| Edited by Kiyotaka Okada. Masayoshi Shigyo: Corresponding author. E-mail: shigyo@yamaguchi-u.ac.jp |
Japanese bunching onion (Allium fistulosum L., 2n = 2x = 16, genome FF) is a most economically important crop in East Asia, i.e., Japan, China, Taiwan and South Korea (Inden and Asahira, 1990). Genetic studies in A. fistulosum are challenging because of cross-pollination, biennial generation, and severe inbreeding depression. A limited number of genetic studies have been published on A. fistulosum, including the mode of inheritance of cytoplasmic male sterility (Moue and Uehara, 1985) and polymorphic isozyme loci (Haishima and Ikehashi, 1992; Haishima et al., 1993; Mangum and Peffley, 1994).
Allium fistulosum contains several characteristics attractive to the breeding of A. cepa (bulb onion and shallot), such as resistance not only to pink root disease (Netzer et al., 1985), caused by the fungus Pyrenochaeta terrestris, but also to onion leaf blight (Currah and Maude, 1984), caused by Botrytis squamosa. Among all the interspecific crosses in Allium, hybridization between A. fistulosum and A. cepa has been carried out the most extensively because A. fistulosum shows desirable agronomic traits. Interspecific F1 hybrids between these two species have been readily produced and shown to be resistant to several kinds of A. cepa diseases (Brewster, 1994).
The F1 hybrids are completely sterile because of chromosomal rearrangements between the two species. The development of alien addition lines, therefore, has proved to be a practical approach for introgressing genes from the related species. Sears (1956) transferred genes for leaf rust resistance from Aegilops into wheat. Recently in Allium, a similar trial via A. fistulosum – A. cepa monosomic addition lines was applied in transferring rust resistance from chromosome 1A of A. cepa into A. fistulosum (Wako et al., unpublished data).
Allium cepa – A. fistulosum chromosome addition lines seem to be beneficial for the study of genome organization in A. fistulosum as well as for the practical breeding of A. cepa. Peffley et al. (1985) reported the production of four A. cepa – A. fistulosum monosomic addition lines (2n = 17) and several hypo-triploids (2n = 20, 22, 25). However, these lines still do not cover all the genomes of A. fistulosum. Further, Shigyo et al. (1994) proposed using hypo--allotriploids (2n = 3x – 1 = 23, AAF - nF) together with the A. cepa – A. fistulosum monosomic additions (2n = 2x + 1 = 17, AA + nF) in allocating the genes and genetic markers to the A. fistulosum chromosome.
The aims of the present study were to find the monosomic additions and hypo-allotriploids in the backcross progeny of amphidiploids between Allium cepa and A. fistulosum and to characterize them by means of several methods of analysis.
Our previous study using fertility restorers (tetraploids) of Wakegi onion (Allium x wakegi Araki), a natural hybrid between A. fistulosum and shallot (A. cepa Aggregatum group), revealed that a considerable number of hypo-allotriploids resulted from the crossing of the tetraploid A. x wakegi and A. cepa Aggregatum group (Tashiro et al., unpublished data). Therefore, the amphidiploids AAFF (2n = 4x = 32) as well as the allotriploids AAF were backcrossed to shallot (A. cepa L. Aggregatum group) as the pollinator in this study. The pedigree of backcross progenies is given in Fig. 1. The parts surrounded by the frames in this figure were carried out in this study. As seed par-ents, one F2 (SUA392) and three F1BC1 (SUA393, SUA394, SUA395) lines were provided by Saga University. During the 2000–2002 flowering season (March to May) in Yamaguchi, Japan, each plant used for seed and pollen parents was planted in a nursery pot, enclosed in a glass house, and covered with mosquito nets to prevent unexpected crossing. All crossings in this study were done by hand pollination. The capability of seed set was evaluated by the percentage of ovules that developed into seeds (PODS) and the germination rate, as described in our previous study (Shigyo et al., 2003).
 View Details | Fig. 1. Pedigree of plant materials obtained in this study. The crossings surrounded by the frame in this figure were conducted in this study. |
In all the backcross progenies in every generation, somatic chromosomes in squashed root tip cells were observed by Feulgen reaction staining.
