| Edited by Fumio Tajima. Alfred E. Szmidt: Corresponding author. E-mail: aszmiscb@mbox.nc.kyushu-u.ac.jp |
Larch species (Larix sp. Mill) are prominent components of the boreal forest. They are widely distributed across Eurasia and constitute 40% of its forest (Farjon, 1990). Relationships among Eurasian Larix species and their classification are still controversial. For instance, in western Urals and western Siberia some authors recognize only one species: L. sibirica (Bobrov, 1972; Bobrov, 1978; Farjon, 1990; Milyutin and Vishnevetskaia, 1995). On the other hand, two species (L. sibirica and L. sukaczewii) have been recognized in this region by some other authors (Dylis, 1947; Abaimov et al., 1998, 2002; Bashalkhanov et al., 2003). Similarly, there are different classifications of Larix species occurring in central and eastern Siberia. For instance, Milyutin and Vishnevetskaia (1995) recognized there only one species: L. gmelinii. On the other hand, L. gmelinii and two varieties (L. gmelinii var. olgensis and L. gmelinii var. japonica) have been recognized there by Farjon (1990). Yet another classification of the Larix species occurring in central and eastern Siberia was proposed by Abaimov et al. (2002) who recognized there: L. cajanderi, L. gmelinii and its three varieties: L. gmelinii var. olgensis, L. gmelinii var. japonica and L. gmelinii var. kamchatica. Several other classifications were also proposed for the Larix species from this region (Bobrov, 1978).
The advance and retreat of glaciers have significantly influenced the distribution and diversity of plant species. Climatic changes associated with glaciations led to large scale migration and reduction in population size and number followed by colonization and population expansion as the glaciers retreated (Pielou, 1991). During the last glacial maximum (LGM) approximately 18,000 ~ 20,000 years ago, a great part of the current taiga zone of the northern Eurasia, where the extant Larix species occur, was covered by ice (Svendsen et al., 1999; Tarasov et al., 2000), while most of Beringia (eastern Eurasia) remained ice-free during LGM (Hamilton and Thorson, 1983; Porter et al., 1983; Ananyeyev et al., 1993; Bennett, 1997). Both pollen and macrofossil evidence clearly indicate that Larix species survived during LGM in multiple and often isolated refugia located south of the Urals, southern Siberia and in western Beringia (Kremenetski, 1994; Tarasov et al., 2000; Andreev et al., 2002). Additional refugia were present in northern Kazakhstan, near the Sea of Azov and in the Yana-Indigirka lowland in the Russian Far East (Tarasov et al., 2000; Anderson et al., 2002). These refugia were suggested to serve as a source of Larix re-colonization of various regions of Eurasia during late Pleistocene (Semerikov et al., 1999).
Previous studies suggested that Eurasian Larix species are weakly diverged and have low levels of population differentiation (Lewandowski et al., 1991; Timerjanov, 1997; Semerikov et al., 1999, 2003; Semerikov and Lascoux, 2003; Wei and Wang, 2003, 2004a,b; Larionova et al., 2004). These findings are surprising taking into account complex and heterogeneous climatic history of Eurasia and certain reproductive features of Larix species such as limited seed and pollen dispersal (Duncan, 1954; Knowles et al., 1992), which serve to restrict gene flow among populations and therefore are expected to promote population differentiation.
Most of previous studies on DNA variation of Eurasian Larix species focused on non-coding and anonymous regions of nuclear genome and included only a single or very few species (Semerikov et al., 2003; Kozyrenko et al., 2004). The objectives of this study are to clarify population differentiation of Eurasian Larix species and to verify their classification. For this purpose we investigated levels and patterns of nucleotide variation of two partial nuclear gene regions: 4–coumarate coenzyme A ligase (4CL) and coumarate 3- hydroxylase (C3H). The 4CL and C3H gene regions play a key role in general phenylpropanoid metabolism in lignin biosynthesis. We examined 19 populations representing five species and three varieties from the genus Larix (Fig. 1). Four of the investigated populations of L. sukaczewii (1D ~ 5A) came from previously glaciated area in northwestern Russia. The other two populations (6B and 7A) came from putative refugia in southern Urals. Populations 10A and 11A of L. sibirica included in our study occur in or near putative refugia in southern Siberia. Populations of L. cajanderi (13A, 13B and 13C) are located near putative refugia in western Beringia. The remaining populations came from ice free areas in central and southeastern Siberia, Sakhalin Island, Kamchatka Peninsula and central Japan.
