| Edited by Koji Murai. Shigeo Takumi: Corresponding author. E-mail: takumi@kobe-u.ac.jp |
Abiotic stress conditions such as cold and drought seriously affect plant growth. Plants exposed to LT and drought conditions exhibit numerous physiological and developmental changes with significant alteration of gene expression profiles. In Arabidopsis, a number of genes are induced by both LT and dehydration however, some are responsive to LT while others are affected by dehydration (Seki et al., 2002). This observation suggests that various transcriptional regulatory mechanisms act in the LT and drought stress signal transduction pathways. Abscisic acid (ABA) is one of key plant hormones that is responsive to environmental stress conditions. Exogenous application of ABA induces a number of genes that respond to LT and drought stress (Thomashow, 1999). On the other hand, some genes that are induced by LT and drought do not respond to exogenous ABA treatment. Therefore, it is likely that both ABA-dependent and ABA-independent signal transduction pathways are operative in the LT and drought responses of plant cells (Thomashow, 1999; Shinozaki and Yamaguchi-Shinozaki, 2000).
Expression of Cor/Lea genes in plants is controlled through both the ABA-dependent and ABA-independent pathways. Significant research results on the Cor/Lea gene regulation are available for the dicotyledonous model plant Arabidopsis. A conserved C-repeat (CRT)/drought-responsive element (DRE), (G/A)CCGAC, is found as a cis element in the promoter region of the Arabidopsis Cor/Lea genes (Yamaguchi-Shinozaki and Shinozaki, 1994; Baker et al., 1994). It has been reported that Arabidopsis C-repeat binding factor (CBF)/DRE-binding protein1 (DREB1) and DREB2 recognize the CRT/DRE motif and belong to the ethylene-responsive factor (ERF)/APETALA2 (AP2) family (Stockinger et al., 1997; Liu et al., 1998). The CBF/DREB1 and DREB2 gene expression can be induced either by LT or by drought stress, and both responses are ABA-independent. Thus, cross-talk between LT and dehydration stress response occurs at the transcription level through binding of the CBF/DREB transcription factors to the CRT/DRE cis element in the ABA-independent pathway of Arabidopsis (Shinozaki and Yamaguchi-Shinozaki, 2000). Expression of a CBF4 gene (another member of the CBF/DREB family), however, is up-regulated by drought stress through the ABA-dependent pathway (Haake et al., 2002). Because a number of LT and drought inducible genes contain both CRT/DRE and ABA-responsible element (ABRE) motifs in their promoters, these cis elements are considered to function independently. A recent study on the Arabidopsis rd29A promoter showed that the CRT/DRE motif functions cooperatively with ABRE as a coupling element in the ABA-responsive gene expression in response to drought stress (Narusaka et al., 2003). The CBF4 may function in the slow response to drought that relies on the accumulation of ABA (Shinozaki et al., 2003). These regulatory pathways identified in Arabidopsis provide considerable information in understanding the LT/drought signal response in monocotyledonous crop species.
In cereals, transcription factors binding to the CRT/DRE motif have been identified in maize, rice and wheat. So far five cDNAs for CBF/DREB homologs have been isolated in rice, and the OsDREB1A and OsDREB2A have been shown to specifically bind to the CRT/DRE (Dubouzet et al., 2003). Expression of the OsDREB1A and OsDREB1B was induced by LT, whereas expression of OsDREB2A was induced by drought and high-salt stress. The different expression patterns of rice OsDREB1 and OsDREB2, under different stress conditions, are basically similar to those of Arabidopsis DREB genes. Only OsDREB1C has been shown to be constitutively expressed in rice plants. On the other hands, maize DBF1 and DBF2 bind to the CRT/DRE cis elements in the maize rab17 gene promoter, and expression of the DBF1 gene was reported to be induced by drought, salt and ABA (Kizis and Pages, 2002). Wheat TaDREB1 encoding a CRT/DRE-binding protein is also up-regulated by drought, LT and ABA (Shen et al., 2003). Expression patterns of these transcription factor genes of maize and wheat are unique in their ABA-responsiveness, as compared to most of the rice and Arabidopsis CBF/DREB family genes. Recent studies show that cereal plants possess a unique transcription factor with a binding motif distinct from CRT/DRE besides the characteristic expression patterns. Barley HvDRF1 gene belongs to the CBF/DREB subfamily of the ERF/AP2 family (Xue and Loveridge, 2004). The HvDRF1 transcription factor is able to bind to the CRT/DRE motif, but preferably to a CT-rich element (T(T/A)ACCGCCTT) in the HVA1 promoter. The core sequence of the CT-rich element is called a DRF1E motif (ACCGCC). The HvDRF1 gene is transcriptionally enhanced by ABA, drought and NaCl to produce three different transcripts, HvDRF1.1, HvDRF1.2 and HvDRF1.3, by alternative splicing (Xue and Loveridge, 2004). Out of the three transcripts, HvDRF1.2 mRNA is the most abundant transcript form and encodes 71 amino acids lacking any ERF/AP2 domain. Both HvDRF1.1 and HvDRF1.3 transcripts encode an ERF/AP2 transcription factor activating the HVA1 expression.
In the present study, we report the isolation of two cDNA clones encoding a wheat DREB2 homolog. The Wdreb2 genes showed high homology to the HvDRF1 (Takumi et al., 2003) and were identical to the TaDRF1 reported by Xue and Loveridge (2004). One of the two cDNA clones contained an internal insertion. The insertion caused frame shift and produced a stop codon to encode truncated polypeptide without any ERF/AP2 domain, which resembled to the HvDRF1.2 transcript. A detailed expression analysis of the Wdreb2 gene, in response to abiotic stress, and the differential accumulation of alternatively spliced transcripts is presented.
