| Edited by Hirokazu Inoue. Akio Toh-e: Corresponding author. E-mail: toh-e@biol.s.u-tokyo.ac.jp |
Anesthesia is a state that involves loss of consciousness, muscle relaxation, lack of memory of pain and insensitivity to pain. Although anesthetics are essential for millions of patients, the mechanisms of action and toxicity of these drugs remain poorly understood. Currently, approaches to understanding the action of anesthetics have been pursued in worms (Jayanthi et al., 1998; Sedensky and Meneely, 1987), flies (Allada and Nash, 1993), mice (Carla and Moroni, 1992; Marganella et al., 2005), humans (Rudolph and Antkowiak, 2004), plants (Nakao et al., 1998) and yeast (Palmer et al., 2002; Toh-e and Oguchi, 1998; Wolfe et al., 1999). Worms are a good tool for understanding anesthetics’ actions because they have a simple nervous system. The fly’s nervous system is more complicated than that of the nematode. Budding yeast (Saccharomyces cerevisiae) is the one of the best genetic model organisms in this research field because it possesses a high density of genetic markers and its genome has been completely sequenced. The fact that yeast has no nervous system is obviously a drawback to conducting studies related to the nervous system; however, it may be a useful experimental system to track direct cellular effects of anesthetics. Wolfe et al. (1999) found that a volatile anesthetic, isoflurane, inhibited the growth of yeast cells at a concentration approximately 10-fold higher than that required for anesthesia in mammals. The inhibiting concentrations followed the Meyer-Overton relationship (Humphrey et al., 2002); the potency of volatile anesthetics for ablating responses to noxious stimuli was correlated with their solubility in olive oil. In addition, the growth inhibition by the anesthetics is reversible and dose-dependent. These similarities indicate that the manner in which anesthetics inhibit yeast growth and induce anesthesia in mammals or other eukaryotes may be closely related, if not identical. Wolfe et al. (1999) also identified mutations, termed zzz1 and zzz4, which conferred on cells resistance to isoflurane. Zzz1p was found to be identical to Bul1p, which interacts with Rsp5p ubiquitin ligase. Zzz4p is identical to Doa1p/Ufd3p, which is involved in the degradation of ubiquinated proteins. These results suggest that the ubiquitin system plays a critical role in the response of yeast to anesthetics. Palmer and colleagues (2002) found that overproduction of amino acid permease Tat1p conferred resistance to anesthetics on yeast cells. This finding suggests that nutrient availability may be a critical factor in the response of the yeast cells to anesthetics.
In contrast to the mechanism of action of general anesthetics, the mechanisms of action of local anesthetics are relatively well understood. When local anesthetics reach and enter the ion channels of nerves, they interrupt nerve activity and a conduction block occurs. Tetracaine, a type of local anesthetic, blocks sarcoplasmic reticulum (SR) Ca2+ release channels (ryanodine receptors, RyRs) (Tinker and Williams, 1993).
The precise lesions and pathological characteristics produced by neurotoxicity of tetracaine are not well understood. However, several investigators have shown nerve injury in association with intrathecal injection of a high concentration of tetracaine. Adams et al. (1974) showed that intrathecal 2% tetracaine caused small foci of degeneration in the nerve roots of rabbits. Ready et al. (1985) showed that 8% tetracaine caused central necrosis within the spinal cord in rabbits. The immediate effects of administered tetracaine remain elusive.
Toh-e and Oguchi (1998, 1999) found that tetracaine inhibited yeast growth. To investigate the action of tetracaine in yeast, they isolated and characterized mutants hypersensitive to tetracaine. The LAS17 gene was found to encode a WASp (Wiskott-Aldrich syndrome protein) homologue that promotes actin filament assembly by activating the Arp2/3p complex and thereby spatially and temporally regulating the assembly of specialized actin structures (Lechler and Li, 1997; Madania et al., 1999). The LAS21 gene was found to be involved in glycosylphosphatidylinositol (GPI) anchor synthesis and in the stress response pathway (Toh-e and Oguchi, 1999). These studies emphasized that the yeast can be used for analyzing the local anesthetic action in spite of the absence of the nervous system in yeast. How tetracaine inhibits the growth of these mutants remains unknown.
In the course of our molecular genetic studies of local anesthetics with the yeast Saccharomyces cerevisiae, we found that the hypersensitivity to tetracaine and, simultaneously, temperature-sensitivity for growth of las24 mutants were due to mutations occurring in the KOG1 gene. Recently, Hall and colleagues (Loewith et al., 2002) reported that Kog1p, alias Las24p, associated with Tor1p (Target of Rapamycin) and with Tor2p. Tor1p coimmunoprecipitated with Kog1p, Tco89p and Lst8p, and Tor2p coimmunoprecipitated with Kog1p, Lst8p, Avo1p, Avo2p, Avo3p and Bit61p (Fadri et al., 2005; Loewith et al., 2002; Reinke et al., 2004). These coimmunoprecipitation studies, supplemented by depletion assays with genes under the control of a glucose-repressible promoter, led them to propose the existence of two TOR complexes, TORC1 and TORC2. Two protein complexes, Tor1p-Kog1p-Lst8p-Tco89p and Tor2p-Kog1p-Lst8p-Tco89p belong to TORC1. TORC2 is the protein complex Tor2p-Lst8p-Avo1p-Avo2p-Avo3p-Bit61p. Kog1p is contained specifically in TORC1. TORC2 may have distinct roles in growth control (Loewith et al., 2002). Schmelzle et al. (2004) demonstrated that TOR controled several cellular events via the Tap42/Sit4p pathway and the Ras/cAMP pathway. Since Las24p/Kog1p is a specific subunit contained in TORC1, it is possible to investigate the function of TORC1 by employing las24 mutants.
In this communication, we describe a study of TORC1 function in which we employed a las24-1 mutant that has a defect in a specific TORC1 component. Although the las24-1 mutant exhibited tetracaine-sensitivity as well as temperature-sensitivity, we think it is unlikely that the direct target of tetracaine is Las24p/Kog1p. Our analyses of the las24-1 mutant led us to hypothesize that the tetracaine-sensitive step may be a new downstream element of the TORC1 pathway.
The yeast strains used in this study are listed in Table 1. All the strains except Y06864 have the W303 background. The yeast strain carrying the myc-tagged GLN3 gene (GLN3-13xmyc) and that carrying the myc-tagged VSP10 gene (VSP10-myc) were constructed by a PCR-based procedure as described previously (Longtine et al., 1998). The LAS24-8xGFP and LAS24-3XFLAG strains were constructed by integrating the linearized plasmid pTA19 or pTA21 using SalI, respectively. The Npr1p-HA strains were constructed by integrating plasmid pTA32 linearized by SphI. The TOR1 deleted strain YTA1201 was created by integrating the PCR-amplified tor1::kanMX4 cassette from Y06864c.
 View Details | Table 1. Yeast strains used in this study |
Yeast cells were usually grown in YPD medium containing 2% glucose, 1% yeast extract (DIFCO) and 2% Polypepton (Nihon Seiyaku) or the synthetic complete (SC) medium described by Sherman (1986). Dropout medium was prepared by removing the appropriate supplements from SC. Standard methods for yeast genetics and molecular cloning were described by Sherman (1986). Tetracaine hydrochloride (Sigma) was stored at 200 mM in water and was added to growth medium at the indicated concentration. Plasmid DNAs were constructed and propagated in Escherichia coli DH5α (deoR endA1 gyrA96 hsdR17 (rk–mk+) recA1 relA1 supE44 thi-1 Δ(lac ZYA-argF)U169 φ80lacZΔM15 F– λ–). Yeast cells were grown at 25°C (permissive temperature) or 37°C (restrictive temperature) and E. coli cells were grown at 37°C.
