| Edited by Hirokazu Inoue. Akihiko Ichiishi: Corresponding author. E-mail: akihiko@itakura.toyo.ac.jp |
DNA is continuously threatened by endogenous and exogenous genotoxic agents. In order to maintain genomic integrity, every organism from bacteria to human beings has developed various DNA repair mechanisms during evolution (Friedberg et al., 2006). Nucleotide excision repair (NER) is the main mechanism responsible for removing a wide range of bulky DNA lesions, including UV-induced cyclobutane pyrimidine dimers (CPDs) and 6-4 photoproducts (6-4PPs). NER consists of multistep reactions and requires the coordinated action of approximately 30 different proteins implicated in damage recognition, helix opening, lesion verification, dual incision of the damaged strand, removal of lesion-containing oligonucleotides, gap-filling DNA synthesis, and ligation. NER is functionally divided into two mechanism designated transcription-coupled repair (TCR) and global genomic repair (GGR). TCR responds to damage of the transcribed strand of transcriptionally active genes while GGR deals with general genome damage. These two NERs utilize different factors during the damage recognition step, although thereafter they both recruit the same components to remove the lesion.
In the yeast Saccharomyces cerevisiae, a combination of Rad14, Rad4-Rad23, RPA, TFIIH, Rad1-Rad10, and Rad2 mediates the dual incision of the damaged DNA strand, releasing lesion-containing DNA fragments (Guzder et al., 1995). Rad14, Rad4-Rad23, and RPA function at the damage recognition step. The Rad3 and Rad25 DNA helicases, which are components of TFIIH, unwind the duplex DNA around the lesion, and the Rad1-Rad10 and Rad2 nucleases incise the damaged DNA strand on the 5 and 3 side of the lesion, respectively (Prakash and Prakash, 2000).
In the filamentous fungus Neurospora crassa, only a single gene involved in NER has previously been identified. This gene, mus-38, was characterized as a homolog of S. cerevisiae RAD1 (Hatakeyama et al., 1998; Ishii et al., 1998; Inoue, 1999). Interestingly, N. crassa has another excision repair mechanism for the removal of UV-induced damage that is mediated by the MUS-18 protein (Ishii et al., 1991; Yajima et al., 1995). This protein is a homolog of Schizosaccharomyces pombe UVDE (Yonemasu et al., 1997). The mus-18 homolog gene does not exist in S. cerevisiae. Recently, the N. crassa genomic sequence database has become available, thereby facilitating the identification of structural homologs of NER genes in this organism (Galagan et al., 2003; Borkovich et al., 2004).
In this study, we describe the identification and characterization of mus-43 (homolog of S. cerevisiae RAD14) and mus-44 (homolog of S. cerevisiae RAD10) in N. crassa. The mutants of both these genes are sensitive to UV irradiation and 4-nitroquinoline 1-oxide (4NQO), and are epistatic to mus-38. Immunological assays demonstrated that the mus-18 excision repair pathway removed UV-induced damage more effectively than mus-43 and mus-44. These results indicate that in N. crassa mus-43 and mus-44 function in NER but independently of the mus-18 excision repair pathway.