Two cultivars of common wheat (Triticum aestivum L., genome constitution AABBDD), a winter-type ‘Mironovskaya 808 (M808)’ and a spring-type ‘Chinese Spring (CS)’, were used throughout the study. A cultivar of tetraploid wheat, T. durum cv. ‘Langdon’ (AABB), was additionally used for studying the Wdreb2 copy number. A nulli-tetrasomic series of CS (Sears, 1966) was used for chromosome assignment. Each line of the nulli-tetrasomic series lacks a given pair of homoeologous A, B or D genome chromosomes (nullisomic condition) that are replaced by the corresponding homoeologous chromosome pair (tetrasomic condition). The ditelosomic lines of CS (Sears and Sears, 1978) of the homoeologous group 1 chromosomes were also used for chromosome assignment. Each line of the ditelosomic series lacks a chromosome arm.
Wheat seedlings were grown under standard conditions according to Kobayashi et al. (2004). For analysis of dehydration tolerance, 7-day-old seedlings (n = 20) were cold-acclimated at 4°C for 3 days or sprayed with 20 μM ABA solution containing 0.1% (w/v) Tween 20, and then removed from the soil and kept on dry filter paper for 4 h under the standard conditions. For analysis of salt tolerance, 7-day-old seedlings (n = 20) were cold-acclimated at 4°C for 3 days, removed from the soil, and then the roots were soaked in 0.4 M NaCl solution for 1 week. For the control, the seedlings without any pre-treatment were directly treated with drought or salt stress. Stress-treated seedlings were transferred back to the standard conditions, and at the 7th day of the transfer, number of surviving seedlings was recorded. The means with standard error were calculated based on 3 independent experiments.
From an EST library constructed using developing caryopses of CS (T. Nakamura, unpublished result), an EST clone designated as Wdreb2 was selected that showed high homology to the Arabidopsis DREB2 gene. Based on the sequence, the following gene-specific primer was designed for determining the 5’ end of the Wdreb2 cDNA; 5’-GTCATAAGCACGGGCAGCATCCTCAGCA-3’. Rapid amplified cDNA end (RACE)-PCR was conducted using a Marathon cDNA Amplification Kit (CLONTECH). Single-stranded cDNA for the RACE-PCR was produced using the total RNA extracted from cold-acclimated seedling leaves of M808. To isolate the complete open reading frames (ORFs) of the wheat Wdreb2, reverse transcriptase PCR (RT-PCR) was conducted with the following primer set using the total RNA as template: 5’-AAGAAAACAGGCGACAAGAT-3’ and 5’-ACGAAGCACAAAAAACTAGC-3’. The amplified cDNA fragments were cloned into pGEM-T vector (Promega, USA) and their sequences were determined by the automated fluorescent dye deoxy terminator cycle sequencing system using ABI PRISM™ 310 Genetic Analyser (PE Applied Biosystems, USA). Nucleotide sequences of the cDNAs and their deduced amino acid sequences were analyzed by DNASIS (Hitachi, Tokyo, Japan). Amino acid sequence homology was searched with the BLAST algorithm (Karlin and Altschul, 1993) and a multiple alignment was calculated according to Waterman (1986).
To confirm the nuclear localization of the WDREB2 protein, the SalI and NcoI linkers were added to the ORF of the Wdreb2 cDNA without the stop codon. The high-fidelity DNA polymerase KOD-plus (TOYOBO, Japan) was used for the PCR amplification, and the PCR product was cloned into SalI and NcoI site of the sGFP(S65T) vector (Chiu et al., 1995) to produce a fusion protein construct with the green fluorescent protein (GFP). The GFP construct was introduced into onion epidermal cells via particle bombardment for transient expression analysis. The bombardment conditions were according to Takumi et al. (1999). Fluorescence of the DAPI and GFP images were observed under fluorescence light microscopy BX60 (Olympus, Japan). The GFP fusion construct, NLS, known to target into the nuclei was also used as a positive control.
For genomic Southern blot analysis, total DNA extracted from CS and ‘Langdon’ was single-digested with DraI, EcoRI, HindIII, BamHI and EcoRV. Total DNAs from nulli-tetrasomic and ditelosomic series of CS were digested with BamHI. The digested DNA was fractionated by electrophoresis through 0.8% agarose gel and transferred to Hybond N+ nylon membranes (Amersham Bioscience, Piscataway, NJ). The Southern blots were hybridized with the 32P-labeled Wdreb2 cDNA clone as a probe. Washing conditions were 2 × SSC and 0.1% SDS followed by 0.1 × SSC and 0.1% SDS. Probe labeling, hybridization and autoradiography were performed according to Takumi et al. (1999).
For studies of gene expression, 7-day-old seedlings of M808 and CS grown under the standard conditions were transferred to 4°C and kept for the indicated periods under the standard light conditions. Seven-day-old seedlings were also treated with a solution containing 20 μM ABA or 0.4 M NaCl by foliar spray or dehydrated on dry filter papers in a desiccator for 4 h. Total RNA was extracted by guanidine thiocyanate method from the aboveground tissues and also from the seedling leaves. RNA (15 μg) was fractionated by electrophoresis through 1.2% formaldehyde/agarose gel and transferred onto the Hybond N+ nylon membranes. RNA blots were hybridized with the 32P-labeled whole sequences of the Wdreb2, Wdhn13, Wrab17, Wrab18 and Wrab19 cDNA clones as probes. Wdhn13 encodes a LEA D-11 DHN (dehydrin) protein responsive to both LT and dehydration stresses (Ohno et al., 2003). Three Wrab genes were previously characterized and their LT- and dehydration-responsive expression has been confirmed (Tsuda et al., 2000; Kobayashi et al., 2004). Probe labelling, hybridization, washing and autoradiography were performed in the same way as for Southern blot analysis. Because of the low levels of the Wdreb2 expression, the amount of their transcripts was determined by RT-PCR analysis using a first strand cDNA synthesis kit (TOYOBO, Japan). The total RNA was treated with DNaseI to remove the contaminated DNA. RT-PCR with the gene-specific primer set resulted in amplification of the single fragment for Wdreb2. The amplification at fewer cycles was in the exponential range of amplification. For RT-PCR analysis of the Wdreb2 expression, 31 amplification cycles was in the exponential range. As an internal control, a fragment from the wheat ubiquitin gene (Ubi-1) was amplified. Primer information for Ubi-1 and the PCR condition were according to Murai et al. (2003).
