| Edited by Minoru Murata* Corresponding author. Present address: 6-14-10 Kasugadai, Nishi-ku, Kobe 651-2276, Japan |
Two true-bred lines that have the AABB genome constitution and ne1ne1 Ne2Ne2 genotype were obtained by transferring the Ne2 gene (one of two complementary genes for hybrid necrosis; Tsunewaki, 1960) from the common wheat ‘Jones Fife’ (abbreviated JF) to the emmer wheat ‘Vernal’. One line stably expressed a chlorophyll abnormality known as striato-virescence. During the genetic analysis of this abnormality, two other chlorophyll abnormalities, delayed virescence and albino, appeared. Lines fixed for the genes of those abnormalities were bred and subjected to aneuploid analysis using a set of disomic substitution lines of the durum wheat ‘Langdon’ (abbrev. Ldn DS lines), in which the Ldn’s A- or B-genome chromosome is substituted by its D-genome homoeologue of the common wheat cultivar ‘Chinese Spring’ (abbrev. CS) (Joppa and Williams, 1988).
Striato-virescence was reported by Frankel (1950), but the chromosome basis of its inheritance is unknown (McIntosh et al., 1998). Delayed virescence is a new chlorophyll abnormality. Albino is a chlorophyll mutation frequently seen by wheat workers, but so far no locus has been identified for it due to the lethality of albinotic plants. Aneuploid analyses of these three chlorophyll abnormalities therefore have been made.
The findings reported here provide new information on the genetic bases of chlorophyll abnormalities in wheat. Moreover, they show the limitation of aneuploid analysis that uses DS lines for traits determined by duplicated recessive genes.
The following names are used here to indicate different wheat genotypes: ‘Emmer wheat’ for tetraploid genotypes having the AABB genome constitution (2n = 4x = 28) and ‘common wheat’ for hexaploid genotypes of the AABBDD genome constitution (2n = 6x = 42). Cultivars of T. dicoccum and T. durum, both belonging to Emmer wheat, respectively are called emmer and durum wheat.
Designations of the wheat chromosomes used in this study follow the recommendations of the 7th International Wheat Genetics Symposium (Miller and Koebner, 1988). The former 4A is renamed 4B, and the former 4B renamed 4A.
The abbreviation ‘sv’ is used for this phenotype, and the basic symbol, sv, adopted for its genes. This abnormality is characterized by normal green seedling leaves in autumn, albinotic stripes on rosette leaves in winter (Fig. 1A, right), and recovery of normal leaf color in spring. When sv homozygotes are grown in a greenhouse adjusted at 15°C or higher temperature, they do not express this abnormality.
 View Details | Fig. 1. (A): Rosette stage of a normal (left), delayed virescent (center), and striato- virescent plant (right). (B): Normal (four seedlings at left) and albinotic plants (four seedlings at right) segregated in an F4 progeny of the cross Ldn x (Vernal x sv Ne2-4x)F1. |
Two tetraploid lines (2n = 28) homozygous for Ne2 genes were bred by transferring this gene from common wheat JF to the Emmer (Vernal) background. One line, named sv Ne2-4x, stably expressed the sv phenotype. Because Ldn carries Ne1 which causes hybrid necrosis in the presence of Ne2 (Tsunewaki, 1960), breeding an Ne2-free sv homozygote was necessary to determine the chromosomal locations of sv genes by means of aneuploid analysis with use of Ldn DS lines. This was done by selecting sv homozygotes in the offspring of the cross, Ldn x (Vernal x sv Ne2-4x) F1, in which Vernal is free of both Ne1 and Ne2. The sv-homozygous line obtained that is free of Ne2 was named sv ne2-4x.
The abbreviation for this phenotype is ‘dv’. The basic symbol dv is used for its genes. This phenotype is characterized by normal green-colored seedlings in autumn, which turn almost completely albino in winter (Fig. 1A, center). Most dv seedlings die at that stage, but a few survive through winter. Those that do gradually recover chlorophyll and growth vigor in the spring and finally set seeds. Their maturity, however, is about one month later than that of normal plants. When dv homozygotes are grown in a greenhouse adjusted at 15°C or higher temperature, they do not show delayed virescence, although their growth vigor is subnormal. A few fixed dv lines free of Ne2, obtained in the offspring of the cross Ldn x (Vernal x sv Ne2-4x) F1, were used in the present analysis.