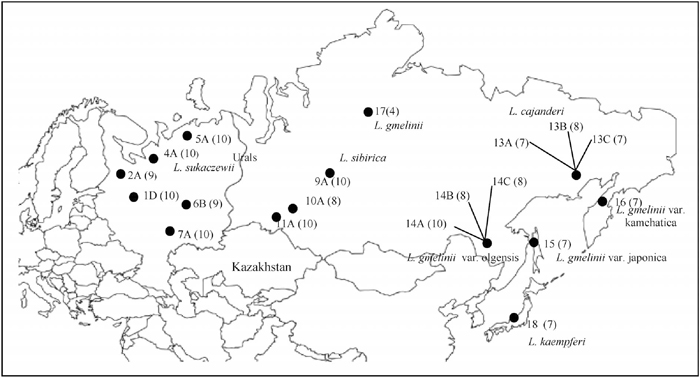 View Details | Fig. 1 Locations of the investigated Larix species from Eurasia, numbers of individuals are shown in parentheses near each population. |
Seed samples were collected in 19 natural populations of the following five species and three varieties of Eurasian Larix species: L. sibirica, L. sukaczewii, L. cajanderi, L. gmelinii, L. gmelinii var. japonica, L. gmelinii var. kamchatica, and L. gmelinii var. olgensis (Abaimov et al., 2002). Seeds of L. kaempferi were collected in Japan. In this study we adopted classification proposed by Abaimov et al. (2002). Details about the locations and number of individuals of the investigated populations are presented in Fig. 1.
Megagametophytes, which represent haploid maternal tissue were isolated from germinating seed. Genomic DNA was isolated from megagametophytes using SDS method (Ish-Horowicz, 1989). Two partial gene regions (758 bp and 873 bp in alignment length) were amplified for the 4CL and C3H genes respectively. The PCR primers for the 4CL gene region were the same as those reported by Wang et al. (2000). The forward primer 5’- CCAATCCTTTYTACAAGCCG- 3’ was located in exon 1 and the reverse primer 5’ - CGGGGAARGGCTYCTTTGC- 3’ was located in exon 2. Primers for the C3H gene region were designed based on DNA sequences of Pinus taeda from GeneBank using primer3 program (http://fokker.wi.mit.edu/primer3/input-030.htm) (Rozen and Skaletsky, 2000). The forward primer 5’- CGAGCATTCCCTATCTCC-3’ was located in exon 1 and the reverse primer 5’ - AACAAGCCCTGGATTCTCTG- 3’ was located in exon 2. The PCR mixture was prepared to the total volume of 50 μl containing 50–100 ng DNA template, 50 mM KCl, 10 mM Tris-HCl pH 8.3, 1.5 mM MgCl2, 2.5 pM of each primers and 200 μM of each dNTP and 1 unit of Taq polymerase. Amplification was carried out as follows: 95°C for 3 min. followed by 35 cycles of 30 sec. at 95°C, 30 sec. at 55°C for annealing, 90 sec. at 72°C, and finally with 7 min. at 72°C for further extension. PCR products were purified using WizardR SV Gel and PCR Clean-Up System (Promega, USA). The purified products were directly sequenced using the BigDyeTM Terminator (v 3.1) and ABI Prism 3100 automatic sequencer (Applied Biosystems, Foster City, CA, USA). Sequences obtained in this study have been deposited in the GenBank database under accession numbers EU280809 through EU280840 and EU280841 through EU280886 for the C3H and 4CL gene regions respectively.
Three copies of the 4CL gene (4CL-A1, 4CL-A2 and 4CL-B) exist in some Larix species (Wei and Wang, 2004a). However, we have several reasons to believe that the 4CL primers used in our study amplified only one particular copy (4CL-B) of the 4CL gene region. We compared the 4CL sequences obtained in our study with those reported by Wei and Wang (2004a). We found that the 4CL-A1 and 4CL-A2 copies are similar to each other and differ by only 14 nucleotides. On the other hand, they are highly diverged from the 4CL-B copy. The alignments between 4CL-A1 vs. 4CL-B and 4CL-A2 vs. 4CL-B copies differed by more than 50 nucleotides. Some of our sequences (28 out of 159) were identical with the 4CL-B region reported by Wei and Wang (2004a), while others differed by no more than 18 nucleotides. On the other hand, they differed from the other two copies by more than 50 nucleotides. In another study by Wang et al. (2000) where the same primers were used only one copy of the 4CL gene was detected in L. gmelinii. We compared the 4CL sequences obtained in that study with those reported by Wei and Wang (2004a) as well as with those obtained in our study. We found that similar to our results, the sequences obtained by Wang et al. (2000) represented the 4CL-B copy. Furthermore, if multiple and diverged copies were present in our material, we would expect to observe multiple peaks during sequencing using ABI 3100 sequencer, such as those reported by e.g., Gernandt and Liston (1999). Yet we did not observe such peaks. We therefore believe that direct sequencing method used in our study detected only one copy (4CL-B) of the 4CL gene region. Based on our data alone we cannot determine the reason why only this copy was amplified in our study and in that by Wang et al. (2000). Nevertheless, such selective amplification has been often reported in other studies and its possible causes have been reviewed by e.g., Wagner et al (1994).
