Synchronized HeLa cells were fixed at 16 h after removal of hydroxyurea, and stained with anti-dynamin 2 (red), anti-α-tubulin (green) and DAPI (blue). Cells were observed using a confocal laser-scanning microscope. Arrows indicate centrosomes (B), polar microtubules (D–F). Arrowheads indicate kinetochore microtubules (E and F) and midbody (G). Bar: 20 μm (A–C and G), 12 μm (D–F). |
To elucidate the association between dynamin and microtubules, HeLa cells were permeabilized with 1% TX100 before fixation, then the cells were double stained by immunofluorescence (Fig. 2). Immunoreactivity for dynamin was hardly visible at interphase because of the cytosol extraction. However, dynamin began to appear on microtubules at prophase, (Fig. 2B), and was well colocalized with the mitotic spindles throughout the entire mitotic stages (Fig. 2C–E). Dynamin was also present at midzone at anaphase and telophase (Fig. 2E and F). At cytokinesis, dynamin was concentrated at regions near midbody but not along the entire bundle of intercellular bridge microtubules (Fig. 2G). Subcellular localization of dynamin at microtubular structures during mitosis was confirmed by immunoelectron microscopy. As shown in supplementary Fig. 1, spindle microtubules and astral microtubules were heavily decorated with immunogold particles for dynamin. These results clearly indicate that dynamin 2, at least isoform aa in HeLa cells, strongly associates with microtubules in mitosis.
 View Details | Fig. 2. Dynamin 2 localization on microtubules during mitosis becomes apparent after cytosol extraction. HeLa cells were synchronized by thymidine-hydroxyurea, fixed at 14 h after removal of hydroxyurea. Before fixation, cells were treated with 1% TX100 for extraction of cytosol fraction. Cells were stained by anti-dynamin 2 (red), anti-α-tubulin (green) and DAPI (blue). Bar: 20 μm (A), 10 μm (B–G). |
Considering that dynamin associates with microtubules during mitosis, either microtubule polymerization or depolymerization would alter the localization of dynamin. To test this hypothesis, microtubules of HeLa cells were stabilized with taxol, the microtubule polymerization drug, or nocodazole, the microtubule polymerization inhibitor, and the cells were double stained by immunofluorescence. As shown in Fig. 3, taxol treatment resulted in apparent colocalization of dynamin and microtubules not only in mitosis but also in interphase. On the other hand, nocodazole treatment resulted in the disappearance of dynamin (see supplementary Fig. 2). These results clearly indicate that dynamin is localized on the microtubules.
 View Details | Fig. 3. Dynamin 2 localization is enhanced by microtubule polymerization. Synchronized HeLa cells were treated with 2.5 uM Taxol for 16 h, fixed and visualized by anti-dynamin 2 (red), anti-α-tubulin (green). Cells at interphase (A), at metaphase (B) and anaphase (C) are shown. Bar: 20 μm (A), 10 μm (B and C). |
Since dynamin binds microtubules via PRD (Hamao et al., 2009), we investigated whether this is the case in mitosis. Toward this end, localization of exogenously expressed dynamin 2 wild type (WT) or deletion of PRD (ΔPRD) was examined during mitosis. Dynamin 2 WT was localized on both the mitotic spindle and midbody (supplementary Fig. 3), but dynamin ΔPRD did not (Fig. 4). This difference was clearly observed in cytosolic-extracted cells (Fig. 5). These results indicate that dynamin is localized on the mitotic spindle through its PRD domain.
 View Details | Fig. 4. PRD of dynamin 2 is required for the localization on microtubules. Synchronized HeLa cells, which were introduced by dynamin 2-ΔPRD, were fixed and stained by anti-dynamin 2 (red), anti-α-tubulin (green) and DAPI (Blue). Cells were observed using confocal scanning microscope. Bar: 20 μm (A and G), 10 μm (B–F). |
 View Details | Fig. 5. PRD is required for the localization on both mitotic spindle and midbody. HeLa cells were introduced by dynamin 2 WT (A and C) or dynamin 2-ΔPRD (B and D), and were subjected to cytosol extraction by the treatment of TX100 as decribed in Materials and Methods. Cells were fixed and stained by anti-V5 (red) and anti-α-tubulin (green) antibodies. Arrows indicate mitotic spindle (A) and midbody (C), respectively. Bar: 10 μm. |
Next, we investigated whether the dynamin PRD-microtubule interaction is required for cell cycle progression, in which microtubules play an indispensable role. As the middle domain of dynamin is known to interact also with γ-tubulin, a component of centrosome that plays a key role at the entry of mitosis (Thompson et al., 2004), a deletion mutant of the middle domain (ΔMiddle) was also included in following experiments. We introduced dynamin 2 WT, ΔPRD, ΔMiddle or K44A (as control) to HeLa cells, and measured the mitotic index. In this assay, the mitotic cell population was subdivided into two groups, those from prophase to telophase and those at cytokinesis, representing the entry into mitosis and the exit from mitosis, respectively. Although cells at prophase-telophase were not changed significantly by ΔPRD or K44A (Fig. 6A), both mutants doubled the percentage of cells at cytokinesis (Fig. 6B), indicating that both the GTPase domain and PRD might be required for the exit from mitosis. As the expression of ΔPRD did not affect dynamin-dependent endocytosis, in contrast to K44A (Fig. 6C), this suggested that the GTPase domain and PRD might function through two distinct mechanisms.