For studying light/dark response of Wdreb2, 7-day-old seedlings were grown for 0–8 days at 4°C under different light/dark regimes (see Results), and total RNA was extracted at different time points for northern blot analysis.
To amplify the internal region of the Wdreb2 ORF, the following primer set was designed; 5’-AAGAAAACAGGCGACAAGAT-3’ and 5’-GTTGTTGGTTCACTTCTTTC-3’. The Wdreb2 genomic sequences and the partial Wdreb2 cDNA fragments were amplified, cloned into the pGEM-T vector and sequenced. The obtained sequence data were analyzed by the DNASIS software and compared. Based on the exon/intron structure and sequences of spliced sites, three splicing product-specific upper primers were designed as follows; 5’-TCTATTCTGCGGAAAAAGCG-3’ for Wdreb2α, 5’-CAAACCAATAGGAAAAAGCG-3’ for Wdreb2β and 5’-CTCCAGCCTGGAAGGAAAAA-3’ for Wdreb2γ. Spliced product-specific band was amplified by RT-PCR with the specific upper primer and 5’-GTTGTTGGTTCACTTCTTTC-3’ lower primer and separated by electrophoresis on the 1.5% agarose gel.
The HybriZAP-2.1 Two Hybrid system (STRATAGENE, USA) was used to investigate trans-activation ability of the two WDREB2 proteins. The entire ORF sequences of the Wdreb2β and Wdreb2γ cDNAs were amplified with the following primer sets containing the EcoRI and SalI linkers; 5’-GGGAATTCGCGGACAAGATTGCGAAC-3’ and 5’-GCGTCGACACTTCTTTCCATCGCCTG-3’ for Wdreb2β, and 5’-GGGAATTCGCGGACAAGATTGCGAAC-3’ and 5’-GCGTCGACCGGTAGGGGATCACAAC-3’ for Wdreb2γ. The PCR products were digested with EcoRI and SalI, and cloned into the EcoRI and SalI site of pBD-GAL4 Cam vector. These constructs (pBD-Wdreb2β and pBD-Wdreb2γ) contained chimeric genes encoding fusion proteins of the WDREB2 and GAL4 binding domain. The pBD-Wdreb2β and pBD-Wdreb2γ constructs were sequenced and introduced into a yeast strain YRG-2. The trans-activation ability was assessed on the SD medium without tryptophan and histidine and with 3mM 3-amino-1,2,4-triazole (3-AT).
The 5’ upstream regions of the Wrab17, Wrab18 and Wrab19 genes were amplified with the following promoter-specific primer sets containing the HindIII and XbaI linkers; 5’-CCAAGCTTGGTTTCTTCAGCTACTAGGC-3’ and 5’-CCTCTAGAGTGTTGGTACCTACTAATTTGTGTTC-3’ for Wrab17, and 5’-CCAAGCTTTTAATTTCATCTTCAATGGCTTTC-3’ and 5’-CCTCTAGACTCTTGTCTCTCACCAACAA-3’ for Wrab18 and Wrab19. The PCR-amplified promoter fragment was digested with HindIII and XbaI and replaced by the CaMV35S promoter of pBI121 (Clontech) to produce Wrab17::GUS, Wrab18::GUS and Wrab19::GUS constructs.
The Wdreb2γ cDNA sequence was amplified with the following primer set containing the BamHI linker; 5’-CGGGATCCGACAAGATTGCCTAACGCTAGA-3’ and 5’-CGGGATCCCCGACCAAACACCATAGACA-3’. The PCR fragment was digested with BamHI and inserted into the BamHI site between the CaMV35S promoter and the Nos terminator in pROK1 (Baulcombe et al., 1986). The chimeric Wdreb2 construct was named 35S::Wdreb2. All these constructs were purified by the Maxi-V500 Ultrapure Plasmid Extraction System (VIOGENE, CA, USA) and introduced into wheat callus line ‘HY-1’ by particle bombardment according to Takumi et al. (1999). During the particle bombardment, a chimeric construct of the luciferase gene, under control of the CaMV35S promoter, was included in all the assays. The GUS activity from various chimeric constructs was quantified according to Jefferson (1987) and normalized by the luciferase activity estimated by the microplate reader MTP-800AFC (Hitachi, Tokyo, Japan).
Our previous studies have shown that the winter cultivar M808 develops much higher levels of freezing tolerance than the spring cultivar CS after cold acclimation (Ohno et al., 2001; Kobayashi et al., 2004). We now compared the dehydration stress tolerance between these two cultivars. Seven-day-old seedlings grown under the standard conditions were pre-treated either with ABA or LT or non-pretreated before transferring onto dry filter paper and incubated at the standard temperature for 4 h. No significant difference of dehydration tolerance was observed between the two cultivars (Fig. 1A). Although not significant, pre-treatment with 20 μM ABA before dehydration slightly increased the survival rate compared with the non-treated control plants of both the cultivars. The LT treatment before dehydration however dramatically improved the level of dehydration tolerance in both the cultivars. Tolerance against salt stress (0.4 M NaCl for 1 week) was also compared between M808 and CS using 7-day-old seedlings. Although no significant cultivar difference was observed, LT treatment before salt stress significantly improved the level of dehydration tolerance in M808 (Fig. 1B). These results suggested that Cor/Lea genes induced during cold acclimation contributed to the increased tolerance to dehydration and salt stress.