DNA sequences obtained for each gene region were checked and assembled using the ATGC program ver. 4 (GENETYX CORPORATION). Sequence alignments were done with CLUSTAL X program (Thomson et al., 1997) and adjusted manually using BioEdit program (Hall, 1999). DnaSP program ver. 4.0 (Rozas et al., 2003) was used to estimate the level of nucleotide diversity (π) (Nei, 1987) and nucleotide polymorphism (θ) (Watterson, 1975). Fixation index values (FST) (Hudson et al., 1992) were calculated without gaps using the Proseq program ver. 2.9 (Filatov, 2002) with 10,000 permutations to determine significance level. To infer relationships among haplotypes neighbor-joining (NJ) trees (Saitou and Nei, 1987) were constructed including gaps using CLUSTAL X program (Thomson et al., 1997), bootstrap values were calculated from 1000 replicates.
The sequences of haplotypes for 159 individuals were obtained for the two partial gene regions: 758 bp of the 4CL region and 873 bp of the C3H region (following alignment). In the 4CL region, there were 21 segregating sites and 26 haplotypes (Table 1). The first nucleotide position was numbered as +1 in both gene regions. In exon 1 (1 ~ 654 bp), there were 18 segregating sites including 13 synonymous substitutions and five non-synonymous substitutions (positions 244, 270, 454, 485 and 547). There were only one segregating site and two indels (insertion or deletion) in the intron (655 ~ 736 bp). On the other hand, there were no segregating sites in exon 2 (737 ~ 758 bp). In the C3H region, there were 17 segregating sites and 17 haplotypes (Table 2). In exon 1 (1 ~ 187 bp) there were five segregating sites including four synonymous substitutions and one non-synonymous substitution (position 70). Eleven segregating sites were found in the intron (188 ~ 588 bp). There was only one synonymous substitution in exon 2 (589 ~ 873 bp). No indels were detected in the C3H gene region.
 View Details | Table 1 Summary of polymorphic sites in the 4CL gene region including 26 haplotypes with 21 segregating sites, nucleotide positions relative to the beginning of the sequence are indicated by digits on the top, – indicates indels, * indicates nonsynonymous substitution |
 View Details | Table 2 Summary of polymorphic sites in the C3H gene region including 17 haplotypes with 17 segregating sites, nucleotide positions relative to the beginning of the sequence are indicated by digits on the top, * indicates nonsynonymous substitution |
Measures of nucleotide variation, nucleotide diversity (π) and nucleotide polymorphism (θ) were similar, so only π values for both gene regions are presented (Table 3). Total nucleotide diversity at all sites (πT) in the 4CL region varied between 0.0007 in L. gmelinii and L. kaempferi to 0.0042 in L. gmelinii var. japonica. Nucleotide diversity at silent sites (πsil) varied between 0.0020 in L. gmelinii to 0.0116 in L. gmelinii var. japonica. In the C3H region πT varied between 0.0011 in L. kaempferi to 0.0038 in L. gmelinii var. japonica and πsil varied between 0.0019 in L. kaempferi to 0.0066 in L. gmelinii var. japonica.
 View Details | Table 3 Summary of nucleotide diversity (π) in the 4CL and C3H gene regions |
The values of FST for all pairs of populations were estimated by the method of Hudson et al. (1992) (Table 4). In the 4CL region, the FST values varied among pairs of populations and ranged between –0.069 (populations 1D-2A) to 0.800 (populations 17-18). In the C3H gene region, FST values ranged between –0.145 (populations 13C-14A) to 0.823 (populations 6B-18). In both regions, high and statistically significant FST values (> 0.15) were found for some pairs of populations of the same species e.g., L. sukaczewii (populations 1D-7A) and L. cajanderi (populations 13A-13B). There were also significant FST values between some species e.g., between population 7A of L. sibirica from putative Larix refugium in southern Siberia and most populations of other species included in our study. Significant FST values were also often observed in comparisons of L. gmelinii and L. kaempferi with other species. On the other hand, no population differentiation was observed among most L. sukaczewii populations except for comparisons involving population 7A. Similarly, weak population differentiation was observed among populations of L. gmelinii var. olgensis and in comparisons of this variety with populations of L. gmelinii var. japonica and L. gmelinii var. kamchatica.