The N. crassa strains used in this study are listed in Table 1. C1-T10-37A and C1-T10-19a are wild-type strains that are closely related to the standard Oak Ridge wild-type strains (Tamaru and Inoue, 1989). pT7Blue AccepTor Vector (Novagen) was used as a vector for T-cloning of PCR products. Plasmid pCB1003 (Carroll et al., 1994), carrying the Escherichia coli hygromycin B resistance gene driven by the Aspergillus nidulans trpC promoter, was used as a vector for transformation of N. crassa and was obtained from the Fungal Genetics Stock Center (University of Missouri, Kansas City, MO). The mus-43 and mus-44 mutants were generated by repeat-induced point mutation (RIP), in which duplicated sequences are disrupted by GC to AT transitions during the dikaryotic stage preceding meiosis (Selker et al., 1987). For the induction of RIP, the genes of interest were first duplicated in N. crassa by introducing an additional copy into a wild-type strain. The resulting strains were then crossed with another wild-type strain, and individual sexual spores were picked and germinated. In order to confirm the isolation of RIPed mutants, we examined mutagen sensitivity and performed DNA sequencing. For the induction of RIP, DNA fragments of mus-43 and mus-44 were generated by PCR using N. crassa genomic DNA as a template and the primer pairs ncrad14-F (5’-CCT GCT ATC CCG CCC AAC T-3’) with ncrad14-R (5’-CAC ATG ACG GCT GCG GAG T-3’) and ncrad10-F (5’-CGA GAC ACG TTG AAT CGA GAC T-3’) with ncrad10-R (5’-CAT CAG GCA ATT CCA GAC TTC C-3’), respectively. The resulting PCR products were cloned into pT7Blue AccepTor Vector in order to generate pMI14 and pYK10, respectively. The KpnI and XbaI fragment from pYK10 was ligated into pCB1003 to generate pYK10-T and the EcoRI fragment from pCB1003 including the hygR gene was ligated into the EcoRI site of pMI14 to generate pMI14-T.
 View Details | Table 1 Strains of N. crassa used in this study |
Genetic manipulation of N. crassa was performed as described by Davis and de Serres (1970). The transformation of N. crassa was performed as described by Vollmer and Yanofsky (1986) and Tomita et al. (1993), except that Trichoderma harzianum lysing enzymes (Sigma) and 2 mg/ml of 1 M sorbitol were used for the production of spheroplasts.
For the reversion assay, two pantothenic acid-requiring strains, pan-2 (B36) and pan-2 (OGW1), were crossed with mus-43 and mus-44 to yield pan-2 (B36) mus-43 and pan-2 (OGW1) mus-43, and pan-2 (B36) mus-44 and pan-2 (OGW1) mus-44 strains, respectively. The pan-2 (B36) and pan-2 (OGW1) alleles carry base substitution and frameshift mutation, respectively (Suzuki et al., 2005; Kawabata et al., 2007). Conidial suspensions (1 × 107 conidia/ml) were irradiated with various doses of UV and cell survival and reversion were measured as follows: (i) A portion of the conidial suspension was diluted to 104 conidia/ml and 0.1 ml aliquots of the diluted suspension were mixed with medium supplemented with panthotenate (10 μg/ml) and plated on Petri dishes. Each plate was incubated at 30°C for 3 days and the number of colonies was counted. (ii) In order to determine the number of pan-2 revertants, 1 ml aliquots of conidial suspension were mixed with a medium supplemented with panthotenate (0.04 μg/ml) and plated on Petri dishes. The number of revertants was counted and the reversion frequency was calculated as the number of revertants per 107 survivors.
Qualitative assay of the sensitivity to chemical mutagens and other chemicals was performed using spot tests as described by Watanabe et al. (1997). Methyl methanesulfonate (MMS), camptothecin (CPT), N-methyl-N’-nitro-N-nitrosoguanidine (MNNG), 4-nitroquinoline 1-oxide (4NQO), hydroxyurea (HU), or bleomycin (BLM) were added to agar medium at final concentrations of 0.015%, 0.3 μg/ml, 0.5 μg/ml, 0.075 μg/ml, 1.5 mg/ml, and 4 μg/ml, respectively. Conidial suspensions were spotted onto these plates and grown at 30°C for 2 days. In order to test UV sensitivity, conidial suspensions spotted onto a plate were irradiated at a dose of 300 J/m2.
Quantitative sensitivity to UV was determined as described by Inoue and Ishii (1984). Conidial suspensions (106 conidia/ml) were irradiated at various doses of UV and 0.1 ml aliquots were mixed with medium. Each plate was incubated for 2 day at 30°C and the numbers of colonies were counted.