The abbreviation for this phenotype is ‘abn’. The basic symbol abn is used for its genes. Albino seedlings lack chlorophyll from the time of germination (Fig. 1B), are pure white (or slightly purple when anthocyanin is present), and die at about the third leaf stage when the starch in the attached seeds is exhausted. Because albinos are completely lethal, no fixed albino lines were obtained. True-bred lines for each of the duplicated (or complementary) recessive abn genes therefore were selected from the offspring of the cross Ldn x (Vernal x sv Ne2-4x) F1, and used for abn gene identification. Details of the procedure for their breeding are given in the corresponding section in “RESULTS”.
A complete set of the 14 Ldn DS lines produced by Joppa and Williams (1988) was used. Each DS line is indicated by its D-genome chromosome, the replaced A- or B-genome homoeologue being given in parentheses. For example, Ldn 1D(1A) is a DS line in which a pair of Ldn 1A chromosomes is replaced by a pair of CS 1D chromosomes. Of the 14 DS lines, 4D(4A), 5D(5A), and 6D(6B) respectively retained a single 4A chromosome, single 5AL arm, and single 6BS arm.
In a preliminary study, the 14 DS lines and a normal Ldn line, the control, were crossed as female to (Vernal x sv Ne2-4x) F1’s. For convenience, seeds obtained from these crosses are hereafter called F1 seeds. Open-pollinated F2 seeds were harvested from 30 F1 plants of each DS line and 40 F1’s of the control, but the 3D(3B) line gave only 11 F1 plants. The F2 seeds were sown in flats in a greenhouse, and the germination rate and frequency of albinos examined. All the green seedlings were transplanted to the field in late autumn. Segregation of dv and sv plants was examined a few times during the winter.
In the advanced studies, the same set of DS lines and a normal Ldn line were crossed to sv ne2-4x, to the dv homozygote, and to the homozygote for each of the two complementary abn genes. F1 plants were grown in pots in a greenhouse, and F2 seeds obtained by self-pollination were sown in flats also in a greenhouse where segregation of albinos was observed. In early winter, the flats were taken outside. Segregation of dv and sv plants in the individual F2 progenies was recorded a few times during the winter.
As expected, the F1 populations of all the DS and control lines, except the Ldn 5D(5B) line, segregated normal and necrotic plants in about a 1 : 1 ratio. Line Ldn 5D(5B) lacks the 5B chromosomes of Ldn which carry Ne1. All the offspring therefore should be non-necrotic. In fact, 137 plants were normal, two others necrotic. The origins of these off-type plants are unknown. All the normal hybrids are assumed to be homozygous for ne2. All the parental lines in the advanced studies were descendants of those normal F1’s.
All three chlorophyll abnormalities segregated (Table 1) in F2 offspring of the normal F1’s. Several DS lines clearly showed segregation ratios for each chlorophyll abnormality that differed from those of the disomic population. It was, however, difficult to definitively determine which chromosomes carry the responsible genes. The reasons were (1) frequencies of all the abnormalities were low in the disomic (control) population, (2) co-segregation of all three abnormalities occurred, and interaction was suspected, (3) the Ne2 gene appeared to be linked to some chlorophyll abnormality genes. For a clear identification of the carrier chromosomes of the responsible genes, advanced studies therefore were made on fixed lines for each abnormality, which lines were free of the other abnormality genes.
 View Details | Table 1. Segregation of three chlorophyll abnormalities in the F2 generation of the crosses Ldn DS x (Vernal x sv Ne2-4x)F1. |
A fixed line for this abnormality, free of Ne2 and other chlorophyll abnormality genes, was crossed to Ldn DS lines and to a normal line (the control). F2 populations obtained by the self-pollination of F1 hybrids were examined for segregation of sv plants (Table 2). The frequency in most lines was close to 6.25% (= 1/16), indicative that two duplicated recessive genes determine the expression of this phenotype. The postulated genes were assigned the symbols sv1 and sv2.