 View Details | Table 4 Fixation index (FST) values for the two nuclear gene regions, 4CL (below diagonal) and C3H (above diagonal) for all pairwise population comparisons |
Relationships of the haplotypes for the 4CL and C3H regions were inferred using the neighbor-joining trees (Fig. 2 and Fig. 3 respectively). In both gene regions the haplotypes differed by only small number of mutational steps and except for L. kaempferi did not form groups corresponding to the taxonomic classification of the investigated species. In the 4CL region there were 26 haplotypes (designated as 4CL-H1 ~ H26) (Table 5), while in the C3H region there were 17 haplotypes (designated as C3H-H1 ~ H17) (Table 6). Some haplotypes e.g., 4CL-H15 and C3H-H14 were found in most populations. Haplotypes 4CL-H10, H12 and H13 and C3H-H10, H12 and H13 were found in some populations of L. sibirica and L. sukaczewii but were absent in the remaining species, while haplotypes 4CL-H7, H8, H16 ~ H18 and 4CL-H20 ~ H26, C3H-H1 ~ H7 and C3H-H15 ~ H17 were absent in L. sibirica and L. sukaczewii but present in the remaining species. On the other hand, some other haplotypes were species specific. For instance, haplotypes 4CL-H1, H2, H6, 4CL-H9 and C3H-H11 were specific to L. sukaczewii. Two of them (4CL-H2 and 4CL-H6) were rare, while haplotypes 4CL-H9 and C3H-H11 had high frequency in population 7A. Haplotypes 4CL-H3 and C3H-H8 were specific to populations 10A and 11A of L. sibirica respectively. Haplotypes 4CL-H5 and 4CL-H22 were specific to L. cajanderi, while haplotypes 4CL-H23, 4CL-H24 and C3H-H15 and C3H-H16 were specific to L. kaempferi. Haplotypes 4CL-H21 and C3H-H17 were specific to L. gmelinii, while haplotype C3H-H17 was shared only with L. gmelinii var. japonica.
 View Details | Fig. 2 Neighbor joining tree for the 4CL gene region, the numbers shown on the branches are bootstrap values based on 1000 replicates. |
 View Details | Fig. 3 Neighbor joining tree for the C3H gene region, the numbers shown on the branches are bootstrap values based on 1000 replicates. |
 View Details | Table 5 Haplotypes found in Larix populations in the 4CL gene region |
 View Details | Table 6 Haplotypes found in Larix populations in the C3H gene region |
Most of the previous studies suggested that Larix has lower nucleotide diversity compared to other genera of the family Pinaceae (LePage and Basinger, 1995; Wang et al., 2000). However, the level of nucleotide diversity revealed in our study was in the same order or slightly higher than those reported for other conifers e.g., Cryptomeria japonica (πsil = 0.0039) over seven loci (Kado et al., 2003), Cathaya argyrophylla (πsil = 0.0024) (Wang and Song, 2006), Pinus tabuliformis, P. yunnanensis and P. densata (πsil = 0.0087 ~ 0.0128 over 7 loci, (Ma et al., 2006). It thus appears that contrary to previous suggestions Larix species have levels of nucleotide diversity comparable to other conifers.
Certain reproductive characteristics such as pollen lacking air sacks and low seed viability suggest that Larix species are likely to have high levels of population differentiation (Duncan, 1954; Knowles et al., 1992). Yet, most of the previous studies suggested weak population differentiation in several Larix species (Fins and Seeb, 1986; Lewandowski et al., 1991; Timerjanov, 1997; Jaquish and El-Kassaby, 1998; Semerikov et al., 1999, 2003; Semerikov and Lascoux, 2003; Wei and Wang, 2003, 2004a,b; Larionova et al., 2004). Taking into account huge size and heterogeneous climatic history of Eurasia and the aforementioned reproductive features of Larix species these results are surprising. Contrary to the previous studies, we often found high levels of population differentiation. Significant FST values were found for pairs of populations of the same species e.g., populations 13A-13B of L. cajanderi and populations 6B-7A of L. sukaczewii. Significant FST values were also found between populations of different species e.g., between L. sukaczewii and L. sibirica, L. cajanderi and L. gmelinii var. olgensis and between L. gmelinii and L. kaempferi. These results indicate that in contrast to previous suggestions some Eurasian Larix species are diverged and populations of individual species are often highly differentiated even if they are separated by short distances e.g., populations 13A-13B of L. cajanderi, which are separated by less than two kilometers.