All 224 plants of the F2BC1 (194 plants), F1BC2 (eight plants) and F2BC2 (22 plants) progenies obtained in 2002 were morphologically studied six months after sowing. Three characteristics of plantlets--number of tillers, number of expanded leaves per tiller and length of leaf blade --were evaluated to determine the morphological effects of different numbers of extra chromosomes in an Allium cepa diploid background.
In order to develop a rapid method for the identification of hyper- and hypo-allotriploids, flow cytometric analysis of three allotriploids with different numbers of chromosomes (2n = 23, 24, 25) was performed with a PA flow cytometer (Partec, Münster, Germany). The assay plants consisted of six hypo-allotriploid plants, six allot-riploids, and two hyper-allotriploids. These were selected from each chromosome group at random. Bet-ween eight and 10 months after sowing, a young leaf blade was collected from the assay plant, cut with a razorblade, and placed in DAPI staining solution (Partec) with a leaf blade of Allium vavilovii M. Pop. Et Vved., a diploid wild relative for A. cepa, which was used as the internal standard. Nuclei were counted for every fluorescence intensity (FI) at the G1 peak, and the extra chromosome index (EI; the mean FI of the test plant in relation to that for the internal standard, expressed as percentage) described in our previous report (Shigyo et al., 2003) was calculated for three different allotriploids. The measurement was repeated three times for each assay plant.
In our previous study (Shigyo et al., 1994), a monosomic addition of AA + 6F and hypo-allotriploid AAF - 6F (2n = 3x – 1 = 23) was employed to assign an isozyme gene locus Got-2 to chromosome 6F of Allium fistulosum. In this study, the chromosome 6F-specific isozyme marker, Got-2, was used to identify seedlings carrying this chromosome in every backcross progeny. All procedures for the isozyme analysis of glutamate-oxaloacetate transaminase (GOT; EC 2.6.1.1) through native-PAGE with the crude extract of enzyme from a young expanding leaf were conducted according to the procedure of Shigyo et al. (1994).
The amphidiploids and allotriploids were crossed with shallot as the male to produce BC1 and BC2 seeds, respectively. The amphidiploids had higher seed fertility [PODS = (1114 / 16140) × 100 = 6.9] than the allotriploids [PODS = (254 / 50520) × 100 = 0.5]. The germination rates of the BC1 seeds reached 38.4% [= (377 / 981) × 100]. The rates of the BC2 seeds were nearly the same [(86 / 220) × 100 = 39.1%]. Finally, 237 BC1 and 46 BC2 seedlings survived. The F2BC1 progenies were mainly allotriploids and hypo-allotriploids with 23 chromosomes. The frequency of two different allotriploids accounted for approximately 89% of all the F2BC1 seedlings (Table 1). The chromosome numbers of the BC2 seedlings varied from 16 to 24 for two different generations (F1BC2, F2BC2). Hypo-allotriploids (23 chromosomes) were the most frequent (43.5%), followed by monosomic additions (17.4%). Thus, several kinds of Allium cepa – A. fistulosum chromosome additions were generated from the first or second backcross.
 View Details | Table 1. Variation of chromosome numbers in BC1 and BC2 generations |
Several morphological characteristics of plantlets were evaluated in all the generations obtained in 2002 (Table 2). In the F2BC1 population, great differences among seedlings possessing different numbers of chromosomes were observed only in the length of the expanding leaf. Regarding the two major types of derivatives, the leaf lengths of the hypo-allotriploids (mean ± SE = 140.2 ± 4.9 mm) were clearly shorter than those of the allotriploids (183.3 ± 2.3). Except for a few cases in this generation, there was a tendency for the euploids to be larger and more vigorous than the aneuploids. In the BC2 population composed of the two generations, F1BC2 and F2BC2, clear differences appeared only in the leaf length. However, the euploids were smaller and less vigorous than the aneuploids in this population, contrary to our expectations.