The plasmids used in this study are listed in Table 2. Plasmids in the pRS series were described by Sikorski and Hieter (1989), while those in the pYO series were multicopy derivatives of pRS plasmids (Y. Ohya) and YEp24 was described by Botstein et al. (1979). The LAS24-expressing vectors pTA25 and pTA26 contain the PCR-amplified LAS24 promoter (~1 kbp), ORF and 3’-UTR (~500 bp). The amplified ~6.2 kbp KpnI-XhoI fragments were digested with KpnI and XhoI and cloned into pRS316 or pYO326 at the KpnI-XhoI sites. pTAs24, which was used for stabilizing the las24 mutants as described below, was created as follows. The ~300 bp LAS24 3’-UTR segments amplified by PCR using primers 5’-TATGTGCCATTTCTTTTTTT-3’ and 5’-ATCGATGTTAAAATGTTCCA-3’ were digested with BamHI and XhoI, and integrated into the BamHI-XhoI site of pRS303 to create pTAs24. The plasmids containing LAS24-8XGFP (pTA19) or LAS24-3XFLAG (pTA21) were p8XGFPIU and p3XFLAGIU expressing C-terminally GFP-tagged LAS24 or FLAG-tagged LAS24, respectively. The 1.7 kb HindIII-XbaI fragment containing the LAS24 ORF was generated by PCR using the primers 5’-CACAAAGCTTTGCTTGAATTAAGCGCCCAT-3’, and 5’-CACATCTAGAAAAATAATCAATTCTCTCGT-3’, and then was cloned into p8×GFPIU and p3XFLAGIU. P8XGFPIU is a YIplac211 (Gietz and Sugino, 1988) based vector containing an insert encoding the 8XGFP tag and the GAP-DH transcription terminator adjacent to the multiple cloning site. The p3XFLAGIU vector was created by introducing the PCR-amplified 3XFLAG epitope tag and the CYC1 transcription terminator cassette (Saeki et al., 2002) with artificially created BamHI and EcoRI sites at the 5’- and 3’-ends into the YIplac211 BamHI-EcoRI site. pNPR1-HA (pTA23) is a pTS901IU-based plasmid expressing the C-terminally HA-tagged NPR1. The 0.6 kb HindIII-XbaI fragment containing the NPR1 ORF was generated by PCR using the primers 5’-CACAAAGCTTTCGATTTTGGTGCTGCTGTG-3’ and 5’-CACATCTAGATTGATTATTTTGCTTTTTCT-3’, and then was cloned into pTS901IU to fuse the HA tag in frame. pTS901IU is a YIplac211-based vector containing an insert encoding the 5×HA tag and the GAP-DH transcription terminator adjacent to the multiple cloning site.
 View Details | Table 2. Plasmids used in this study |
To generate a temperature- and tetracaine-sensitive allele of las24, we adopted the PCR-based random mutagenesis method (Toh-e and Oguchi, 2000). The N-terminally truncated LAS24 ORF fragment (+201 ~ +4674), was amplified by PCR using primers 5’-CACATCTAGACCAGCCGATTACCGATTGGA-3’ and 5’-CACACTCGAGTCAAAAATAATCAATTCTCT-3’, and the resultant fragments were cloned into the XbaI-XhoI sites of pBluescript II KS(+). XbaI and XhoI sites were artificially created at the 5’- and 3’-termini of the DNA fragment, respectively. The resulting plasmid was used as a template for mutagenic PCR (Tintut and Gralla, 1995) using PCR buffer containing manganese. A pair of primers, the T3 promoter 5’-AATTAACCCTCACTAAAGGG-3’ and the T7 promoter 5’-TAATACGACTCACTATAGGG-3’ were used for PCR-based random mutagenesis amplification. The amplified fragments were digested with XbaI and XhoI, and cloned into the same sites of pRS306. The mutant library thus obtained was linearized with BamHI within the LAS24 ORF and integrated into the W303-1A strain. Ura+ transformants were checked for temperature-sensitivity at 37°C and tetracaine-sensitivity at 25°C. The resulting mutants were stabilized by removing the duplicated LAS24 sequence generated by the integration of mutant library DNA using plasmid pTAs24 (see above “Construction of plasmids”) after having been linearized with XhoI (Tohe and Oguchi, 2000).
Tolerance to tetracaine was determined in 2 ml of liquid YPD culture containing various concentrations of tetracaine. Cultures were started at an OD600 of 0.02 after diluting overnight cultures and incubated at 25°C with shaking. After 16 hr of incubation, the OD600 was determined with appropriate dilutions. A concentration of tetracaine that reduced growth by 50% was denoted as IC50.
To screen for high-copy suppressors of the las24-1 mutant, strain YTA1010 was transformed with a yeast genomic DNA library constructed in a 2 μ vector containing the URA3 marker. Ura+ transformants were selected at 25°C and then replica-plated onto SC-Ura plates and screened for the clones that grew at 37°C. Plasmids were isolated from the candidate clones, retransformed into YTA1010 and retested for growth at the restrictive temperature. The yeast DNA segments cloned in the plasmids were then subcloned and sequenced. The adenine residue of the putative translation initiation codon ATG was defined as +1 nucleotide.
Cells were lysed in the presence of an equal volume of lysis buffer and glass beads by three cycles of 30s vortexing-30s interval using a MULTI-BEADS SHOCKER (Yasui Kikai). Protease inhibitor cocktail (Roche Diagnostics) and phosphatase inhibitors (50 mM NaF, 100 mM β-glycerophosphate) were added to the lysis buffer (50 mM Tris-HCl [pH 7.5], 5 mM EGTA, 5 mM EDTA, 150 mM NaCl, 10% glycerol). The samples were denatured at 96°C for 5 min. Fifty micrograms of protein was loaded per lane for 7.5% or 5% (Gln3p-myc) SDS-PAGE. For immunoblotting, mouse anti-FLAG (clone M2: Sigma), mouse anti-alkaline phosphatase (clone 1D3: Molecular Probes), mouse anti-dolichol phosphate mannose synthase (clone 5C5: Molecular Probes), mouse anti-porin (clone 16G9: Molecular Probes), goat anti-Pma1p (clone yN-20: Santa Cruz), mouse anti-HA (clone 12CA5: Roche) and mouse anti-myc (clone Ab-1: CALBIOCHEM) antibodies were used for primary antibodies. The immune complexes were visualized with the enhanced chemiluminescence system (Amersham Biosciences), using horseradish peroxidase (HRP)-conjugated anti-mouse IgG (Amersham Biosciences) or HRP-conjugated anti-goat (CHEMICON) IgG antibody as secondary antibodies.
The rates of both incorporation of amino acids and protein synthesis were measured by the pulse labeling method (Uesono et al., 2004).