During the last glacial maximum (LMG) Eurasian Larix species survived in many distant and isolated refugia (Kremenetski, 1994; Tarasov et al., 2000; Andreev et al., 2002). Some of these refugia were located in southern Siberia (where populations 10A and 11A are located), south of Urals (where populations 6B and 7A are located) and in the Russian Far East (where populations 13A, 13B and 13C are located). Refugial populations are expected to be more differentiated, because they are likely to have evolved for a long time in isolation from each other. High population differentiation revealed in our study suggests that the present distribution of Larix species is a result of independent expansion and population mixing events involving multiple and genetically differentiated refugia.
Northwestern Russia, where populations 1D ~ 5A of L. sukaczewii occur, was heavily glaciated during LGM. Therefore, extant populations of Larix occurring in this region must be of recent, postglacial origin. This is consistent with our results, which showed that populations 1D ~ 5A are not differentiated because they share some haplotypes and the frequencies of these haplotypes are similar. Moreover, in both 4CL and C3H regions populations 1D ~ 5A shared some haplotypes with population 6B. They also had low FST values in comparisons involving this population. On the other hand, they showed significant FST values in comparisons with population 7A. In fact, population 7A also showed significant values in most comparisons with populations of other species included in our study. Populations 6B and 7A are located in southern Urals, which have often been suggested as Larix refugia during LGM (Kremenetski, 1994; Tarasov et al., 2000). Similarities between population 6B and populations 1D ~ 5A suggest that the part of the southern Urals where population 6B is located could be one of the sources of postglacial expansion of L. sukaczewii into the northwestern Russia. At the same time however, distinct character of population 7A from the same region indicates that populations in southern Urals are highly differentiated and that some of them did not contribute to post-glacial expansion of L. sukaczewii into northwestern Russia. It is also possible that the extant populations in northwestern Russia have originated from other refugial areas such as those near the Sea of Azov, which are now dominated by steppe vegetation or desert (Tarasov et al., 2000).
Currently L. gmelinii occurs in central Siberia. We found that it harbored a unique haplotype 4CL-H21, which was absent in all other species included in our study. It also harbored another haplotype C3H-H17, which was absent in other species except L. gmelinii var. japonica. Furthermore, it showed significant FST values in comparisons with other species included in our study. These results suggest that the extant populations of L. gmelinii may have different origin from other Eurasian Larix species. The sources of postglacial expansion of L. gmelinii could be refugia located in the Russian Far East, which at the time of glaciations was only locally glaciated (Porter et al., 1983; Ananyeyev et al., 1993; Bennett, 1997). Fossil data also indicate that Larix species were present in this region during LGM (Kremenetski, 1994; Tarasov et al., 2000; Andreev et al., 2002). Postglacial expansion of L. gmelinii from the Russian Far East has been suggested by Semerikov et al. (1999). However, population of L. gmelinii included in our study was very different from populations of L. cajanderi located in the Russian Far East. It is therefore possible that the extant populations of L. gmelinii in central Siberia originated from other parts of Eurasia. Unfortunately, we have only a single population of L. gmelinii from central Siberia. Therefore, based on our data alone we can not infer more detailed picture of migration history of this species.