 View Details | Table 2. Morphological characteristics of plantlets obtained in 2002 |
Fig. 2 shows typical histograms of flow cytometric analyses for three materials with different numbers of chromosomes in comparison with the internal standards. Bimodal peaks were formed when test plants were measured together with the internal standard for G1- and G2-phase nuclei. All three cases with different numbers of chromosomes had fluorescence intensities (FI) higher than the internal standard. The results from the G1 peak are summarized in Table 3. The FI value of test plants ranged from 111.66 to 115.3, whereas those for the internal standard ranged from 83.3 to 86.5. The P value (t-test) obtained when comparing the means of the FI in the test plants with those in the internal standard showed, in all three cases, significant differences (P < 0.001). There were sig-nificant differences in the extra chromosome index (EI) among three different kinds of allotriploids, and the mean values of EI rose as the number of chromosomes increased. Consequently, it was possible to distinguish three kinds of plant material with different numbers of chromosomes by flow cytometric analysis.
 View Details | Fig. 2. Representative histograms of nuclei isolated from mixtures of leaves of three plants with different numbers of chromosomes. (A) Allium vavilovii – hyper-allotriploid (2n = 3x + 1 = 25); (B) A. vavilovii – allotriploid (2n = 3x = 24); (C) A. vavilovii – hypo-allotriploid (2n = 3x – 1 = 23). |
 View Details | Table 3. Flow cytometric analysis for G1 peak of fourteen plant materials with different numbers of chromosomes in comparison with an internal standard, diploid Allium vavilovii |
Allium fistulosum possesses a pair of distinctive sub-telocentric chromosomes 6F (Jones, 1990). In this study, banding patterns for Got-2 were observed to determine whether the chromosome 6F existed in hyper-diploids (2n = 17–18) and hypo-allotriploids (2n = 21–23) (Fig. 3). According to Shigyo et al. (1994), Got-2 is a dimeric enzyme in A. fistulosum and A. cepa, and each of the two species has different alleles (f and a, respectively). In the backcross progenies produced in this study, there were two types of band patterns: a triplicate band pattern for both two alleles (f and a) and a single band pattern for one allele (a). A dosage effect of the GOT genes was clearly observed on zymograms of the triplicate band pattern: bands 2 and 3 were stained more intensely than band 1. These results indicated that genotypes of the triplicate and single band patterns showed, respectively, aaf and aa. This result allowed us to identify the alien chromosome assortment of hyper-diploids and hypo-allotriploids. Seventy out of seventy-nine aneuploids examined (88.6%) carried chromosome 6F. We were able to confirm this result by means of a simple karyotypic observation (Fig. 4).
 View Details | Fig. 3. Glutamate-oxaloacetate transaminase zymograms. Lanes (left to right): Allium fistulosum, shallot, monosomic additions (2n = 17), double monosomic additions (2n = 18) and hypo-allotriploids (2n = 21, 22, 23). aa and ff indicate homozygous genotypes. aaf shows the heterozygous genotype. The frequencies of chromosome numbers in respective generations are shown in the bottom half of the figure. |
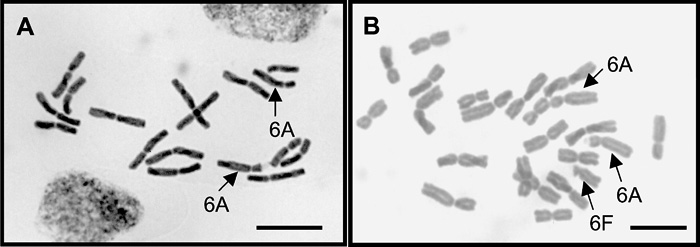 View Details | Fig. 4. Typical somatic metaphase chromosomes of monosomic addition (2n = 17) (A) and hypo-allotriploid (2n = 23) (B). Arrows point to the sub-telocentric chromosomes (6A, 6F). The monosomic addition does not possess the chromosome 6F. The hypo-allotriploid possessed the chromosome 6F. Scale bar = 10 µm. |
It was impossible to complete eight possible Allium cepa – A. fistulosum monosomic additions due to the insufficient number found in the BC2 progeny. The low seed fertility in the second backcross added to the difficulty. The main factor contributing to the low fertility could be the existence of deleterious alleles on a diploid background of shallot (A. cepa Aggregatum group). Because shallot has been asexually propagated for centuries, it is likely that mutation has produced deleterious alleles that are maintained in the heterozygote state. After sexual propagation, progeny plants homozygous for these deleterious alleles should show reduced vigor or perish, as shown in garlic, A. sativum L. (Jenderek, 2004), which is another Allium asexually propagated plant. To avoid this risk, the seed-propagated cultivar of bulb onion (A. cepa Common onion group) can be effectively substituted for shallot (A. cepa Aggregatum group) in the next backcrossing to produce the monosomic additions.