Yeast cultures were grown to an OD600 of 0.6 at 25°C and then 100 μg/ml cycloheximide was added. All subsequent steps were performed at 4°C. The cells were centrifuged and washed with polysome buffer (20 mM Tris-HCl [pH 7.4], 2 mM MgOAc, 100 mM KOAc, 100 μg/ml cycloheximide, 0.5 mM DTT). For temperature shift experiments, cells were incubated at a restrictive temperature (37°C) for 4 hr before harvesting cells. The cells were lysed in the presence of 1 volume of glass beads by three cycles of 20s vortexing and 40s interval using a MULTI-BEADS SHOCKER (Yasui Kikai). Lysates were cleared at 10,000g for 20 min in an Eppendorf microcentrifuge tube to give the final lysate. The lysate of 7 OD260 units were loaded onto a 10–45% linear sucrose gradient. The gradients were centrifuged in a P40ST rotor (Hitachi Koki) for 3 hr at 35,000 rpm, after which the gradients were collected using an ALC-20 automatic liquid charger (Advantec), and OD254 of fractions was monitored using an LKB UV monitor (Pharmacia).
For detection of GFP fluorescence, the cells expressing GFP-tagged Las24p were grown to early logarithmic phase in SC-Ura medium at 25°C. The cells were directly mounted and subjected to microscopic analysis. For visualizing actin, yeast cells were grown to early logarithmic phase at 25°C and shifted to and kept at 37°C for 4 hr, or treated with the indicated drugs for 1 hr. The cells were fixed directly in growth medium with formaldehyde (4% final concentration) for 10 min. The cells were collected and resuspended in phosphate-buffered saline (PBS) containing 4% formaldehyde. After incubation for 1 hr, the cells were washed three times with PBS and stained with rhodamine-conjugated phalloidin (Molecular Probes) as described (Adams and Pringle, 1991).
Subcellular fractionation by differential centrifugation was carried out as described by Becherer et al. (1996). For membrane association tests, the combined P13 and P100 pellet fractions were subjected to centrifugation at 100,000g at 4°C for 1 hr after with or without treatment with sodium chloride, sodium carbonate, urea, or Triton X-100. After centrifugation, supernatants were withdrawn, and pellets were resuspended in a volume of lysis buffer equal to the volume of the supernatant. Samples were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then subjected to Western blotting.
A high concentration of a local anesthetic, tetracaine, inhibited the growth of wild-type yeast. The minimum concentration of tetracaine that inhibited the growth of the wild-type cells (W303) was determined to be 2 mM on YPD solid medium or 4 mM in liquid medium (IC50 = 1 mM) (Fig. 1A and B). We then treated W303-1B cells with ethylmethane sulfonate and screened tetracaine-hypersensitive colonies on YPD plates containing 2 mM tetracaine. The resulting mutants were named local anesthetic sensitive (las) mutants (Toh-e and Oguchi, 1998). Among these mutants, in this study, we focused on LAS24.
 View Details | Fig. 1. Mutant strain las24-1 shows tetracaine sensitivity. (A) Tetracaine sensitivity of the las24-1 mutant. Wild type W303-1A (WT) and YTA1010 (las24-1) strains were streaked onto YPD plates with or without tetracaine. Plates were incubated at 25°C for 4 days. (B) Dose-dependent inhibition of yeast growth by tetracaine. Exponentially growing W303-1A (WT) and YTA1010 (las24-1) were diluted to OD600 of 0.01 in YPD medium containing the indicated concentration of tetracaine. Each culture was incubated with shaking at 25°C in the presence of the indicated amounts of tetracaine. At 16 hr of incubation, OD600 was determined after appropriate dilutions. The values are normalized to the OD600 of control cultures grown without tetracaine and given as percentages. |
The las24-e mutant (YAT1861), which was previously isolated after ethylmethane sulfonate mutagenesis, showed temperature- and tetracaine-sensitivity. In order to clone the gene responsible for the las24-e defect, we exploited the temperature-sensitivity of this mutant. A yeast genomic library based on YCUp4 (A. FUJITA) was introduced into the las24-e mutant, and screened for transformants growing at 37°C. We isolated one candidate plasmid. This plasmid also suppressed the tetracaine sensitivity of the las24-e cells. Then, a part of the nucleotide sequence of the insert derived from the yeast genomic DNA was determined. The sequence analysis indicated that the plasmid recovered from the transformant contained only one open reading frame (ORF) YHR186c on the segment of the yeast DNA cloned on the plasmid.
To confirm that the cloned DNA contains the gene responsible for the las24-e mutant phenotype, the YHR186c ORF was cloned into pRS306, and integrated into W303-1B. The resulting Ura+ transformants showed the wild-type phenotype with temperature- and tetracaine- sensitivity. Then the transformant was crossed withYAT1861 cells. The resulting diploid cells were sporulated, and tetrads were dissected. All asci showed 2 Las+ Ura+: 2 Las– Ura– segregation. This result supports that the notion that YHR186c is LAS24. Our sequence of LAS24, which is identical to the sequence determined for the YHR186c ORF on chromosome VIII by the yeast genome sequencing project, indicated that this gene encodes a 1558 amino acid protein. This gene has been identified by another screening and named KOG1 (Kontroller Of Growth) (Loewith et al., 2002).
To isolate various las24 mutant alleles, we adopted the PCR-based random mutagenesis method. Temperature-sensitive mutants were selected at 37°C and then checked for tetracaine sensitivity. All the temperature-sensitive las24 mutants showed sensitivity to tetracaine (data not shown). Intriguingly, the degree of temperature-sensitivity was parallel to that of sensitivity to tetracaine. Among these mutants, the las24-1 mutant showed the severest growth defect at a high temperature (35°C) and in YPD+tetracaine solid or liquid culture (data not shown). Therefore, the las24-1 mutant was used for further analysis. Sequence analysis showed that the las24-1 mutant contained a single point mutation T4637C, resulting in amino acid change I1546T, which is inside of the WD-40 repeats.
We examined whether the tetracaine resistance of the yeast cells could be influenced by a high copy of LAS24. Cells transformed with a high-copy-number plasmid carrying the LAS24 gene could grow in SC-Ura medium containing 4 mM tetracaine but cells with the empty vector could not (Fig. 2A). On the other hand, a high copy number of the las24-1 gene did not provide wild-type cells or las24-1 cells with tetracaine-resistance (Fig. 2B). Overexpression of las24-1 also could not rescue the las24-1 temperature-sensitivity (data not shown).