There is no consensus regarding the number of Larix species in Eurasia and taxonomic position of some species is still an open and controversial issue. Many authors rejected status of L. sukaczewii as a species, declaring that L. sukaczewii and L. sibirica can not be distinguished (Bobrov, 1978; Milyutin and Vishnevetskaia, 1995). However, there is evidence showing that L. sukaczewii differs from L. sibirica in some morphological and biochemical features (Dylis, 1947, 1981; Milyutin et al., 1993; Abaimov et al., 1998; Bashalkhanov et al., 2003). We found that some haplotypes were unique to either L. sukaczewii or L. sibirica. For instance, haplotypes 4CL-H1, H2, H9 and 4CL-H14 were found in some populations of L. sukaczewii but were absent in all populations of L. sibirica. On the other hand, haplotype 4CL-H3 was found only in L. sibirica (population 10A) and absent in populations of L. sukaczewii. Similarly, haplotype C3H-H8 was present in population 11A of L. sibirica, while it was absent in populations of L. sukaczewii. On the other hand, haplotypes C3H-H9 and H11 were observed in some populations of L. sukaczewii but were absent in populations of L. sibirica. These differences in haplotype composition are in favor of the classification of L. sukaczewii and L. sibirica as different species. However, we also found low FST values in comparisons between populations of these two species e.g., population pairs 1D-9A and 2A-9A. Furthermore, some populations of L. sukaczewii and L. sibirica (7A and 10A respectively) were significantly different from other populations of the same species. These results indicate that although some populations currently occurring in western and central Siberia may indeed belong to two or more different species, the current classification recognizing difference only between western and central Siberia (e.g., Semerikov and Lascoux, 2003) is too simplistic and that further studies including additional populations especially from southern Urals and south-central Siberia are necessary to elucidate taxonomic status of L. sukaczewii and L. sibirica.
In central and eastern Siberia some authors recognized only a single species: L. gmelinii (Milyutin and Vishnevetskaia, 1995). Farjon (1990) had also recognized L. gmelinii in this region but with some varieties. On the other hand, other authors recognized there two different species: L. cajanderi and L. gmelinii (Bobrov, 1978; Abaimov et al., 1998, 2002). Our results show that there were differences in haplotype composition between L. cajanderi and L. gmelinii. Larix cajanderi harbored seven 4CL haplotypes and six of them (4CL-H4, H5, H11, H15, H16 and 4CL-H22) were absent in L. gmelinii. In the C3H region it harbored four C3H haplotypes and two of them (C3H-H1 and C3H-H9) were absent in L. gmelinii. Furthermore, the FST values between populations of L. cajanderi and L. gmelinii were higher than those within species. In the 4CL and C3H regions FST values between L. cajanderi (population 13B) and L. gmelinii were 0.398 and 0.619 respectively. FST values within L. cajanderi (between populations 13A-13B) were 0.163 and 0.220 in the 4CL and C3H regions respectively. These results support treatment of L. cajanderi and L. gmelinii as two different species. Similar to L. sukaczewii and L. sibirica, in L. cajanderi we also found high levels of population differentiation within very limited geographic area, which corresponds to LGM Larix refugium in the Russian Far East (Porter et al., 1983; Ananyeyev et al., 1993; Bennett, 1997). This further strengthens our suggestion that more intensive sampling especially from known refugial regions is necessary for correct classification of Eurasian Larix species and for inferring their postglacial migration.
Some authors regarded L. gmelinii var. olgensis as a separate species (Bobrov, 1978; Liu et al., 2006). However, other authors have argued that L. gmelinii var. olgensis is just a variety of L. gmelinii (Shi et al., 1998; Abaimov et al., 2002). Our results show that L. gmelinii var. olgensis has different haplotype composition than L. gmelinii. For instance, haplotypes 4CL-H14 ~ H16 and C3H-H3 and C3H-H7 were found in L. gmelinii var. olgensis and had high frequency but were absent in L. gmelinii. On the other hand, haplotypes 4CL-H21 and C3H-H17 were found in L. gmelinii but were absent in L. gmelinii var. olgensis. In addition, we found significant FST values in comparisons of population of L. gmelinii with populations of L. gmelinii var. olgensis as well as with populations of two other varieties of this species: L. gmelinii var. japonica and L. gmelinii var. kamchatica. These results indicate that L. gmelinii from central Siberia is very different from its varieties occurring in the Russian Far East and Kamchatka Peninsula. Unfortunately, our sampling of L. gmelinii was very limited. Therefore, further studies including additional populations are necessary to determine relationships of this species and its varieties.
Phylogenetic studies suggested that Japanese larch L. kaempferi is closely related to L. olgensis (regarded as L. gmelinii var. olgensis in our present study) (Semerikov et al., 2003; Wei and Wang, 2003, 2004a). However, results of the present study do not support this suggestion. We found that in both gene regions haplotype composition of L. kaempferi differed from all other species included in our study. This was reflected in statistically significant FST values in comparisons with all other species indicating that L. kaempferi is among the most diverged Eurasian Larix species.
We would like to thank Drs Ove Martinsson JiLU, Bispgården, Sweden and Katsuhiko Takata, Institute of Wood Technology Akita Prefectural University, Japan for providing seed samples. We also thank the two anonymous reviewers for helpful comments on this manuscript. This work was financially supported by the grants No. 13575002 and 17405032 to AES from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
|