 View Details | Fig. 6. Both PRD and middle domain are required for cell cycle progression. To analyze mitotic index, HeLa cells expressing V5-tagged WT or mutant constructs were fixed and stained with anti-V5, anti-alpha-tubulin, and DAPI. Mitotic index was calculated as percentage of cells at prophase-to-telophase (A) or at cytokinesis (B) against total V5-positive cells (mean±SEM, three independent experiments, n>20 cells in each condition, *p<0.01, **p<0.001 compared with control). For uptake assay, HeLa cells transfected with dyamin2 wildtype, ΔPRD or K44A, were incubated with 5 μg/ml tetramethylrhodamine-labeled human transferrin for 30 min, fixed and stained with anti-V5 (C). Bar: 20 μm. |
The expression of the ΔMiddle significantly reduced the mitotic index of prophase-telophase ,but not that of cytokinesis (Fig. 6A and B), indicating that the middle domain is required for cell cycle progression, probably at the entry of mitosis. The expression of ΔMiddle consistently resulted in the loss of localization dynamin at the centrosome (supplementary Fig. 5) (Thompson et al., 2004).
Dynamin plays an essential role in endocytic vesicle formation. Other functions, including actin reorganization, centrosome cohesion and dynamic instability of microtubules, have been reported (Kaksonen et al., 2006; Schafer et al., 2002; Tanabe and Takei, 2009; Thompson et al., 2004). Here, we elucidated that dynamin 2 in mitotic cells is associated with the microtubules, and that this interaction depends on the PRD. Moreover, dynamin mutant lacking microtubule localization impaired the completion of cytokinesis, probably through a mechanism independent from intracellular traffic.
In this study, we elucidated the localization of dynamin during whole mitotic stages by immunofluorescence and by immunoelectron microscopy, and demonstrated that dynamin accumulated on the mitotic spindle throughout mitosis. This localization depends on the formation of microtubules, as perturbation of microtubule organization altered dynamin localization (Fig. 4 and supplementary Fig. 2). Dynamin polymerizes by itself into rings or spirals with inner diameter of 25 nm (Hinshaw and Schmid, 1995), or around microtubules, the inner diameter of which is also 25 nm, and provides a preferable scaffold for dynamin polymerization (Maeda et al., 1992).
We have recently reported that dynamin is associated with microtubules at interphase and regulates the dynamics (Tanabe and Takei, 2009). In mitosis, in which the dynamic rearrangement of microtubules actively takes place, dynamin may regulate the dynamics in similar fashion (Tanabe and Takei, 2009). Of course, there are other interpretations as to the localization of dynamin. One is that dynamin might assist mitotic spindle formation as was the case with clathrin. When clathrin is depleted by siRNA, mitotic spindle formation was impaired and mitotic index was increased (Royle et al., 2005). Another possibility is that dynamin is sequestered on the mitotic spindle in order to halt the recycling pathway (Boucrot and Kirchhausen, 2007). In any case, further investigation is required to clarify the physiological significance of the mitotic spindle localization of dynamin.
The PRD of dynamin is a module that interacts with SH3 domain-containing proteins (Cao et al., 1998; Qualmann et al., 2000; Witke et al., 1998) in a way that stimulates dynamin’s GTPase activity, which is required for membrane fission. PRD also mediates interaction with microtubules in vivo and in vitro (Hamao et al., 2009; Herskovits et al., 1993), to stimulate GTPase activity in vitro (Herskovits et al., 1993). Recent evidence shows that deletion of PRD results in the reduction of microtubule dynamics in vivo (Hamao et al., 2009), suggesting a role of dynamin in microtubule dynamics required for cell cycle progress.
Association of dynamin with microtubules was more apparent during mitosis than in interphase, raising the question as to how the interaction is regulated. It has been shown that Cdc2, a cyclin dependent kinase required for entry into mitosis, phosphorylates dynamin 2 in vitro (Hosoya et al., 1994). Cdc2-dependent phorphorylation of dynamin may enhance the association with microtubules during mitosis, although other mechanisms that involve phosphorylation or interaction with other binding partners cannot be excluded.
Our results have shown that PRD was not required for mitotic spindle formation (Fig. 6) and progression, but impaired cytokinesis. Although similar results were observed by the expression of K44A, a dominant-negative dynamin that blocks dynamin-dependent endocytosis, only K44A but not ΔPRD blocked dynamin-dependent endocytosis. This suggests that dynamin ΔPRD mutant blocked cytokinesis independent of membrane traffic, probably through the regulation of microtubules. In accordance with this, the deletion of PRD resulted in the disappearance of colocalization with microtubules. As dynamin regulates dynamic instability of microtubules through the independent mechanism of endocytosis (Tanabe and Takei, 2009), the expression of ΔPRD may impair microtubule dynamics at midbody. There is another possibility that dynamin regulates actin polymerization through PRD interacting proteins, such as cortactin (Schafer et al., 2002; Mooren et al., 2009), and that the PRD deletion results in impairment of actin contractile ring formation. Alternatively, K44A and ΔPRD could indirectly perturb mitotic progression due to sequestration of other binding partners or by promoting dominant effect on other processes that influence mitosis. Whether or not this is the case requires further study to reveal the molecular mechanisms involved.
On the other hand, the middle domain, which associates with γ-tubulin and is required for centrosomal cohesion, was also required for the entry into mitosis. Centrosomes serve as an important platform for various signals, including CDKs, Aurora, and PLKs (Doxsey et al., 2005). Deletion of the middle domain may result in the impairment of the function of centrosomes. We found that dynamin associates with the mitotic spindle and midbody via distinct domains, and that these interactions are required for cell cycle progression, probably both at the entry into mitosis and cytokinesis. Although the loss of localization around the mitotic spindle was not an apparent defect in the cell cycle progression, there may be a checkpoint for the correct formation of the mitotic spindle that resulted in blockage at cytokinesis. Further studies would clarify the physiological significance of localization of dynamin at the mitotic spindle.
We thank M.A. McNiven (Mayo Clinic) for providing Dynamin 2aa constructs. This work was supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
|