 View Details | Fig. 2. LAS24 is involved in tetracaine resistance. (A) Overexpression of LAS24 made yeast cells resistant to tetracaine. Wild type W303-1A strain transformed with a high copy number plasmid (pYO326 (YTA1020; vector) or pTA26 (YTA1042: LAS24)) was streaked across SC-Ura plates containing the indicated concentration of tetracaine, and incubated at 25°C for 4 days. (B) Overexpression of las24-1 can not rescue the temperature- and tetracaine-sensitivity. Ten-fold serial dilutions of overnight cultures were spotted onto SC-Ura plates with or without tetracaine to assess the growth at permissive or nonpermissive temperature. Wild type W303-1A (strain “WT”) and YTA1010 (strain “las24-1”) cells transformed with pYO326 (plasmid “vector”) or pTA30 (plasmid “las24-1”) were examined (YTA1020, YTA1022, YTA1021 and YTA1023). |
The las24-1 mutant cells were more sensitive to tetracaine (IC50 = 0.25 mM) than the wild-type cells (IC50 = 1 mM). The las24-1 mutant cells began to arrest at 3 hr after the temperature shift to a restrictive temperature (37°C) (Fig. 3A). The growth defect of the las24-1 cells at 37°C could be rescued by adding the osmotic stabilizer sorbitol to the medium (Fig. 3B). This phenotype is a feature of a defect in cellular integrity caused by stress at the plasma membrane and/or the cell wall (Evans and Stark, 1997; Reinke et al., 2004). Accordingly, we introduced several genes reported to be involved in the cell integrity pathway, i.e., PKC1, MPK1 and SSD1, into the las24-1 mutant one by one. Pkc1p is the yeast homologue of mammalian protein kinase C, an upstream regulator of a mitogen-activated protein kinase (MAPK) cascade that includes Mpk1p, one of the five MAPKs in yeast. Because MAPKs play key roles in maintaining cell wall integrity, yeast cells lacking PKC1 or MPK1 undergo cell lysis, and require an osmotic stabilizer for growth (Levin and Bartlett-Heubusch, 1992). SSD1 has been shown to play a role in a variety of cellular functions, including the maintenance of cell wall integrity. Each of PKC1, MPK1, and SSD1 was introduced into the las24-1 mutant in a low copy (in case of SSD1) or high copy (in case of MPK1 and PKC1) number plasmid under the control of their respective promoters. Transformants were then examined for temperature-resistant growth. Only SSD1 could partially restore the ability of the las24-1 cells to grow at a high temperature (Fig. 3C). SSD1 has also been implicated in calcium homeostasis (Tsuchiya et al., 1996). Then, we tested the effect of calcium ion on the las24-1 cells. The addition of 0.1 M CaCl2 could partially suppress the las24-1 temperature-sensitive growth phenotype (Fig. 3B). The addition of 0.1 M MgCl2 or 0.2 M KCl to the growth medium did not enable the las24-1 cells to grow at a high temperature (>35°C) but had no effect on the growth of wild-type cells at this temperature (data not shown), indicating that the rescue of temperature-sensitivity of the las24-1 mutant by CaCl2 is due specifically to calcium ion and not to chloride ion. On the other hand, none of these supplements rescued the growth of the las24-null mutant (data not shown). These results indicate that the function of Las24p partially overlaps with that of Ssd1p.
 View Details | Fig. 3. Growth characteristics of the las24-1 mutant. (A) Growth rate of wild type and las24-1 strains. At time zero, exponentially growing wild type W303-1A (WT) and YTA1010 (las24-1) cells at 25°C were diluted to an OD600 of approximately 0.06 in prewarmed YPD and shaken at 25°C or 37°C. At each indicated time point, OD600 was measured. (B) Effects of sorbitol and Ca2+ on the temperature-sensitivity of las24-1 cells. Wild type W303-1A (WT) and YTA1010 (las24-1) strains were streaked across the indicated plates, and were incubated for 3 days at the indicated temperatures. (C) SSD1 partially reversed the temperature-sensitivity of the las24-1 strain. pRS316 (vector), pTA25 (LAS24) or pFK4 (SSD1) was introduced into YTA1010 (las24-1) cells (YTA1092, YTA1025 and TYA1061). One of the resultant transformants was streaked across an SC-Ura plate and incubated at 25°C or 35°C for 3 days. |
To identify genes collaborating with the LAS24 gene, we screened high-copy suppressors of the temperature-sensitive growth phenotype of the las24-1 mutant. Seventy-six plasmids thus isolated were classified into four groups. The DNA segments possessing the suppressor activity were subcloned and sequenced. The responsible genes were found to be GIS3, ZDS1, TIP41 and LAS24 (Fig. 4A).
 View Details | Fig. 4. Phenotypic characterization of las24-1 strain. (A )Multicopy suppressors of the las24-1 strain. The las24-1 cells transformed with plasmids pYO326 (vector: YTA1021), pTA26 (LAS24: YTA1026), pTA31 (GIS3: YTA1032), pTA32 (ZDS1: YTA1033), pTA33 (TIP41: TYA1034), pSM1 (BCY1: YTA1035) or pd9 (PDE2: YTA1036) were streaked across SC-Ura plates. Plates were incubated at the indicated temperature for 3 days. (B) Drug sensitivity of las24-1 strain. Wild type (WT) and YTA1010 (las24-1) strains were streaked across YPD plates containing 6 mM caffeine or 5 ng/ml rapamycin, and incubated at 25°C for 3 or 4 days. |
GIS3 is a putative activator of PDE2 (Balciunas and Ronne, 1999). The PDE2 gene codes for high-affinity 3’,5’-cyclic AMP phosphodiesterase, which negatively regulates the cAMP dependent protein kinase A (PKA) pathway. The isolation of the GIS3 gene as a multicopy suppressor of las24-1 suggested that the RAS/cAMP pathway may negatively regulate the Las24p function directly or indirectly. To test this possibility, we examined the effect of overproduction of Pde2p and of Bcy1p, which encodes a regulatory subunit of PKA. As shown in Fig. 4A, the overexpression of either BCY1 or PDE2 suppressed the temperature-sensitivity of the las24-1 mutant. These results implied that the temperature-sensitive growth of the las24-1 mutant could be suppressed by inhibition of the RAS/cAMP pathway, and that activation of this pathway might aggravate the las24-1 mutant phenotype. In addition, the las24-1 mutant was found to be sensitive to caffeine (Fig. 4B). The site(s) of action of the agent are not well understood, but, one of the characterized targets of caffeine is cAMP phosphodiesterase (PDE), which is inhibited by caffeine (Brenner and Thoms, 1984). The observation that the las24-1 mutant was hypersensitive to caffeine seems consistent with the effect of caffeine on PDE, the inhibition of which might upregulate the RAS/cAMP pathway, thereby exacerbating the las24-1 mutant phenotype.
ZDS1 has been identified in a wide variety of genetic screens, implying that it is a multifunctional protein involved in a large number of seemingly unrelated intracellular processes (Bi and Pringle, 1996; McMillan et al., 1999; Mizunuma et al., 1998). For this reason, the relationship between Zds1p and Las24p function is hard to predict. However, Griffioen et al. (2003) showed that Zds1p controlled the RAS/cAMP pathway via effects on Bcy1p localization and cell wall integrity. Moreover, zds1 mutant cells are more sensitive to rapamycin than wild-type cells (Xie et al., 2005). These and our data suggest that overproduction of Zds1p might suppress the las24-1 mutation by somehow down regulating the RAS/cAMP pathway, thereby activating events downstream of the TOR pathway.