 View Details | Table 2. Frequency of striato-virescence (sv) in the F2 generation of the crosses Ldn DS’s x sv ne2-4x. |
Disomic and pooled data indicated that the frequency of sv plants is slightly, but significantly lower than the expected 6.25%. This deviation is not attributable to female sterility because female fertility of the disomic F1’s was normal (average 94%). One possible explanation is that there is a slight disadvantage (estimated to be about 5%) in fertilization of pollen of the sv1sv2 genotype as compared to pollen having the Sv1 and/or Sv2 allele.
Based on the 15 green to 1 sv ratio, a χ2-test was made (second column at right, Table 2). F2 progenies of two DS lines, Ldn 3D(3A) and Ldn 2D(2B), had ratios that deviated significantly from the expected value. In the Ldn 3D(3A) progenies, the frequency of sv plants was about two times the expected 6.25%. The conclusion is that one of the duplicated genes, sv1, is located on chromosome 3A of sv ne2-4x. According to Joppa and Williams (1988, Table 5), about 41% of the F2 offspring of 3A/3D double monosomics have a pair of 3A chromosomes. Therefore, about 10% (i. e., 41% x 1/4) of the F2 offspring of the cross, Ldn 3D(3A) x sv ne2-4x, are expected to have the sv phenotype, assuming that a double dose of sv1, in addition to the homozygous condition of sv2, is necessary for the sv phenotype. The actual frequency of 10.2% for offspring of 3D(3A) lines fits that frequency.
The extremely low frequency of sv plants in the F2 progenies of Ldn 2D(2B) suggests that chromosome 2B or 2D carries an sv suppressor. The parental F1’s, Ldn 2D(2B) x sv ne2-4x, differ from the control F1’s in the following two respects. One is the presence of the 2B chromosome in a single dose, as compared to the double dose in the control. The other is the presence of the 2D chromosome that is lacking in the control. Because the 2B chromosome in the Ldn 2D(2B) x sv ne2-4x F1’s was derived from the sv ne2-4x parent, it can not be the carrier of the sv suppressor. Therefore, chromosome 2D must carry the suppressor. According to the findings of Joppa and Williams (cited above), the frequency of 2D-chromosome carriers in the F2 offspring of 2B/2D double monosomics is about 34%, close to 1/3. The frequency of sv plants in the F2’s of the cross Ldn 2D(2B) x sv ne2-4x therefore is expected to be about 1/4 x 1/4 x 1/3 (ca. 2.1%). The observed frequency of 1.0% fits the expected ratio (χ2 = 0.670, df = 1).
Based on the above deduction, F2 progenies of the cross Ldn 2D(2A) x sv ne2-4x should segregate sv plants at a much lower frequency than do disomic F2’s and should show an sv plant frequency similar to that of the cross Ldn 2D(2B) x sv ne2-4x because they have the 2D chromosome in common. This, however, was not the case. They segregated sv plants at a frequency similar to that of the disomic F2’s, indicative that chromosome 2A of sv ne2-4x carries the sv2 gene; the other striato-virescence gene. According to Joppa and Williams (cited above), the frequency of F2 plants that have a double dose of the 2A chromosome (accordingly, a double dose of the sv2 gene) but which lack the 2D chromosome (therefore being free of the sv suppressor) is about 35%. The frequency of sv plants in F2 progenies of the cross Ldn 2D(2A) x sv ne2-4x therefore is expected to be about 0.35 x 1/4, about 8.7%. This fits the observed frequency of 5.0% (χ2 = 3.522, df = 1).
The sv suppressor is assumed to be a wild-type homoeoallele of the sv2 gene as they are located on the 2D and 2A chromosomes respectively in the same homoeologous group. This suppressor therefore is designated Sv3. The fact that the frequency of sv plants in the preliminary study (2.1%, ref. Table 1) was about half that in the present study (5.0%) is explained by the genotypic difference of the male parents crossed to Ldn DS lines; Sv1sv1 sv2sv2 in the preliminary study (ref. Table 1) and sv1sv1 sv2sv2 in this one.
Fixed lines with delayed virescence, free of Ne2 and other chlorophyll abnormality genes, were crossed to Ldn DS lines, and the F1 hybrids self-pollinated. F2 offspring were tested for the segregation of dv plants (Table 3).