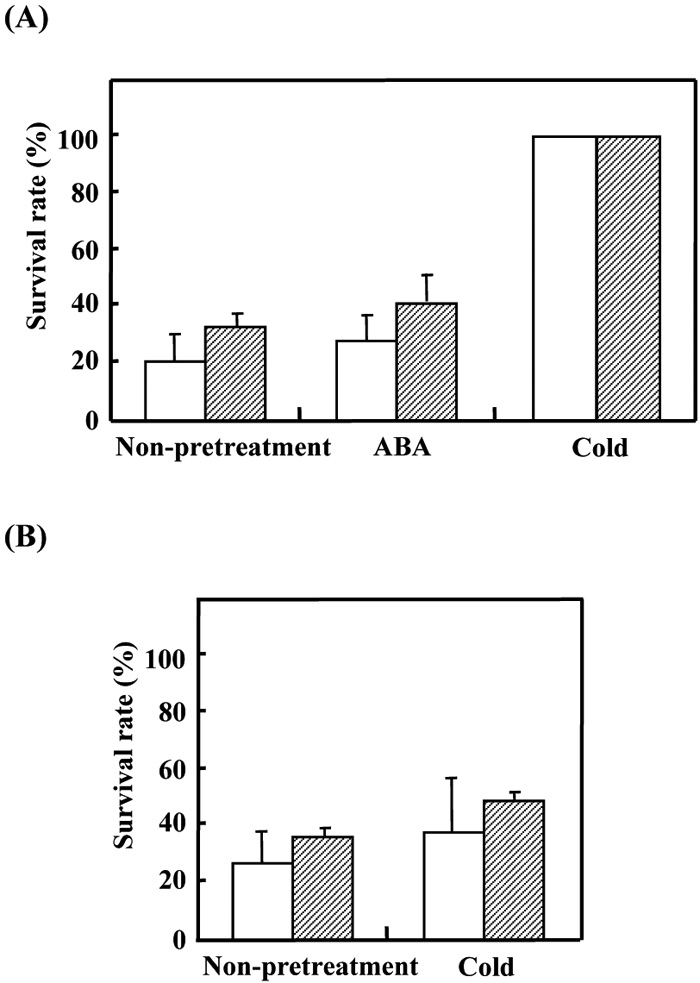 View Details | Fig. 1. Summary of the bioassay data for drought and salt tolerance in two wheat cultivars, CS and M808. Seedlings were grown for one week under the standard condition, and then pretreated either with ABA (20 μM) or 3-day LT (4°C) or non-pretreated before treatment of drought (A) and 0.4 M NaCl (B). Results are represented as means ± standard errors (3 independent experiments) in the two wheat cultivars (open bar: CS, shaded bar: M808). |
The TaDREB1 gene encodes a wheat homolog of Arabidopsis DREB2, and the TaDREB1 protein binds to the CRT/DRE motif (Shen et al., 2003). Different from the Arabidopsis DREB2, however, TaDREB1 is rapidly activated by all the treatments, namely; LT, salt and ABA. We identified another type of wheat DREB2 homolog in the EST library of developing caryopses and designated it as WDREB2 (Takumi et al., 2003). The EST clone contained polyA at the 3’ end and was a partial fragment lacking the 5’ end of the Wdreb2 cDNA. After the RACE-PCR for determining the 5’ end, an entire ORF of the Wdreb2 cDNA was amplified using the single strand cDNA from cold acclimated seedling leaves. Two cDNAs containing the entire ORF were isolated, both of which were identical to the Wdreb2 EST clone within the corresponding region (Fig. 2A). One clone was 1,035-bp in length and encoded a protein with 344 amino acid residues. Another clone had a 53-bp insertion, which resulted in a frame shift to create a new stop codon. The insertion-containing cDNA thus putatively encoded a truncated polypeptide with 66 amino acid residues.
 View Details | Fig. 2. Isolation and structural characterization of the Wdreb2 cDNA clones. (A) Comparison of the cDNA sequences identified in this study, the 5’ RACE product and the two isolated cDNA clones (cDNA 1 and cDNA 2). Arrow-heads represent the position of the primer set for amplification of cDNA clones containing an entire Wdreb2 ORF. Shaded bars and a dotted bar indicate the ERF/AP2 domain-encoding region and the 53-bp insertion sequence, respectively. (B) Phylogenetic tree based on amino acid sequences showing the relationship of WDREB2 with other DREB2 homologs. The tree was constructed by the UPGMA method based on the Nei’s genetic distances. Arabidopsis thaliana AtDREB2A and AtDREB2B (accession no AB007790); Triticum aestivum TaDREB1 (AF303376); Hordeum vulgare HvDRF1.3 (AY223807) and HvDRF2 (AF521302); Oryza sativa OsDREB1 (AF300970) and OsDREB2 (AF300971); Zea mays ZmDBF1 (AF493800) and ZmDBF2 (AF493799). (C) Alignment of amino acid sequences of the ERF/AP2 DNA-binding domain of WDREB2 with those of other DREB2 homologs. Identical amino acids are indicated by asterisks. |
The 344 amino acid polypepetide of WDREB2 contained a conserved ERF/AP2 domain and a nuclear localization signal. The amino acid sequence analysis showed high homology to other plant DREB2 homologs. A phylogenetic tree was constructed by UPGMA method (Fig. 2B). WDREB2 showed high homology to barley HvDRF1 and HvDRF2 (Xue and Loveridge, 2004). The multiple alignment of the ERF/AP2 domains of wheat and other plant DREB2 homologs is shown in Fig. 2C. The amino acid sequence of the WDREB2 ERF/AP2 domain showed identity of 93.3% with that of the HvDRF1.3 protein. Putative polypeptide encoded by the 53-bp insertion-containing cDNA of Wdreb2 lacked the ERF/AP2 domain.