Tip41p is a Tap42p-binding protein that is a regulatory subunit of the type 2A (and 2A-like) protein phosphatases. Tap42p binds to protein phosphatase (PPase) catalytic subunits Sit4p or Pph21p/Pph22p and exerts a positive effect on the TOR pathway. Two interpretations have been proposed for the function of Tip41p; one is that Tip41p may interact with Tap42p and inhibit Tap42p function (Jacinto et al., 2001), and the other is that Tip41p may play an active role in transducing the signal between Tap42p and the protein phosphatase complex (Santhanam et al., 2004). To assess the function of Tip41p in the las24-1 mutant, multicopy TAP42 was expressed in the las24-1 mutant cells. We found that las24-1 temperature-sensitivity could not be suppressed by overproduction of Tap42p (data not shown). Next we focused on Cdc55p, which out-competes Tap42p for Pph21p/Pph22p (Jiang and Broach, 1999). As shown in Fig. 5, cdc55 mutation suppressed las24-1 temperature-sensitivity. Furthermore, the las24-1 mutation suppressed the cold-sensitivity of the cdc55 mutant at 15°C, indicating that las24-1 and cdc55 had antagonistic effects. These results can be interpreted as follows: in the las24-1 mutant, the formation of Tap42p-PPase might be stimulated by inactivation of Cdc55p and, in turn, the las24-1 mutation might promote the formation of non-Tap42p-PPase at a low temperature by reducing Tap42p-PPase under this condition. Overproduction of Tip41p probably modifies Tap42p so that Tap42p can efficiently form Tap42p-PPase. These considerations led us to think that the Tip41p is likely to stimulate Tap42p-PPase.
 View Details | Fig. 5. Double mutant las24-1 cdc55 cancelled the temperature-dependent growth defect but exacerbated the morphological defect of cdc55. (A) Growth of the las24-1 cdc55 double mutant at a lower and a higher temperature. Wild type W303-1A (WT), YTA1010 (las24-1), YTA1051 (cdc55) and YTA1052 (las24-1 cdc55) cells were streaked across YPD plates. Plates were incubated for 3 days at 25°C and 36°C and at 15°C for 6 days. (B) A Morphological defect in the cdc55 and las24-1 cdc55 strains. DIC photomicrographs of yeast cells in panel (A) are shown. All strains were grown at 25°C in YPD liquid culture to logarithmic phase and imaged. |
Las24p is an alias of Kog1p and is a unique component of TORC1 (Target Of Rapamycin Complex 1) (Loewith et al., 2002). In yeast, the immunosuppressive drug rapamycin binds to prolyl isomerase, FKBP or Fpr1p, and the complex of rapamycin-Fpr1p inhibits the TOR activity (Heitman et al., 1991). Yeast cells with no or weakened TOR function showed rapamycin sensitivity (Xie et al., 2005). Down regulation of Kog1p could lead to a hypomorphic effect on TORC1. To test this possibility, we examined the inhibitory effect of rapamycin on the las24-1 mutant. The growth of las24-1 cells, but not of the wild-type cells, was inhibited by the presence of 5 ng/ml rapamycin (Fig. 4B). These results indicate that Las24p plays an active role in the TOR pathway.
Since activating the Tap42 pathway or the down-regulation of the Ras/cAMP pathway suppressed the temperature-sensitivity of the las24-1 cells, we asked whether the same genes also suppressed the tetracaine-sensitivity of the las24-1 cells. The tetracaine-sensitivity of the las24-1 cells was not suppressed by overexpression of the GIS3, ZDS1, TIP41 or SSD1 gene (data not shown).
The results of the analysis of the multi copy suppressor of las24-1 led us to investigate TOR dependent events in the las24-1 mutant. For this we focused on two downstream proteins of the TOR pathway, Npr1p and Gln3p. NPR1 encodes Ser/Thr kinase, which controls the post-Golgi sorting and degradation of amino acid permeases. In yeast cells grown in good nitrogen sources, Npr1p is hyper phosphorylated via the TOR pathway, but in cells grown in poor nitrogen sources, Npr1p is dephosphorylated and activated by the TOR-modulated Sit4p phosphatase complex. TOR inactivation by rapamycin treatment also induced accumulation of dephosphorylated Npr1p (Schmidt et al., 1998). GLN3 encodes a GATA type transcription factor responsive to nitrogen starvation. The function of Gln3p is also regulated by its phosphorylation and intracellular localization in a TOR dependent manner (Beck and Hall, 1999).
We first examined Npr1p dephosphorylation. Wild-type and las24-1 mutant cells transformed with pTA23 were grown to early logarithmic phase at 25°C, and shifted to and kept at a non-permissive temperature (37°C) for 1 hr or the cultures of wild type cells at 25°C were treated with rapamycin for 30 min. Extracts were prepared and subjected to SDS-PAGE and Western blotting analysis (see Materials and methods). As shown in Fig. 6, Npr1p-HA was hyper phosphorylated in the wild-type cells at either 25°C or 37°C and the las24-1 cells at a permissive temperature, and was dephosphorylated in the wild-type cells treated with rapamycin and in the las24-1 cells shifted to 37°C. We next investigated Gln3p phosphorylation levels. Myc-tagged GLN3 was integrated into the wild-type cells and the las24-1 cells and examined as above. Gln3p-myc was phosphorylated in the wild-type and las24-1 cells at 25°C, and the wild-type cells at 37°C. Dephosphorylated Gln3p-myc appeared in the las24-1 cells at 37°C and the wild type cells treated with rapamycin (Fig. 6). Thus Las24p was found to be required to prevent the dephosphorylation of Npr1p and Gln3p. However, the phosphorylation states of Npr1p and Gln3p were not affected by treatment with a high concentration (>2 mM) of tetracaine (data not shown). Our observations suggest that Las24p plays an active role in the TOR-Tap42p pathway and is not a direct target of tetracaine.
 View Details | Fig. 6. Las24p is required for Npr1p and Gln3p phosphorylation. Wild type (WT) and YTA1010 (las24-1) cells with integrated Npr1p-HA (YTA1101 and YTA1102) or Gln3p-myc (YTA1103 and YTA1104) cells were grown to logarithmic phase at 25°C and then shifted to 37°C or kept at 25°C for 1 hr. YTA1101 and TYA1103 cells grown at 25°C were treated with 200 ng/ml rapamycin for 30 min. Dephosphorylated forms of Npr1p-HA and Gln3p-myc are indicated by arrows. Phosphorylated forms of Npr1p-HA (Npr1p(P)-HA) and Gln3p (Gln3p(P)-myc) are indicated by a bar (Npr1p) or by an arrow (Gln3p) |
To determine whether Las24p is required for protein synthesis, as suggested by the observations in the studies of the TOR pathway, we examined the effect of the las24-1 mutation on protein synthesis. The las24-1 strain YTA1010 was shifted to a non-permissive temperature and the levels of amino acids uptake and protein synthesis were determined at various time intervals after the temperature shift. At a non-permissive temperature, we observed a progressive decrease in the level of [35S]methionine incorporation (Fig. 7A). The levels of both amino acid uptake and protein synthesis in the las24-1 cells fell to ~50% of the wild type levels, and those levels of tor1 fell to ~45% and ~40% of the wild type levels, respectively, after 4 hr of incubation at a non-permissive temperature. The levels of incorporation in the las24-1 cells and the tor1 cells at a permissive temperature were almost equal to those of the wild type cells. These results suggest that Las24p is required for amino acid uptake and protein synthesis.