On the other hand, production of the hypo-allotriploids AAF-nF was easier than that of the monosomic additions. An increase in the number of alien chromosomes eases the effect of the deleterious gene found in shallot. Surprisingly, more hypo-allotriploids were obtained in the BC1 than in the BC2 progenies. We can offer the following interpretation of this phenomenon. Tashiro et al. (1981) found that abnormal chromosome pairings were among PMCs at metaphase-I in reciprocal amphidiploid hybrids between Allium fistulosum and shallot (A. cepa Aggregatum group): More than 90% of the pollen mother cells had 16 bivalents, though two univalents and one quadrivalent were also found in rare cases (< 6%). Chromatid non-disjunction might occur in the process of female gamete formation in amphidiploids since such an unexpected chromosome pairing is supposed to be also observed in embryosac mother cells. Several aneuploids (2n = 20, 23, 25, 26, 27, 30) found in the F2BC1 could result from fertilization between the aneuploid female gamete produced via abnormal pairings (e.g. n = 15) and the normal male gamete of A. cepa (n = 8) by chance. Thus, the amphidiploids have a great deal of flexibility in regard to the occurrence of backcross progeny with an unexpected chromosome number, including hypo-aneuploids with 23 chromosomes. We coined the term single-alien deletion (e.g., 2n = 3x-1 = 23) to designate a hypo-allotriploid in which one of the alien chromosomes are missing from the A. fistulosum complement in the diploid background of A. cepa. A complete set of all eight possible single-alien deletions would be very useful for locating the genes to specific chromosomes of A. fistulosum. Further cytogenetic studies including flow cytometric analysis and PCR-based DNA markers will be used to identify the complete single-alien deletion set among the remaining 60 F2BC1 plants. The rapid system for chromosomal identification with the help of flow cytometric analysis would be applicable not only for detecting numerical chromosome changes but also for identifying missing chromosomes in the alien deletions.
In the monosomics of several diploid species, the loss of a chromosome has a very drastic, sometimes lethal, effect on plant morphology (Khush, 1973; Singh, 2003). We observed that, in the juvenile stage, the loss of one chromosome at the triploid level was not as severe as the same loss at the monosomic level. Since this result is due to non-numerical changes of the Allium cepa chromosome complement consisting of a diploid background, it is unnecessary to consider the phenotypic expressions caused by recessive deleterious genes located on A. cepa chromosomes. Such an alien deletion is recommended as a tool for the chromosomal assignment of genes in the donor parent when the vegetative propagation plant is made to be the recipient parent. Furthermore, evaluating several effects of deleted chromosomes on the morphology and physiology of allotriploids could reveal the chromosomal locations of quantitative trait loci (QTL) in A. fistulosum. The simplicity of this technique can be compared with conventional QTL assignment by mapping. The unlinked QTL can be efficiently assigned to a chromosome using the single-alien deletions, although we may not be able to distinguish the linked QTLs due to their cumulative effect.
The authors wish to thank Miss S. Shinozaki, Mr. T. Utsunomiya and Miss N. Matsubara for their contributions to this study. The authors would like to thank Dr. M. J. Havey of USDA and the University of Wisconsin for critically reviewing this manuscript. The authors further express their gratitude to Ms. Lynn Hofstad of Hot Line for reviewing the manuscript regarding the English language.
|