Photoreactivation was determined as described by Sakai et al. (2003). Conidial suspensions (1 × 107 conidia/ml) were maintained under two white-light lamps (FL15W; Toshiba) for 10, 20, and 30 min at room temperature after UV irradiation. The conidial suspensions were then diluted to 104 conidia/ml. Aliquots (0.1 ml) of the diluted suspension were mixed with medium and plated on Petri dishes. Each plate was incubated for 2 days at 30°C and the numbers of colonies were counted.
Six milliliters of conidial suspension (2 × 107 conidia/ml in 67 mM phosphate buffer, pH 7.0) was cooled and irradiated by UV (100 J/m2) in a 6-cm diameter Petri dish. After irradiation, the conidia were immediately cooled in an ice bath, homogenized, and the DNA extracted. In order to determine the amount of UV photoproducts, the DNA was incubated with KTM53 or KTM50 antibodies (Kyowa Medex) that specifically recognize CPDs or 6-4PPs, respectively. Antibody binding was assayed by ELISA as described by Matsunaga et al. (1990) with the following modifications. The concentration of DNA used for the ELISA was 50 ng per well for KTM53 and 300 ng per well for KTM50. Incubation with the primary antibody was for 30 min at 37°C. The secondary antibody, a peroxidase-conjugated goat IgG fraction to mouse IgG (Cappel), was diluted 1/4000 for KTM53 and 1/2000 for KTM50 and incubation was for 40 min at 37°C.
The putative homologs of S. cerevisiae RAD10 and RAD14 were identified by BLAST searches (protein to protein) of the N. crassa genome sequence database at the Whitehead Institute Center for Genome Research (http://www.broad.mit.edu/annotation/fungi/neurospora/). The N. crassa gene mus-43 (NCU08742.3) was identified as an S. cerevisiae RAD14 homolog, which encodes a 435-amino acid protein containing zinc-finger motifs that are characteristic of DNA binding domains which conserved in S. cerevisiae Rad14 and human XPA. The gene exhibited 66% similarity to S. cerevisiae Rad14 and 59% similarity to human XPA, and exhibited 48% and 35% identity to S. cerevisiae Rad14 and human XPA, respectively. We also identified the N. crassa gene mus-44 (NCU07066.3) as a candidate S. cerevisiae RAD10 homolog. This gene encodes a 405-amino acid protein containing a characteristic UvrC-like domain which conserved in S. cerevisiae Rad10 and S. pombe Swi10. It exhibited 66% and 59% similarity to S. pombe Swi10 and S. cerevisiae Rad10, and 48% and 35% identity to S. pombe Swi10 and S. cerevisiae Rad10, respectively.
In order to characterize the mus-43 and mus-44 genes, we generated mus-43 and mus-44 mutants by RIP, in which duplicated sequences are disrupted by GC to AT transitions during the dikaryotic stage preceding meiosis (Selker et al., 1987). Neither mutant exhibited any defects during vegetative growth, conidiation, or sporulation. In order to determine the function of these genes in DNA repair, the mutagen sensitivity of the mus-43 and mus-44 mutants was examined using a spot test. The mus-43 and mus-44 mutants exhibited higher sensitivity to UV and 4NQO than the wild-type, but not to MNNG, MMS, CPT, HU, or BLM (Fig. 1A). Furthermore, in order to measure the quantitative UV-sensitivity of the mus-43 and mus-44 mutants, we carried out a UV dose-dependent killing assay. The mus-44 mutant exhibited higher sensitivity to UV than the wild-type and the same sensitivity as mus-38 (Fig. 1B). The mus-38 is a homolog of S. cerevisiae RAD1 and human XPF (Hatakeyama et al., 1998; Ishii et al., 1998). The mus-43 mutant also exhibited a significant sensitivity to UV irradiation, although its sensitivity was less than that of either the mus-44 or the mus-38 mutant.