Since the ERF/AP2 family function as a transcription factor in higher plants (Sakuma et al., 2002), a transient expression system was employed to confirm the nuclear localization of the WDREB2 protein. A chimeric CaMV35S::Wdreb2-GFP construct was introduced into the epidermal cells of onion via particle bombardment. The transient expression of the GFP fusion protein clearly demonstrated targeting of WDREB2 to the nucleus (Fig. 3).
 View Details | Fig. 3. Intracellular localization of the WDREB2 protein in the onion epidermal cells. Transient GFP expression was observed after bombardment of a chimeric construct of GFP fused with WDREB2 (A–C) or the positive control NLS (D–F). The GFP fluorescences are shown in panels (A) and (D). DAPI-staining nuclei in the corresponding epidermal cell layer are shown in panels (B) and (E), and the epidermal cell layers bombarded are in panels (C) and (F). |
To study copy number and genome distribution of the Wdreb2 gene in wheat genome, Southern blot analysis was conducted using DNA extracted from a single accession of the hexaploid and tetraploid wheat. Southern blot hybridization showed low copy numbers of Wdreb2 in hexaploid and tetraploid wheat genomes (Fig. 4A). Three and two major bands with several minor bands were observed in the BamHI-treated DNA of CS and Langdon, respectively. The minor bands indicated presence of low homologous copies of Wdeb2. To assign the Wdreb2 loci to homoeologous wheat chromosomes, aneuploid analysis was performed using a series of nulli-tetrasomic lines. Only in the nulli-tetrasomic lines of homoeologous group 1 chromosomes, Wdreb2 specific bands were absent (data not shown), indicating that the Wdreb2 loci represented the three homoeologous loci on chromosomes 1A, 1B and 1D in the common wheat genome. Southern blot analysis using ditelosomic lines of the group 1 chromosomes further showed that each band was respectively missing in the ditelo1AS, 1BS and 1DS lines, which lacked the long arm of each chromosome (Fig. 4B). These results indicated that the common wheat genome possesses three homoeologous loci of the Wdreb2 gene on the long arms of group 1 chromosomes.
 View Details | Fig. 4. Genomic distribution and chromosome assignment of the Wdreb2 homologous sequences in the wheat genome. (A) Southern blot analysis of the Wdreb2 gene. DNA from hexaploid (CS) and tetraploid (Langdon) wheat was digested with DraI, EcoRI, HindIII, BamHI and EcoRV and Southern blotted. The blot was probed with the 32P-labeled Wdreb2 cDNA. (B) Chromosome assignment of the Wdreb2 gene. DNA from CS ditelosomic series for the homoeologous group 1 chromosomes was digested with BamHI. Assigned chromosomes are indicated besides the Southern-blot. The ditelosomic lines are abbreviated, for example, dt1AS for the ditelosomic 1AS. |
To study the expression profiles of Wdreb2, LT-responsiveness was first examined by northern blot and semi-quantitative RT-PCR analyses. The level of the Wdreb2 homologous transcripts clearly increased during 3- and 5-day LT treatment under the standard light condition in both CS and M808 (Fig. 5A). After 5 days of continuous dark treatment, the amount of Wdreb2 transcripts showed marked decreases. An additional 3-day LT treatment in the dark after the 5-day LT treatment under the standard light condition did not affect the transcript level, suggesting that either the rate of transcription balanced the rate of transcript-turnover in the dark LT condition or that the transcript was stable at least for 3 days under this condition. These results suggested that the Wdreb2 transcription was enhanced by light and suppressed by darkness under the LT condition. This expression pattern is well conserved in a majority of Cor/Lea genes (Kobayashi et al., 2004). In Arabidopsis, cold-inducible expression of genes possessing the CRT/DRE is greatly enhanced by light (Kim et al., 2002). The expression of CBF3/DREB1a is also regulated by the circadian clock (Harmer et al., 2000). These facts suggest conservation of interaction between cold and light signals in higher plants.
 View Details | Fig. 5. Expression profiles of the Wdreb2 and Wrab17 genes during the LT treatment in the two wheat cultivars. (A) Light/dark modulation of the Wdreb2 expression in the seedling leaves of M808. Transcript accumulation was monitored by northern blot analysis probed with 32P-labeled Wdreb2 cDNA. The rRNA gene was used as a control. NA, non-acclimated control; L3 and L5, 3 and 5 days accumulation under standard light/dark regime; D3 and D5, 3 and 5 days accumulation under continuous dark; L5D3 and D5L3, L5 followed by D3 and D5 followed by L3, respectively. (B) Transcript accumulation of the Wdreb2 gene in response to LT was monitored by RT-PCR analysis using gene-specific primers. dA represents deacclimation under the standard conditions for 1 d after 3-d LT treatment. The ubiquitin gene was used as a control. (C) Northern blot analysis of the Wrab17 transcript using 32P-labeled probes. The rRNA gene was used as a control. |
Expression of some monocot CBF genes including barley HvCBF1 and HvCBF3 and rice OsDREB1A and OsDREB1B has been reported to be induced by low temperature within 2 h (Choi et al., 2002; Xue, 2003; Dubouzet et al., 2003). To examine whether such rapid induction of gene expression occurs in the Wdreb2 gene, RT-PCR analysis was conducted using the primer set amplifying the entire ORF of Wdreb2. The Wdreb2 transcript was detected at a low level under non-stress condition, and its level rapidly increased in M808 seedlings within 15 min of exposure to LT (Fig. 5B). Although there were some fluctuations, a time-dependent increase was delayed in CS compared with M808. To examine if Cor/Lea genes are transcriptionally regulated by the DREB2 homologs in wheat, we compared the Wrab17 expression profile with that of Wdreb2. It was found that the induction of Wdreb2 gene occurred prior to that of Wrab17 and that Wrab17 was induced earlier in M808 than in CS (Fig. 5C).