 View Details | Table 3. Segregation of delayed virescence (dv) in the F2 generation of the crosses Ldn DS’s x dv homozygote. |
The frequency of dv plants was close to 25% in about 1/3 of the DS lines, but all the lines had frequencies lower than 25%. Control F1’s were fully fertile on the female side (97% selfed seed fertility), but the F2 progenies had frequencies of dv plants lower than 25%. This indicates that there is a fertilization disadvantage in having dv-carrying pollen as opposed to having Dv-carrying pollen (the estimated disadvantage is ca. 14%). According to the F2 segregation ratio, the Ldn and dv homozygotes appear to differ by a single gene pair.
Segregation ratios of dv plants in F2 progenies of the DS lines were tested against the control F2 (disomic) ratio because the frequency of dv plants in the controls statistically was lower than 25%. F2 progenies of two DS lines, Ldn 2D(2A) and Ldn 7D(7A), had ratios that deviated significantly from the disomic ratio; lower in the former, higher in the latter. Ldn 2D(2B) progenies did not differ from the disomic ones. These findings suggest that the 2D chromosome carries a suppressor for dv expression and that the 2B chromosome has a dv gene. If not, the F2 progenies of Ldn 2D(2B) would segregate dv plants at a low frequency similar to that of Ldn 2D(2A). The dv frequency in F2 progenies of Ldn 7D(7A) was 22.1%, only a little lower than 25%, indicative that chromosome 7A has no relation to the dv phenotype.
Two conclusions are drawn: (1) delayed virescence is controlled by a single gene, dv1, located on chromosome 2B of the dv homozygote (consequently, the 2B of Ldn has its wild-type allele, Dv1); (2) chromosome 2D of CS carries a dv locus on which a wild-type homoeoallele of dv1is located. This wild-type homoeoallele is designated Dv3 because of its presence in the third wheat genome, D. Based on these assumptions, the respective frequencies of dv plants in the F2 progenies of the two critical DS lines Ldn 2D(2A) and Ldn 2D(2B) should be 6.2% and 25.7%, close to their observed frequencies of 8.5% and 21.0%.
As stated hereafter (ref. Table 6), the F2 population of the cross Vernal x Ldn produced some dv plants, indicative that each parent has one or the other duplicated recessive dv gene. Consistent with the previous conclusions, the genotype of Ldn must be Dv1Dv1 dv2dv2 and that of Vernal dv1dv1 Dv2Dv2. Although the carrier chromosome of the dv2 locus was not determined in this study, that locus is assumed to be on the 2A chromosome based on the homoeology of the 2A chromosome to both the 2B and 2D chromosomes, carriers of the dv1 and dv3 loci, respectively. The dv homozygote should have the dv2 allele in common with Ldn because they showed a single gene difference, dv1vs Dv1.
The preliminary study showed that F2 progenies of the cross Ldn x (Vernal x sv Ne2-4x) F1 segregated some albinos together with other chlorophyll abnormalities (Table 1). About ten green F2 plants (being the ne2ne2 homozygotes, free of Ne2) were selected from these progenies, and self-pollinated. Their F3 generations were tested for segregation of albinos. F3 progenies derived from four F2 plants segregated albinos. A few green sibs were self-pollinated in each of the four F2 families. F4 progenies also were tested for ablino segregation. Three to five F4 progenies that segregated green and albino plants at nearly a 3:1 ratio were selected from each F2 family. Five green sibs were selected in each F4 progeny, and subsequent progenies (F5 generation) again tested for albino segregation. Totally, 74 F5 progenies were tested, of which 32 were fixed for the green phenotype and assumed to be homozygous for one of the duplicated albino genes. Of those progenies, 20 were used in the following allelism test of abn genes.
Each of the 20 test lines and a normal Ldn line were crossed to at least two other test lines. The F1 hybrids (in Table 4, hybrids between F6 lines) were self-pollinated, and their F2 progenies tested for segregation of albino plants. Table 4 shows that neither the F2’s of crosses between lines of the No. 1 to 3 F2 families (Group I cross combinations in the Table) nor those between lines of the No. 4 family (Group II combinations) segregated any albino plants, whereas F2’s between lines of the No. 1 to 3 families and those of the No. 4 family (Group III combinations) all segregated some albino plants. These findings indicate that lines of the No. 1 to 3 F2 families carry one of the duplicated albino genes (abn1), whereas those of the No. 4 family carry the other one (abn2). Ldn should have abn1.