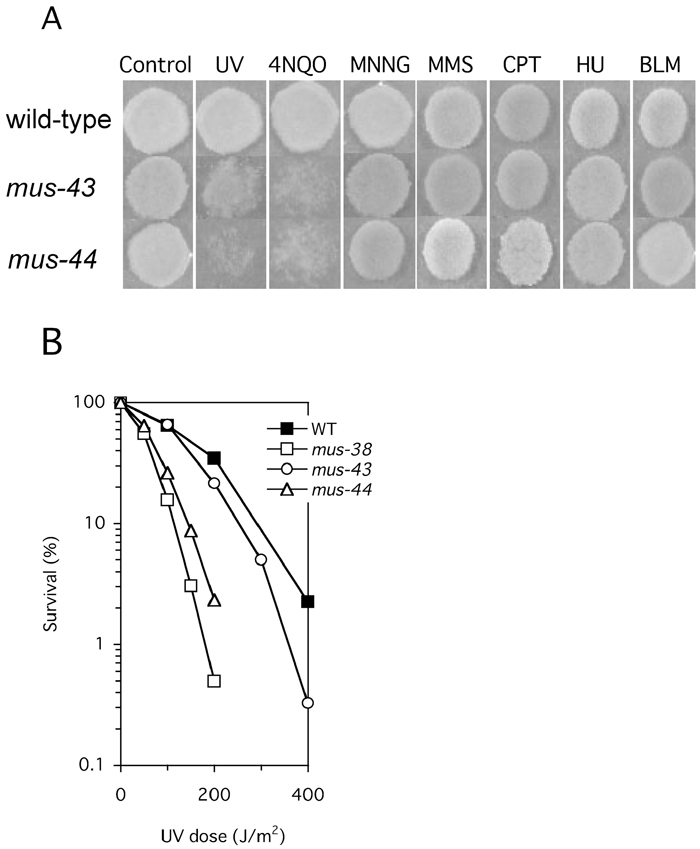 View Details | Fig. 1 Characterization of mutants. (A) Qualitative analysis of sensitivities to chemical agents and UV irradiation. Conidial suspensions were spotted onto agar plates containing the indicated chemicals (see “Materials and methods”). In the case of UV, conidia were irradiated after spotting. (B) Quantitative cell killing assay testing the sensitivity of mus-43 and mus-44 to UV. Conidial suspensions were UV-irradiated, mixed with the plating medium, and incubated at 30°C. Filled squares, wild-type; open circles, mus-43; open triangles, mus-44; open squares, mus-38. Each point represents the mean of at least three independent experiments. |
The mus-38 mutant exhibited approximately 6 time higher UV-induced mutation frequency at the ad-3 locus than the wild-type (Ishii et al., 1998). In this study, we used two pan-2 mutants, B36 and OGW1, as testers for the UV-induced reversion assay. These mutants have base substitution and frameshift mutations at the pan-2 locus. The UV-induced reversion frequencies of the mus-43 and mus-44 strains were 5- to 10-fold higher than that of the wild-type (Fig. 2).
 View Details | Fig. 2 Frequency of UV-induced reversion at the pan-2 locus. The reversion frequency is expressed as panthotenate prototrophs/107 survivals: (A) B36 (base substitution); (B) OGW1 (frameshift). |
It is known that the mus-38 mutant shows a partial photoreactivation defect (PPD) phenotype (Ishii et al., 1998). In the wild-type, the viability of UV-irradiated conidia exhibited a rapid recovery within the first 10 min of photoreactivation; thereafter, the incremental rate of recovery gradually fell, and finally saturated after 30 min (Fig. 3A). Although the cells of the mus-44 mutant (Fig. 3D) exhibited some photoreactivation recovery, the incremental rate was very low and it saturated at a low level of viability similar to the mus-38 mutant (Fig. 3B). However, the mus-43 mutant exhibited almost the same photoreactivation rate as the wild-type, but required a longer time (60 min) than the wild-type (30 min) to reach saturation (Fig. 3C).