The transcript level of the Wdreb2 gene increased until day 10 in M808, while it decreased after the 5-day LT treatment in CS (Fig. 5B). Significant reduction was observed in the transcript level after 1-day de-acclimation of 3-day LT-treated seedlings of both cultivars, suggesting that Wdreb2 transcript was unstable under the normal temperature condition. No notable differences were observed in the Wdreb2 expression patterns between the two cultivars during the long-term LT treatment. The Wdreb2 transcript remained almost constant until day 63, with a slight increase noticed after day 35 in both the cultivars (Fig. 5B). The long-term profile of the Wrab17 expression agreed very well with that of Wdreb2 in both cultivars (Fig. 5C).
To study the effects of abiotic stresses other than low temperature on the Wdreb2 expression, wheat seedlings were either treated with 0.4 M NaCl (salt stress) or dehydrated under the standard temperature condition. Induction of the Wdreb2 gene under the drought condition was transient and after reaching maximum the amount of transcript decreased gradually (Fig. 6A). No significant difference was observed in the drought-induced expression patterns of Wdreb2 between the cultivars. The NaCl-treatment also transiently activated the Wdreb2 expression similar to the drought stress. These results showed that the Wdreb2 gene was responsive not only to LT but also to drought and salt.
 View Details | Fig. 6. Expression patterns of the Wdreb2 and Cor/Lea genes within the 24-h stress treatment in the two wheat cultivars. (A) Enhancement of the Wdreb2 gene expression by stress treatments of drought, NaCl (0.4 M) and ABA (20 μM). RT-PCR analysis was conducted to detect the Wdreb2 gene transcripts. The ubiquitin gene was used as a control. (B) Northern blot analysis of the Cor/Lea transcripts under drought-stress condition using 32P-labeled probes. The rRNA gene was used as a control. |
Expression of Wdhn13, one of wheat Cor/Lea genes, has been shown to be induced by LT, drought and salt stresses (Ohno et al., 2003). Putative CRT/DRE element and DRF1E motif were found in the 5’ upstream region of the Wdhn13 gene (our unpublished data). The DRF1E motif was also identified in the promoter regions of Wrab18 and Wrab19, which are HVA1 homologs of barley (Tsuda et al., 2000; Kobayashi et al., 2004; our unpublished data). HVA1 has been identified as a target gene of the HvDRF1 transcription factor (Xue and Loveridge, 2004). These reports suggest that the WDREB2 protein transcriptionally regulates the Cor/Lea genes. We therefore compared the expression profiles of four wheat Cor/Lea genes (Wdhn13, Wrab17, Wrab18 and Wrab19). As shown in Fig. 6B, all transcript levels of the Cor/Lea genes transiently increased within 1-day of the drought treatment, and the accumulation of Wrab17 transcript was more rapid than the others in both cultivars (Fig. 6B). The expression of Wdhn13, Wrab18 and Wrab19 followed that of Wdreb2. No clear difference was observed in the accumulation of Wrab18 and Wrab19 transcripts between the cultivars, and the expression patterns of Wrab18 and Wrab19 corresponded well to the expression profile of Wdreb2.
Arabidopsis DREB2 transcription factors are reported to be involved in the ABA-independent pathway, and expression of the DREB2 genes are not induced by the exogenous ABA treatment (Liu et al., 1998). We thus studied the Wdreb2 response to the exogenous ABA (20 μM). The level of Wdreb2 transcript increased within 15 min after the ABA-treatment and kept at a high level until 10 h (Fig. 6A). After 10 h, the amount of Wdreb2 transcript decreased. No significant difference between the cultivars was observed in the ABA response of Wdreb2 expression. The result suggests that the Wdreb2 gene is involved at least partly in an ABA-dependent pathway of stress signal cascades in wheat.
The Wdreb2 expression was activated by LT, drought, salt and exogenous ABA, and the accumulation patterns of the Wdreb2 transcript under LT condition were distinguished from those under the other stress conditions. The Wdreb2 expression was enhanced earlier in M808 than in CS within 24 h of exposure to LT, while the other stresses showed no significant difference of the Wdreb2-induction patterns between CS and M808 (Fig. 6). These results suggested that two independent pathways were related with the Wdreb2 activation, and that one pathway for drought and salt stresses was ABA-dependent and another for cold stress seemed to be ABA-independent. In Arabidopsis, the DREB2 transcription factors are activated only by drought and salt stresses through ABA-independent pathway (Liu et al., 1998). Cross talk between cold and drought stresses is realized at the cis-element level of Cor/Lea gene promoters in Arabidopsis (Shinozaki and Yamaguchi-Shinozaki, 2000). The DREB2 and CBF/DREB1 transcription factors recognize the identical CRT/DRE cis-element. However, some cereal transcription factors such as TaDREB1, HvDRF1 and WDREB2 were activated by both LT and drought (Shen et al., 2003; Xue and Loveridge, 2004; Fig. 5 and Fig. 6). Signal networks against cold and drought/salt stresses of cereals seem to be differed from those of Arabidopsis.