 View Details | Table 4. Segregation of albinos in self-pollinated progenies of hybrids between F6 lines derived from the cross Ldn x (Vernal x sv Ne2-4x) F1. |
Because Ldn is a carrier of the abn1 gene, the abn2 gene alone was subjected to aneuploid analysis, for which a few lines of the abn2 homozygote (Group II lines in Table 4) were crossed to Ldn DS lines and to a normal Ldn line as the control. The F1 hybrids were self-pollinated, and the F2 progenies tested for abn plant segregation.
Table 5 shows that the overall frequency of abn plants was lower than the 6.25% expected based on two recessive gene segregations. Again, the disadvantage for fertilization of having abn-carrying pollen as compared to Abn-carrying pollen is indicated. Because the female fertility of the disomic F1’s was normal (ca. 98% after self polli-nation), the disadvantage for fertilization of having abn1abn2-pollen is estimated to be ca. 15%.
 View Details | Table 5. Albino frequencies in the F2 progenies of the crosses Ldn DS’s x abn2 homozygote. |
To identify which chromosomes carry the abn loci, abn frequencies in F2’s of the DS lines were compared to the disomic ratio. Only a single line, DS 2D(2A), which segregated no albino, deviated significantly from the disomic ratio. This is explained by the hypothesis that chromosome 2A of Ldn carries the abn1 gene and that the 2D chromosome of CS carries its suppressor. Owing to the homoeology of chromosomes 2A and 2D, this suppressor was named Abn3 for its presence in the third wheat genome.
The presence of the Abn3 gene on chromosome 2D should reduce the frequency of abn plants in the F2 population of the cross DS 2D(2B) x abn2 homozygote as compared to the disomic frequency, but it did not (ref. Table 5), indicative that chromosome 2B of the abn2 homozygote carries abn2, the other albino gene. Therefore, 2.2% of the above F2’s would be expected to be albino, assuming that the respective frequencies of the 2D-free female and male F1 gametes are 19.7% and 62.5% (after Joppa and Williams, 1988) and that there is a 15% disadvantage for participation in fertilization in having abn1abn2-pollen as compared to having wild type pollen. The observed frequency of 3.3% fits this expected frequency (χ2 = 2.81, df = 1). Two conclusions therefore were drawn: (1) abn1 and abn2 respectively are located on chromosomes 2A and 2B, and (2) chromosome 2D of CS carries Abn3, the wild-type homoeoallele.
The sv trait first was found in six-times self-pollinated offspring of the SB2 hybrids, JF x Vernal3, in which Vernal was the recurrent pollen parent. Of the SB2 hybrids, those having the normal chromosome complements of Emmer wheat were selected cytologically and used for further self-pollination. Because sv Ne2-4x has both the sv1 and sv2 genes and Vernal shows a single gene difference, the undoubted origin of either sv1 or sv2 is Vernal, and therefore the other gene must be derived from JF. It is not clear, however, which gene is from Vernal and which from JF.
As for the origin of dv genes, the F2 generation of the cross, Vernal x Ldn, segregated some dv plants (ref. Table 6), indicative that the dv trait is controlled by two recessive genes and that Vernal carries one and Ldn the other. The dv homozygote is the double recessive for those genes. One of them, dv1, is located on chromosome 2B of the dv homozygote, and Ldn carries the dominant allele Dv1 on the same chromosome. Clearly, the dv1 and dv2 of the dv homozygote originated respectively from Vernal and Ldn.
 View Details | Table 6. Segregation of three chlorophyll abnormalities in the F2 generations of the parental stocks. |
The F2 generation of the same cross, Vernal x Ldn, also segregated some albino plants. Because Ldn carries the abn1 gene (ref. Table 4), Vernal must carry the other albino gene, abn2. The respective origins of abn1 and abn2 therefore are Ldn and Vernal.