 View Details | Fig. 7. Las24p is required for translation initiation. (A) Time courses of the rates of methionine uptake and protein synthesis were determined as described in MATERIALS AND METHODS. Wild type W303-1A (WT) and YTA1010 (las24-1) cells were assessed for incorporation of [35S]methionine for 5 min at the indicated time points (0, 30, 60, 120, 240 and 480 min) after temperature shift. Cells were trapped on GF/B glass filters (Whatman) and washed with 10 ml of cold water 3 times (for measurement of amino acid uptake). [35S]methionine taken up by the cells was counted using an LS6000IC liquid scintillation counter (Beckman). Cells were similarly collected and washed with 10 ml of 10% TCA (TCA) 3 times and acid insoluble [35S]methionine was counted. Protein synthesis was expressed by [TCA insoluble count]/[amino acid taken up]. Data shown are the average of three independently performed experiments. (B) Polysome profiles of wild type, las24-1 and tor1 cells. Wild type W303-1A (WT), YTA1010 (las24-1), YTA1201 (tor1) cells were grown at 25°C and shifted to and kept at 37°C for 4 hr. Cultures kept at 25°C throughout the experiment were used as references. Rapamycin was added to the wild type culture growing at 25°C and the culture was incubated for 1 hr before harvesting (WT + rapamycin). In all cases, cycloheximide was added to prevent polysome “run-off”. The wild type strain was slightly stimulated with respect to polysome accumulation. The positions of 40S ribosomal subunits (40S), 60S ribosomal subunits (60S), and 80S monosomal ribosomes (80S) are indicated by arrows. The peaks of polysomal ribosomes (polysomes) are indicated by a bracket. |
To determine whether the inhibition of protein synthesis took place at the level of initiation or elongation, the polysome profile of the las24-1 strain was analyzed. To this end, the las24-1 cells were cultured at a non-permissive temperature (37°C) for 4 hr, at which time las24-1 had stopped growing. Cell lysates were prepared in the absence of the translation elongation inhibitor cycloheximide. No polysomes were present in the extract, and only a peak corresponding to 80S monosomes and ribosome subunits were visible (data not shown). Therefore, translation elongation was not affected in the las24-1 mutant at the non-permissive temperature.
Next, cells were treated with cycloheximide to prevent “polysome run-off” and subjected to polysome analysis. The profile of the las24-1 mutant cells was similar to that of the isogenic wild type cells (W303-1A) when grown at 25°C. However, when the cells were shifted and kept for 4 hr at 37°C, a progressive decrease of polysomes and a coincident increase in the 80S peak were observed in the las24-1 mutant extract but not in the wild type extract. The tor1 cells also ceased translation initiation at 37°C. A similar but less pronounced effect was observed in rapamycin-treated cells (Fig. 7B), as described in the report by Barbet et al. (1996). Altogether, these data suggest that Las24p is required for amino acid uptake and translation initiation and that since Las24p/Kog1p is a unique component of the TORC1 complex, amino acid uptake and translation initiation are specifically regulated by this TOR complex.
Previous studies have demonstrated that rapamycin-treated cells displayed a random distribution of actin (Wang and Jiang, 2003). This observation suggests that actin polarization is controlled by the TOR pathway. In the experiment using the Kog1p-depleted cells, it could not be clarified whether polarization of the actin cytoskeleton depended on the Las24p/Kog1p function (Loewith et al., 2002). If Las24p acts as a positive regulator of TOR, then the las24-1 mutation would result in the disorganization of the actin cytoskeleton. To test this possibility, we examined the localization of the actin cytoskeleton in the las24-1 mutant and wild type cells treated with rapamycin. Exponentially growing wild type and las24-1 cells at 25°C were shifted to 37°C or kept at 25°C for 4 hr, or wild type cells were treated with 200 ng/ml rapamycin for 1 hr. Cells were fixed, stained with fluorescent phalloidin, and observed under a fluorescence photomicroscope. In wild type cells at 25°C or 37°C, unbudded cells had an actin cap at the site of the incipient bud. In small and middle size buds, actin patches accumulated in the daughter cells, and actin cables running in parallel to the longitudinal axis of the cells were clearly visible. In large budded cells, actin patches were concentrated at the mother bud necks. Following treatment with rapamycin for 1 hr, on the other hand, actin cables disappeared and the normally asymmetric distribution of the actin patches was replaced with a randomized distribution pattern (Fig. 8). When grown at 25°C, the las24-1 cells displayed normal cell morphology and normal actin distribution. Following incubation at 37°C, however, the actin cytoskeleton was depolarized as in the cells treated with rapamycin (Fig. 8). Using the same experimental setting, we found the abnormal distribution of actin in the tor1 cells at a non-permissive temperature. These results suggest that the TORC1 complex is required for the polarized distribution of actin.
 View Details | Fig. 8. Las24p is required for the polarized distribution of actin. Exponentially growing cells were shifted from 25°C to 37°C for 4 hr, or treated with 200 ng/ml rapamycin for 1 hr. Wild type (WT), YTA1010 (las24-1), YTA1201(tor1), and rapamycin-treated wild type (WT + rapamycin) cells were fixed, stained with fluorescent phalloidin, and imaged in DIC or fluorescence mode. |
To examine the intracellular location of Las24p, we constructed a FLAG-tagged version of the gene. Three copies of the eight amino acid FLAG epitope were introduced at the C-terminus of Las24p (Table 2). FLAG-tagged LAS24 behaved the same as the wild type counterpart (data not shown). Monoclonal antibody (anti-FLAG M2) recognized approximately 180kDa Las24p-FLAG protein on SDS-PAGE (Fig. 9A).
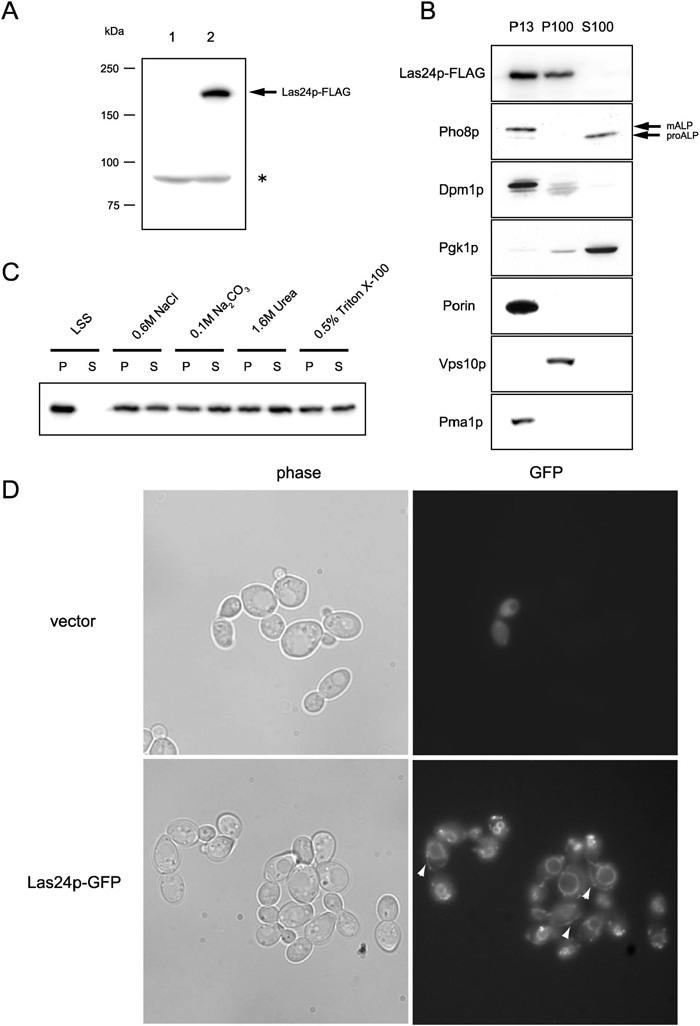 View Details | Fig. 9. Las24p is membrane protein that is localized at vacuoles, the plasma membrane and small vesicle structures. (A) Anti-FLAG antibody (M2) recognized approximately 180kDa Las24p-FLAG proteins. Wild type W303-1A cells transformed with control p3XFLAGCU vector (YTA1041: lane 1) or Las24p-3XFLAG expressing cells (YTA1042: lane 2) were lysed and subjected to Western analysis. (*) non-specific band. (B) Las24p is distributed in membrane fractions. Cleared cell lysate from the strain (YTA1042) with integrated LAS24-3XFLAG and VPS10-13Xmyc was fractionated by centrifugation at 13,000g (P13), and then at 100,000g (P100 and S100). Fractionation markers: Pho8p, vacuole (mature ALP; mALP) and cytosol (precursor ALP; proALP); Dpm1p, ER, Pgk1p, cytosol; porin, mitochondoria; Vps10p, Golgi; and Pma1p, plasma membrane. (C) Differential extraction of Las24p. The P13 and P100 membrane fractions from the YTA1042 cells were combined and incubated with the indicated chemicals for 1 hr at 4°C, and then centrifuged for 1 hr at 100,000g. (D)Las24p is localized to vacuoles, the plasma membrane, and small vesicle structures. Yeast cells transformed with control plasmid p8XGFPCU (YTA1044: vector) and a Las24p-8XGFP-expressing strain (YTA1045: Las24p-GFP) were imaged in the DIC or fluorescence mode. Las24p-8XGFP fluorescence at the plasma membrane is indicated by white arrow-heads. |