 View Details | Fig. 3 Photoreactivation of wild-type, mus-38, mus-43, and mus-44 mutants. Conidial suspensions were UV-irradiated with the indicated doses of UV and immediately exposed to visible light for the indicated times to induce photoreactivation. (A) Wild-type, (B) mus-38, (C) mus-43, (D) mus-44. Each point represents the mean of at least three independent experiments. |
Genetic interactions between mus-43, mus-44 and various DNA repair genes were examined by measuring the UV sensitivity of single or double mutants carrying mus-43 or mus-44 and mus-38, mus-18, mei-3, or uvs-2 genes (Table 1). The mei-3 encodes the S. cerevisiae Rad51 homologous protein and functions in homologous recombination repair (Hatakeyama et al., 1995), and uvs-2 encodes the Rad18 homologous protein that functions in post-replication repair (Tomita et al., 1993). Double mutants of mus-43 mus-18, mus-43 mei-3, and mus-43 uvs-2 exhibited higher sensitivity to UV than did the parental strains (Fig. 4B–D). However, the sensitivity of the mus-43 mus-38 double mutant was almost the same as that of the mus-43 mutant (Fig. 4A). The sensitivity of the double mutants in the mus-44 background exhibited the almost same results as for the mus-43 background double mutants (Fig. 5). These results indicated that mus-43 and mus-44 are epistatic to mus-38. Therefore, mus-43 and mus-44 may function in NER, but not in the mus-18 excision repair pathway, homologous recombination repair, or post-replication repair.
 View Details | Fig. 4 The epistatic relationship of mus-43. (A) Open circles, mus-43; open triangles, mus-38; filled squares, mus-43 mus-38. (B) Open circles, mus-43; open triangles, mus-18; filled squares, mus-43 mus-18. (C) Open circles, mus-43; open triangles, uvs-2; filled squares, mus-43 uvs-2. (D) Open circles, mus-43; open triangles, mei-3; filled squares, mus-43 mei-3. Each point represents the mean of at least three independent experiments. |
 View Details | Fig. 5 The epistatic relationship of mus-44. (A) Open circles, mus-44; open triangles, mus-38; filled squares, mus-44 mus-38. (B) Open circles, mus-44; open triangles, mus-18; filled squares, mus-44 mus-18. (C) Open circles, mus-44; open triangles, uvs-2; filled squares, mus-44 uvs-2. (D) Open circles, mus-44; open triangles, mei-3; filled squares, mus-44 mei-3. Each point represents the mean of at least three independent experiments. |
Each of the mus-18, mus-43, and mus-44 mutants are moderately sensitive to UV; however, the mus-18 mus-43 and mus-18 mus-44 double mutants exhibited extremely high sensitivity to UV (Fig. 4B and Fig. 5B). The roles of these genes in the excision of UV damage were further characterized by direct assay of the two major UV-induced lesions, CPDs and 6-4PPs. This was achieved by the use of antibodies that specifically bind to these types of damage. In wild-type cells, binding sites for the antibodies to both CPDs and 6-4PPs decreased rapidly with incubation time after UV irradiation (Fig. 6). The mus-18 mutant was slow in removing both types of damage, with approximately 70% of the CPDs and 35% of the 6-4PPs remaining after 3 h. In contrast, the rate of repair of UV-damage in mus-43 and mus-44 mutants were only slightly lower than that in wild-type. Furthermore, there was no decrease in antibody binding sites in the mus-18 mus-43 and mus-18 mus-44 double mutants over the 3 h of the assay, indicating a failure to excise both types of UV damage in these strains. These results suggest that N. crassa has two redundant repair mechanisms—the NER and mus-18 excision repair pathways—for UV-induced CPDs and 6-4PPs, and that the mus-18 excision repair pathway is predominant in the removal of these lesions.