From the barley HvDRF1 locus, three different mRNA transcripts are generated due to alternatively splicing. Homologs of HvDRF1.1-HvDRF1.3 are present in wheat, and the DRF1 alternative spliced pattern is conserved between barley and wheat (Xue and Loveridge, 2004). Because our isolated cDNA clones of Wdreb2 showed high homology to the HvDRF1 genes (Fig. 2B), its alternative splicing pattern was studied. The 344 and 66 amino acids-encoding cDNAs of Wdreb2 were corresponding to the HvDRF1.3 and HvDRF1.2 transcripts, respectively.
To determine the splicing site, one genomic sequence of the Wdreb2 internal region was amplified using CS DNA and the PCR fragment was sequenced. Based on the genomic sequence, a primer set was designed for amplification of the alternatively spliced transcripts (Fig. 7A). The RT-PCR products from the cold-acclimated RNA of CS gave three different bands (Fig. 7B). Comparing nucleotide sequences of the genomic PCR fragment with the three different-length RT-PCR products, the exon/intron structure of Wdreb2 was revealed. The Wdreb2 gene consisted of 4 exons and 3 introns, and the 2nd and 3rd exons were 53- and 91-bp in length, respectively (Fig. 7A). The RT-PCR products were classified into three different types. The longest type consisted of all the four exons and was designated as Wdreb2α. The middle and shortest types lacked the 3rd exon and the 2nd and 3rd exons, respectively, and designated as Wdreb2β and Wdreb2γ. The Wdreb2α, Wdreb2β and Wdreb2γ forms corresponded to the HvDRF1.1, HvDRF1.2 and HvDRF1.3 transcripts, respectively. The Wdreb2β form putatively encoded the truncated polypeptide with 66 amino acids. These results indicated that pre-mRNA of the Wdreb2 gene was alternatively spliced into the three different forms similar to the barley HvDRF1 gene. One 6-bp deletion mutation and 5 single nucleotide polymorphisms (SNPs) were found among the 1st exon sequences of RT-PCR products. Based on the deletion and SNPs, the 1st exon sequences were grouped into three types (Fig. 7A), which likely represented the homoeologous sequences in the wheat genome. In other exons, no polymorphism was recognized among the RT-PCR products. The Wdreb2β and Wdreb2γ transcripts were both divided into three homoeologous groups. Only one RT-PCR product of the Wdreb2α transcript was sequenced. The alternative splicing of the Wdreb2 homologs is highly conserved in wheat and barley as previously mentioned by Xue and Loveridge (2004). Moreover, the SNPs in each spliced form indicate that the alternative splicing equally occurs in all Wdreb2 transcripts derived from the three homoeologous loci assigned to 1A, 1B and 1D chromosomes.
 View Details | Fig. 7. Structural characterization and stress-response-mediated alternative splicing of the Wdreb2 transcripts. (A) Exon/intron structure of Wdreb2 and the three spliced forms. Arrow-heads indicate position of the primer sets used in this study. The Wdreb2 gene contained four exons (E1 to E4) and three introns (I1 to I3). The three spliced transcripts were designated as Wdreb2α, Wdreb2β and Wdreb2γ. Only in the 1st exon sequence, some SNPs and a 6-bp insertion/deletion were found among the cloned RT-PCR products. The position of the polymorphisms and the corresponding spliced forms are represented below (see Results). (B) Effect of drought, NaCl (0.4 M), cold (4°C) and ABA (20 μM) on accumulation of the three spliced transcripts in the two wheat cultivars. Total RNA was isolated from the seedling leaves after 2 or 6 h stress treatment. One primer set amplifying the entire ORF of Wdreb2 transcripts gave a single band on 0.8% agarose gel. Another primer set amplified three different bands indicating the Wdreb2α, Wdreb2β and Wdreb2γ forms, and the RT-PCR products after 31 and 35 cycles were separated on 1.5% agarose gel. The ubiquitin gene was used as a control. |
Stress responses of Wdreb2 were compared among the three alternative spliced forms. RT-PCR products generated by the primer set amplifying the entire ORF were recognized as a single band on the 0.8% agarose gel, but the primer set amplifying the internal region generated three spliced forms on the 1.5% gel (Fig. 7B). Wdreb2β showed the most prominent expression level among the three forms in all stress-treated and non-treated samples both from CS and M808. The Wdreb2α and Wdreb2γ transcripts showed varying accumulation levels among the samples. To study more in detail the expression levels of the different spliced forms, we designed three primer sets specific to the three forms. The primer sets generated single spliced product-specific bands. Accumulation patterns of the three spliced transcripts were studied in response to LT, drought, salt and ABA. The Wdreb2β transcript was present under the non-stress condition. The Wdreb2α and Wdreb2γ transcripts transiently appeared in response to LT treatment, and the maximum levels were reached earlier in M808 than in CS (Fig. 8A). The Wdreb2β level reached a plateau during 2–8 h LT treatment in M808, and the time-dependent increase was delayed in CS compared with M808. Drought, NaCl (0.4 M) and ABA (20 μM) also transiently induced the Wdreb2α and Wdreb2γ transcripts in both CS and M808 (Fig. 8B–D). These results indicate that the ratio of the Wdreb2α and Wdreb2γ forms to the Wdreb2β form changes in response to drought and salt stresses. No cultivar difference of the ratio was clearly observed, and the accumulation patterns of the three spliced forms under the drought and salt stress conditions were well corresponding to those after the ABA treatment. These results suggest that the quantitative change of the Wdreb2α and Wdreb2γ forms should be regulated by splicing-related factor(s) through ABA-dependent signal cascade. The Wdreb2β form kept continuous levels under the drought-, salt- and ABA-treated conditions.