Loci and alleles identified for the three chlorophyll abnormalities of striato-virescence, delayed virescence, and albino are summarized in Table 7.
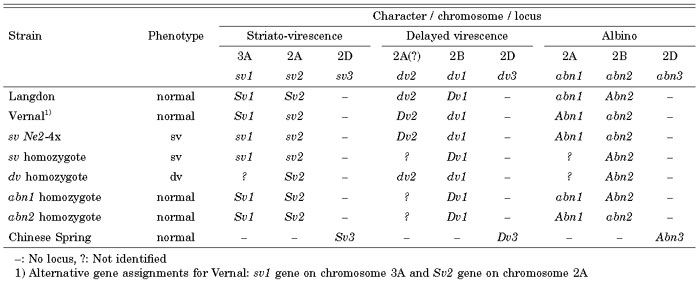 View Details | Table 7. Genotypes determined by aneuploid analyses of parental and derivative strains of the three chlorophyll abnormalities. |
Frankel (1950) reported triplicated recessive genes for striato-virescence in common wheat. The present studies indicate the presence of two loci, sv1 and sv2, in Emmer wheat, and a third sv3 locus in the D genome of common wheat in agreement with his findings. Nishikawa (1986) reported strong association of the albino phenotype with a 2B telosome in various hybrid populations derived from F1’s between Vernal and 2B double ditelocentrics of the durum wheat ‘LD222’. This suggests that the abn1 gene is present on chromosome 2B of LD222 as in Ldn and of abn2 in Vernal, which supports the present findings.
Except for sv1, all the loci are located on group 2 homoeologous chromosomes. Chromosome 2A bears the sv2, dv2 (possibly), and abn1 loci. Of the three mutants, the sv2 allele is linked to neither dv2 nor abn1. The last two genes may be linked, but recombination takes place between them because both delayed virescent and albinotic plants were segregated among normal plants in self-pollinated offspring of a single hybrid plant (ref. Table 1). All three mutant genes therefore must have separate loci. Similarly, chromosome 2B carries the dv1 and abn2 loci. Again, recombination takes place, providing evidence that their loci differ (Table 1 and Table 7).
Two other types of chlorophyll abnormalities, virescence and chlorina, have been reported in common wheat (ref. McIntosh et al., 1998). Virescence is controlled by two loci; v1 on chromosome 3B (Neatby’s virescence) and v2 on chromosome 3A (Hermsen’s virescence). Chlorina is controlled by three homoeoloci; cn-A1, cn-B1 and cn-D1 on group 7 chromosomes. Here, except for the sv1 locus on chromosome 3A, all the sv, dv and abn loci located on group 2 chromosomes are independent of the v and cn loci. Virescence, chlorina, striato-virescence, delayed virescence, and albino therefore all have different genetic bases, but whether there is a relationship between v2 and sv1, both on chromosome 3A, is not known.
Complementary genes, such as the Ne1 and Ne2 for hybrid necrosis, Ch1 and Ch2 for Type 1 hybrid chlorosis, and Cs1 and Cs2 for Type 2 hybrid chlorosis (Hermsen, 1963; Tsunewaki and Nakai, 1973), have proved useful in studies of phylogenetic differentiation in polyploid wheats. This type of system is extremely efficient because the genotype of the individual being tested can be determined immediately from phenotypes of the F1 hybrids between the testers and that individual. Unfortunately, in polyploid wheats the known complementary gene systems are few.
In Emmer wheat, the duplicated recessive genes for chlorophyll abnormalities reported here can be used. For example, Ldn and Vernal can be used as respective testers for the delayed virescence genes dv1 and dv2. If a cultivar carries dv1 or dv2, its F2 progeny from a cross to Ldn or Vernal will segregate some delayed virescent plants. For albino, the same tester combinations, Vernal (or the abn2 homozygote bred here) and Ldn (or the abn1 homozygote) respectively can be used to determine the presence of abn1 and abn2 genes. For striato-virescence, we need to breed a tester set, sv1 and sv2 homozygotes, from the present sv homozygote.