 View Details | Fig. 6 Repair of cyclobutane pyrimidine dimers (CPDs) and 6-4 photoproducts (6-4PPs). UV-photoproducts remaining in the UV-irradiated DNA were determined by using antibodies that specifically bind to CPDs (A) or 6-4PPs (B). Filled squares, wild-type; open circles, mus-43; open triangles, mus-44; open diamonds, mus-18; filled circles, mus-43 mus-18; filled triangles, mus-44 mus-18. Error bars indicate the standard deviations of three independent experiments. |
In this study, we report the identification of N. crassa homologs of the S. cerevisiae NER genes RAD10 and RAD14 from the N. crassa genomic sequence database. The mutagen sensitivity of the mus-43 and mus-44 mutants was similar to that of the previously identified mus-38 mutant, which is an N. crassa RAD1 homolog (Fig. 1). The mus-43 mus-38 and mus-44 mus-38 double mutants did not elevate the UV sensitivity of either parent strain; however, the mus-43 mus-18 and mus-44 mus-18 double mutants synergistically elevated UV sensitivity (Fig. 4A, 4B, Fig. 5A, and 5B). These results suggest that both the mus-43 and mus-44 genes are involved in NER, but not in the mus-18 UV-damage specific excision repair pathway.
Unlike S. cerevisiae and other eukaryotes, N. crassa and S. pombe have a second excision repair pathway for UV damage, mus-18 and UVER pathway, respectively. Therefore, both organisms exhibit hyper-resistance to UV irradiation. Immunological assay demonstrated that the removal of UV-induced lesions in mus-43 and mus-44 single mutants (in which only mus-18 excision pathway is operative) was almost normal (Fig. 6). In mus-18 mutant (in which NER is operative) the rate of repair of both types of damage was considerably slower. These results indicate that the mus-18 excision repair pathway more effectively removes UV damage than the NER pathway, and that in N. crassa most photolesions are removed by the mus-18 excision repair pathway. However, the mus-43 mus-18 and mus-44 mus-18 double mutants completely abolished the ability to remove UV damage. In fact, the mus-43 mus-18 and mus-44 mus-18 double mutants are synergistically more sensitivity to UV than either single mutant (Fig. 4B and Fig. 5B). The mus-18 mutant (in which NER pathway is operative) and mus-43 or mus-44 mutant (in which the mus-18 excision repair pathway is operative) could remove approximately 30% and 75% of CPD after 3 h incubation, respectively (Fig. 6A). In contrast, NER pathway and mus-18 excision repair pathway could remove approximately 65% and 85% of 6-4PPs after 3 h, respectively (Fig. 6B). There results indicated that N. crassa possesses the excess activity for the removal of 6-4PPs. The ability to remove the 6-4PPs may be more important for tolerance to UV in N. crassa, because it is known that 6-4PPs are more toxic than CPDs in NER-deficient human cells (Nakajima et al., 2004). Moreover N. crassa has a CPD specific photolyase which functions under the light condition, suggesting that N. crassa also possesses the excess activity for the repair of CPDs (Shimura et al., 1999). In addition to repair of UV damage, the NER pathway appears to be necessary for the repair of the chemical adducts since the mus-43 and mus-44 mutants are both sensitive to 4NQO (Fig. 1A), whereas the mus-18 mutant does not exhibit sensitivity (Ishii et al., 1991).
The mus-43 mutant exhibits low UV sensitivity compared to both the mus-38 and mus-44 mutants (Fig. 1B). The UV sensitivities of the S. cerevisiae RAD14 mutant and the S. pombe rhp14 mutant are similar to those of the RAD1 and swi10 mutants, respectively (Bankmann et al., 1992; Hohl et al., 2001). The mus-43 may be involved exclusively in the NER pathway; consequently, the mus-43 mutant exhibits low sensitivity to UV irradiation since a majority of the UV damage is removed by the mus-18 pathway. The observation that the mus-44 and mus-38 mutants exhibit higher sensitivity to UV than the mus-43 mutant suggest that mus-44 and mus-38 may participate in other DNA repair pathways in addition to NER. This is similar to S. cerevisiae Rad1-Rad10, which also functions as a nuclease in recombination repair (Davies et al., 1995) and base excision repair (Boiteux and Guillet, 2004).
We thank Hirokazu Inoue for providing the Neurospora strains. This work was supported by a research grant from The Specific Research Foundation for promoting science at Toyo University.
|