 View Details | Fig. 8. Expression profiles of the three spliced forms of the Wdreb2 transcripts within 24-h stress conditions in the two wheat cultivars. RT-PCR analysis was conducted using the specific primer set to the three forms. Accumulation patterns of the three transcript forms under cold-, drought-, NaCl- and ABA-treated conditions are shown in (A), (B), (C) and (D), respectively. The ubiquitin gene was used as a control. |
Because the accumulation patterns of the three spliced forms under LT condition were clearly differed from those under the other stress conditions, the alternative splicing of the Wdreb2 transcript was differentially regulated through two independent pathways. One pathway for drought and salt stresses should be ABA-dependent, and another for cold stress ABA-independent. These results strongly suggest that significant change of splicing-related factors occurs under the abiotic stress conditions and the modified factors affect the alternative splicing patterns of the Wdreb2 transcripts. This interesting phenomenon should be studied in more detail, in future. Thus, amount of the WDREB2 transcription factors was differentially controlled at the transcript accumulation level and alternative splicing under cold and drought/salt stress conditions in wheat cells.
We studied the trans-activation activity of the WDREB2 protein toward the downstream target genes. First, the trans-activation ability of the WDREB2β and WDREB2γ was assessed using the yeast system. The yeast transformants carrying the pBD-Wdreb2γ construct survived on the His– medium (Fig. 9), indicating that WDREB2γ acted as a transcriptional activator. The result also showed that the pBD-Wdreb2β lacked not only ERF/AP2 domain but also transcriptional activation domain.
 View Details | Fig. 9. Trans-activation activity of the WDREB2β and WDREB2γ protein in yeast cells. Growth of yeast transformants and the YRG-2 wild type was monitored on the SD and SD-TRP-HIS+3AT media. 3AT was used as a competitive inhibitor of the HIS3 gene product. The position of three transformed constructs on the medium is indicated on the right side. |
Next, the trans-activation ability was assessed using three Cor/Lea genes as targets in the wheat cell line. It has been reported that the HvDRF1 exhibit very little activity towards the DRE/CRT motif recognized by Arabidopsis DREB2 and maize DBF1 (Xue and Loveridge, 2004). In the 5’ upstream sequences of Wrab18 and Wrab19, the DRE/CRT and DRF1E motifs were identified (our unpublished result). The Wrab17 promoter sequence contained the DRE/CRT motif, but no typical DRF1E motif-containing sequence was found (our unpublished result). The 5’ upstream regions of the three genes were combined with the GUS gene and the Nos terminator. The three chimeric constructs, designated as Wrab17::GUS, Wrab18::GUS and Wrab19::GUS, were introduced into the cultured line HY-1 via particle bombardment. A chimeric construct of the Wdreb2γ cDNA under control of the CaMV35S promoter (35S::Wdreb2) was co-bombarded with the Wrab::GUS constructs. After bombardment, the GUS activities were normalized by the luciferase activities. In the Wrab17::GUS and Wrab18::GUS constructs, bombardment with the 35S::Wdreb2 construct appeared to result in slightly higher GUS activities than that without the 35S::Wdreb2, but the differences of GUS activity were not significant (Fig. 10). In the Wrab19::GUS construct, the introduction together with 35S::Wdreb2 significantly increased the GUS activities, although the CaMV35S promoter is relatively weak in wheat cultured cells compared with rice Actin 1 and maize Ubiquitin 1 promoters (Takumi et al., 1994). These in vivo monitoring of the interaction between WDREB2 and Wrab promoter sequences indicated that the WDREB2 protein functioned as a transcriptional activator similar to the HvDRF1 protein and strongly suggested that at least the Wrab19 gene is a downstream target of the Wdreb2 gene in wheat cells. These results strongly indicate that the at least some of wheat Cor/Lea genes are regulated by the Wdreb2 genes under abiotic stress conditions. Further studies are required to prove the direct physical interaction of the CRT/DRE and/or DRF1E elements in these Cor/Lea promoter regions with the WDREB2 protein.
 View Details | Fig. 10. Trans-activation of Cor/Lea genes by WDREB2 in wheat cultured cell line ‘HY-1’. GUS activities were normalized as the luciferase activities, expressed under the control of the CaMV35S promoter. The meansÅ}standard errors were calculated from data in 3–5 experiments. The GUS gene under the control of a CaMV35S promoter and maize Adh1 1st intron (pCaMVIGN) was used as a control. |
The Wdreb2 gene seemed to be a wheat ortholog of the barley HvDRF1 gene based on their high sequence homology and identical feature of alternative splicing. Wrab18 and Wrab19 showed high homology to barley HVA1 (Tsuda et al., 2000; Kobayashi et al., 2004). Together with the presence of the DRF1E motif in the promoter region of Wrab18 and Wrab19, it is indicated that they are wheat orthologs of HVA1. Thus, these stress signal pathways from Wdreb2 to Wrab18/Wrab19 and from HvDRF1 to HVA1 are functionally conserved in wheat and barley.
We are grateful to Dr. T. Shimada for providing us with the HY-1 callus line and Dr. Y. Niwa for the sGFP(S65T) expression vector and nuclear-targeting GFP construct (NLS). We also thank Dr. N. Shiraishi for his kind technical advices. The work was supported in part by Grant-in-Aids from the Ministry of Education, Culture, Sports, Science and Technology of Japan to CN (no. 13306002) and ST (no. 17780005). The sequence of Wdreb2 has been deposited in the DDBJ database (accession number AB193608). Contribution no. 171 from the Laboratory of Plant Genetics, Faculty of Agriculture, Kobe University.
|