Las24p-FLAG expressing cell extracts were subjected to differential centrifugation to separate various organelles as described by Becherer et al. (1996). Lysates were fractionated by centrifugation into a 13,000g pellet (P13), 100,000g pellet (P100) and 100,000g supernatant (S100). Each fraction was examined for the presence of the marker proteins ALP (vacuole membrane), Dpm1 (endoplasmic reticulum), Pgk1p (cytosol), porin (mitochondria), Vps10p-myc (Golgi) and Pma1p (plasma membrane) by SDS-PAGE and immunofluorography. As shown in Fig. 9B, most of Las24p was associated with P13 and P100 fractions. Large membranes in structures such as vacuoles, the plasma membrane, endoplasmic reticulum and mitochondoria are known to be associated mainly with P13. Cytosolic protein Pgk1p and precursor ALP (proALP) are known to be predominantly contained in the S100 fraction and Golgi protein Vps10p-myc in the P100 fraction. Las24p-FLAG was detected in the P13 and P100 fraction, suggesting that it is likely to be localized in large and small membrane organelles.
We examined the nature of Las24p-FLAG association with the pellet fraction (P13 and P100). For this, the combined pellet fraction (P13 + P100) was treated with 0.6 M NaCl, 0.1 M Na2CO3 (pH11), 1.6 M Urea or 0.5% Triton-X100. In this test, NaCl, Na2CO3 and urea solubilize peripheral membrane proteins, and Triton X-100 solubilizes transmembrane proteins (Wang et al., 2005). As shown in Fig. 9C, all of the treatments solubilized about half of Las24p-FLAG, indicating that Las24p-FLAG in particulate fractions does not appear to be an integral membrane protein, but more likely exists at the surface of membranes as a component of some protein complexes.
Next, we examined the intracellular distribution of Las24p-GFP in living cells using fluorescence microscopy. First, we created yeast cells expressing GFP-tagged Las24p driven by the authentic LAS24 promoter, but GFP fluorescence was not detected. Then, we created a fusion protein containing eight tandemly repeated GFP (8XGFP) at the Las24p C-terminus by integrating plasmid pTA19, and the resulting strain was employed for microscopic analysis. LAS24-8XGFP cells grew as robustly as the wild type cells. Moreover, the multi tandem repeat GFP did not create artificial signals and its localization was dependent on the fused protein (Yoshida et al., 2003; our unpublished results). All these results indicate that Las24-8XGFP could be used as a tracer for monitoring Las24p localization. GFP fluorescence was found at vacuole membranes and punctate structures like the Golgi or small vesicular organelles in the LAS24-8xGFP cells but not in the cells with the control vector (CEN 8XGFP) (Fig. 9D). GFP fluorescence was also observed at the plasma membrane in some cells (Fig. 9D: white arrow-heads). These results indicate that Las24p is localized at large (vacuole, plasma membrane) and small membrane organelles, and is part of a protein complex.
Las24p, alias Kog1p, is a TOR-binding protein that is widely conserved in many organisms. In mammals, the Las24p/Kog1p homologue raptor (regulatory associated protein of mTOR) binds to mTOR protein at its N-terminus RNC (raptor N-terminal conserved) domain, and regulates translation initiation via translation elongation initiation factor eIF4E (Hara et al., 2002; Kim et al., 2002). Budding yeast Las24p/Kog1p and its homologues contain the WD-40 repeats at the C-terminus, and might bind to other protein(s) at the WD-40 domain. For this reason, Las24p/Kog1p (or with its binding protein(s)) is predicted to control TOR function(s). Budding yeast, in contrast to other eukaryotes, has two TOR proteins, Tor1p and Tor2p. Previous studies revealed that Las24p/Kog1p was present in only the TORC1 complex (Loewith et al., 2002; Wedaman et al., 2003). In yeast and other organisms, Tor proteins are components of two structurally and functionally distinct complexes, TORC1 and TORC2. Recent studies demonstrated that TORC1 contains Tor1/2p, Lst8p, Tco89p and Kog1p, and mediates rapamycin-sensitive Tor signaling pathways, such as protein synthesis, ribosome biogenesis and autophagy (Loewith et al., 2002; Reinke et al., 2004). On the other hand, the TORC2 complex was found to contain Tor2p, Lst8p, Bit61p and Avo1-3p, and mediates rapamycin-insensitive Tor signaling pathways such as the normal distribution of the actin cytoskeleton (Fadri et al., 2005; Loewith et al., 2002; Reinke et al., 2004). In many previous studies, characterization of Las24p/Kog1p/raptor family protein was carried out using gene knock-out cells (Hara et al., 2002; Kim et al., 2002; Loewith et al., 2002). However, temperature-sensitive mutants are easy to handle and their characterization may allow us to identify a domain with a specific function. With these considerations in mind, we attempted to characterize the las24-1 mutant in this study.
Since Las24p/Kog1p is a specific subunit contained in TORC1, it is possible to investigate the function of the rapamycin-sensitive TOR pathway mediated by TORC1 by employing the las24-1 mutants. A similar line of experiments were done by Reinke et al. (2004) using the tor1 and tco89 genes, specific components of TORC1. They demonstrated that tor1 and tco89 strains displayed a defect in cell wall integrity, and their phenotypes were correlated with those of mutants in SSD1 and other unknown gene(s). Tor1p is a major Tor protein in the TORC1 complex, but Tor2p contributes in small part to the TORC1 complex. Therefore, Tor1p alone cannot represent the TORC1 complex. Indeed, Tco89p is a specific component of the TORC1 complex. This protein is dispensable, and disruption of the TCO89 gene was used for analysis of its function. The LAS24/KOG1 gene is an essential gene and a temperature-sensitive allele of this gene would be a promising tool for analysis of the TORC1 pathway. We could confirm that cell integrity was under the control of the TORC1 pathway and this pathway had some functional connection with the SSD1 gene, as described in the analysis of tco89Δ.