The efficiency of using this type of gene system, i. e., duplicated recessive genes, in phylogenetic studies is much lower than using the complementary gene systems because genotype determination only is possible by observing F2 segregation which requires additional generation time and a large number of offspring for segregation detection. A presumed advantage is the probable existence of a greater number of duplicated recessive gene systems in polyploid wheats as compared to complementary gene systems. This is suggested by the fact that all three chlorophyll abnormalities studied are controlled by this type of gene system.
In common wheat, monosomic analysis that uses a complete series of 21 monosomics (Sears, 1954), has proved a simple, very effective means of determining the chromosomal locations of genes for a variety of characters. In Emmer wheat, Mochizuki (1968) produced a similar monosomic series. Its use in the gene analysis of Emmer wheat, however, is limited because recovery of monosomics in the offspring averaged only 11% for all the monosomics compared to the ca. 75% for common wheat offspring. No monosomic analysis therefore has been conducted with the Mochizuki’s series. The set of DS lines of Ldn wheat produced by Joppa and Williams (1988) provides an alternative means of aneuploid analysis. Konzak and Joppa (1988) first used the set in their gene analysis of chocolate chaff in durum wheat and successfully located the responsible recessive gene, cc, on chromosome 7B of that wheat and its suppressor (wild-type homoeoallele) on 7D of CS. My previous work on Type 2 hybrid chlorosis, in which Ldn DS lines were used, also was successful in locating Cs1 on chromosome 5A of Emmer wheat and Cs2 on chromosome 4G of Timopheevi wheat (Tsunewaki, 1992).
In contrast, the present findings on the chromosomal bases of three chlorophyll abnormalities are somewhat ambiguous, because all are controlled by duplicated recessive genes plus their wild-type homoeoalleles present on a D-genome chromosome of CS. As for the striato-virescence (sv) controlled by the duplicated loci sv1 and sv2, there is certainty that the location of the sv1 gene is on chromosome 3A, whereas the location of sv2 on chromosome 2A was deduced from a higher frequency of sv plants in offspring of the 2D(2A) line than in those of the 2D(2B) line, despite the fact that both carry the same suppressor, Sv3, on their 2D chromosomes.
Delayed virescence (dv) also was shown to be controlled by the duplicated recessive genes dv1 and dv2. The dv homozygote and Ldn differ in a single gene pair on their 2B chromosomes; the former has the dv1 allele, the latter the Dv1 allele. Direct evidence also indicated that chromosome 2D of CS carries its wild-type homoeoallele, Dv3. F2 data for the cross Ldn x Vernal show that each parent has one duplicated recessive gene, dv1 or dv2. Because Ldn was shown to carry Dv1 gene, this wheat was deduced as having the dv2 gene and Vernal dv1. The chromosome location of the dv2 locus is not clear from these findings. Based on the homoeology of chromosomes 2B and 2D, which respectively carry the dv1 and dv3 loci, I assumed that the third locus, dv2, may be on the remaining group 2 chromosome, 2A.
As for albino (abn), both the allelism test results for the abn homozygotes and the F2 data for the cross Ldn x Vernal indicated that this character also is controlled by two recessive genes, abn1 and abn2, and that Ldn carries the former and Vernal the latter gene. Aneuploid analysis of a fixed line for the abn2 gene showed that the 2A chromosome of that line carries the abn1 gene and that chromosome 2D of CS carries its wild-type homoeoallele, Abn3. Evidence for the abn2 gene being on chromosome 2B is indirect. F2 progenies of the DS 2D(2B) line segregated abn plants at a high frequency than those of the DS 2D(2A) line, being similar to that of the control F2’s, although this line carries the suppressor Abn3 on its chromosome 2D.
All the findings indicate that aneuploid analysis of Emmer wheat done with a set of Ldn DS lines is limited for making a clear-cut determination of the carrier chromosomes of duplicated recessive genes, in particular when a D-genome chromosome carries the wild-type homoeoallele. Taking into account this limitation, at present the set of Ldn DS lines is the only useful means of determining the chromosomal position of genes in this wheat group.
The task of breeding abn homozygotes was done with the help of Kenji Ichikawa, Imari Oda and Kayoko Morinaga, to whom I express my deepest gratitude. I also am grateful to L. R. Joppa, North Dakota State University, for his kindness in supplying the disomic substitution lines of Langdon durum, thereby facilitating the investigation reported here.
|