Phosphorylation of Npr1p and Gln3p and translation initiation were inhibited by treatment with rapamycin, indicating that these are under the control of the rapamycin-sensitive pathway, but the contribution of the TORC1 pathway in these phenomena remains elusive. Thus we tested these responses in las24-1 cells. In las24-1 cells, dephosphorylated Npr1p and Gln3p were accumulated and translation initiation ceased at a non-permissive temperature, indicating that regulation of the phosphorylation of Npr1p and Gln3p and translation initiation are under the control of the TORC1 complex and that Las24p/Kog1p acts as an activator of the TORC1 complex. Schmelzle et al. (2004) presented a model in which the rapamycin-sensitive TOR signaling pathway bifurcates into the Tap42/Sit4p pathway and RAS/cAMP pathway. Evidence supporting this model included the fact that activation of the RAS/cAMP pathway conferred rapamycin-resistant growth only in the gln3 gat1 background, and many of TOR-mediated responses are not dependent on the Tap42/Sit4p pathway but on the RAS/cAMP pathway. Intriguingly, temperature-sensitivity of the las24-1 mutant was suppressed by down regulation of the RAS/cAMP pathway by overexpression of PDE2 or BCY1 even in a GLN3 background. Obviously, further work should be done to reconcile the contradictory effect of the Ras/cAMP pathway on the TOR pathway. Our tentative explanation is that the Ras/cAMP pathway has dual functions; (i) the RAS/cAMP pathway may function as a down stream effector of the TOR pathway and (ii) the RAS/cAMP pathway may regulate the TORC1 complex, in which interaction between Tor1p and Las24-1p may be inhibited by excessive activity of PKA.
Moreover, overexpression of TIP41 also restored the temperature-resistance of the growth of the las24-1.mutant. Tip41p was identified as a Tap42p-interacting protein and loss of Tip41p function induced rapamycin-resistance, suggesting that Tip41p acted as an inhibitor of Tap42p in the rapamycin-sensitive TOR signaling pathway (Jacinto et al., 2001; Xie et al., 2005). However, our result indicated that Tip41p played a positive role in the Las24p/Kog1p-mediated TOR pathway. Santhanam et al. (2004) demonstrated that tap42(Ts) tip41 double mutants grew more slowly than the isogenic tap42(Ts)TIP41 parent at both permissive and semi permissive temperatures in the absence of rapamycin. Thus, Tip41p may function differently in the presence and absence of rapamycin.
It has been believed that regulation of the actin cytoskeleton is a Tor2p-specific event and that Tor1p does not have a role in this phenomenon. This model is supported by the fact that a defect in the actin cytoskeleton is associated only with a mutation in Tor2p and is not suppressed by the presence of Tor1p. Previous studies (Loewith et al., 2002) could not show clearly whether Kog1p can mediate actin polarization, because the Kog1p-depleted cells induced vacuole expansion and disturbed polarized actin distribution. In contrast, las24-1 cells maintained normal vacuole morphology at both the permissive and non-permissive temperatures. Thus, we were able to investigate the state of the actin cytoskeleton in the las24 cells. In contrast to the previously reported results (Loewith et al., 2002), our results showed that the distribution of actin cytoskeleton was dependent on the Las24p/Kog1p function. Therefore, it is obvious that the regulation of the actin cytoskeleton is not carried out by Tor2p alone. This conclusion was supported by two independent studies. One is the result of Wang and Jiang (2003). They demonstrated that rapamycin-treated cells displayed a random distribution of actin, whereas cells expressing TOR1S1972R, a rapamycin-resistant TOR1 gene, showed the normal actin distribution. This result indicates that Tor1p is also involved in regulating actin distribution. The other is the result of Uesono et al. (2004). They showed that cycloheximide-treated cells displayed abnormal distribution of actin. This data suggests that translation is required for actin polarization. In the las24-1cells and the tor1 cells, translation ceased at a restrictive temperature. For these reasons, rapamycin treatment or loss of Las24p (and probably Tor1p) function directly or indirectly induced random distribution of the actin cytoskeleton via the control of translation.
Previous studies of Tor1/2p localization have led to a variety of conclusions about the identity of the membranes with which Tor is associated: Cardenas and Heitman (1995) reported that Tor2p associated with vacuolar membranes and Kunz et al. (2000) reported that Tor1/2p associated with the plasma membrane and with an unidentified membrane compartment. Lst8p, containing WD-40 repeats, also a component of TORC1, and of TORC2 as well, was associated with endosomal/Golgi compartment (Chen and Kaiser, 2003). Tco89p was localized at vacuole membranes (Reinke et al., 2004). These results, sometimes conflicting with each other, showed some features of the distribution of Tor proteins in the yeast cells. By biochemical fractionation and the cytological method using GFP-fusion constructs, we found that Las24p, and therefore the TORC1 complex, was associated with membranes and appeared to be localized to the vacuole membrane, plasma membrane and small particulate structures. The complicated pattern of distribution of Las24p/Kog1p may reflect diverse functions of the TORC1 complex in yeast cells. Another level of complexity is produced by the fact that the TORC1 complex contains different molecular species; one containing Tor1p and the other Tor2p. Furthermore, according to the results of proteomic quantification of protein levels of budding yeast, there are several-fold differences among the relative abundance of subunits of the TORC1 complex (Ghaemmaghami et al., 2003). Therefore, previous studies of the localization of the TORC1 complex using subunits other than Las24p/Kog1p were unable to specifically detect all the locations of the TORC1 complex in the cell. Tco89p is a specific subunit of the TORC1 complex; however, since this subunit is nonessential, the TORC1 complex without this subunit can exist as a functional TORC1 complex. In contrast, since Las24p/Kog1p is an essential and specific component of the TORC1 complex, the localization of this subunit must correspond to the whole localizaion of the TORC1 complex. From these considerations, it is reasonable to assume that our demonstrated localization of the Las24p subunit includes that of other subunits of the TORC1 complex.
We have initiated molecular genetic studies aimed at elucidating the mechanism(s) of action of the local anesthetic tetracaine using the model eukaryote S. cerevisiae. The growth of yeast cells is inhibited by this local anesthetic agent. Inhibition of the growth of yeast cells by the anesthetic required >2 mM tetracaine. In rat or rabbit, this concentration of tetracaine causes profound damage to nerve cells (Adams et al., 1974; Ready et al., 1985; Takenami et al., 2000). One of the mutations that rendered yeast cells sensitive to tetracaine was found to occur in a nuclear gene, LAS24. Overexpression of LAS24 conferred tetracaine-resistance on yeast cells, suggesting that the quality and quantity of Las24p is important for resistance to this agent.
However, we do not think that Las24p/Kog1p is the target of the tetracaine action. The genes that can suppress the temperature-sensitivity of the las24-1 mutant did not confer the tetracaine-resistance on the las24-1 cells. Furthermore, Tor inactivation by rapamycin and by the las24-1 mutation induced dephosphorylation of Npr1p and Gln3, two factors downstream of the Tor pathway; however, in the las24-1 cells treated with tetracaine at a permissive temperature, did not dephosphorylate these proteins, indicating that tetracaine does not inactivate the TORC1 complex. Accordingly, we propose the existence of a tetracaine-sensitive effector that acts downstream of the TORC1 complex pathway.
|