| Edited by Hiroshi Iwasaki. Masahiro Terasawa: Corresponding author. E-mail: mterasw@iwate-nurse.ac.jp |
Meiotic recombination is crucial for ensuring chromosome segregation in meiosis I, and it results in the production of a variety of genomes. Recombination in the budding yeast Saccharomyces cerevisiae is initiated by the introduction of DNA double-strand breaks (DSBs) by Spo11p at specific sites on meiotic chromosomes (Keeney et al., 1997). After cleavage, Spo11p is attached covalently to the 5’ ends of the DSB, and following its removal single-stranded tails are produced at the DSB ends. These tails then carry out a homology search with the assistance of the RecA-like proteins Rad51 and Dmc1 (Bishop et al., 1992; Shinohara et al., 1992). After the formation of recombination intermediates, recombinant molecules are produced by strand exchange, DNA strand resolution, and meiotic recombination-related DNA synthesis (MRDS) (Terasawa et al., 2007).
During recombination, the Mre11, Rad50, and Xrs2 proteins play an important role in the introduction of DSBs and the resection of DSB ends. These proteins form a complex termed the MRX complex. Usui et al. (1998) demonstrated that Mre11p physically interacts with Rad50p and Xrs2p, and that Mre11p colocalizes with Rad50p or Xrs2p on meiotic chromosomes. Mre11p requires many other factors for the introduction of DSBs (Borde et al., 2004), and it exhibits double-stranded (ds) DNA-dependent 3’-5’-exonuclease and single-stranded (ss) DNA-dependent endonuclease activities in vitro. The MRX complex is thought to play a role in the resection of DSB ends. Association of Spo11p with short (24–40 or 10–15 bp) oligonucleotides in meiosis (Neale et al., 2005) suggests that Mre11p removes Spo11p by resecting the ssDNA adjacent to the 5’ ends, which allows the ends to be processed. The DSB ends are resected to produce 3’ ss tails of approximately 600 bp in length (Bishop et al., 1992). Recently, we demonstrated that MRDS of approximately 0.8 or 0.8–1.9 kb in length occurs to complete recombinant formation (Terasawa et al., 2007). In addition, Rad50p, a component of the MRX complex, belongs to the structural maintenance of chromosomes family and probably has a role in binding DNA ends and holding them in close proximity (D’Amours and Jackson, 2002). Xrs2p is required for the translocation of Mre11p to the nucleus and it regulates the functions of the MRX complex in meiotic recombination and telomere maintenance (Shima et al., 2005; Tsukamoto et al., 2005).
The rad50S, mre11-H125N, and sae2/com1Δ mutants are defective in the formation of ssDNA at DSB ends, and they accumulate DSB ends during meiotic recombination (Alani et al., 1990; McKee and Kleckner, 1997; Prinz et al., 1997; Moreau et al., 1999). The rad50S mutant was shown to possess separation-of-function phenotypes and to be defective in spore formation (Alani et al., 1990). In rad50S cells, Spo11p remains bound to the 5’ ends of DSBs and prevents ssDNA production, even when cells in meiosis are returned to mitotic conditions. The survival rate of the rad50S mutant cells drops sharply and no recombinants are formed. Spo11p is suggested to attach to the 5’ ends and protect them from processing after DSB formation (Zenvirth et al., 1997). The Mre11-H125N protein has a point mutation in a conserved region of phosphoesterase motif III, and it is defective in ssDNA endonuclease and dsDNA 3’-5’-exonuclease activities (Krogh et al., 2005). The SAE2/COM1 gene was identified by analysis of mutants that show separation-of-function phenotypes. These mutant cells fail to form spores and show a weak sensitivity to MMS as compared with wild type cells (McKee and Kleckner, 1997; Prinz et al., 1997).
Mec1p and Tel1p phosphorylate Sae2p, which is required for the DNA damage response and DSB end processing in meiosis (Baroni et al., 2004; Cartagena-Lirola et al., 2006). Although phosphorylation of putative phosphorylation sites near the C terminus is required for meiotic recombination, phosphorylation of sites near the N terminus is not (Cartagena-Lirola et al., 2006). In addition, the defective single-strand annealing activity of the rad50S mutant is suppressed by the presence of multiple copies of SAE2 (Clerici et al., 2005). It has also been suggested that Sae2p controls DSB ends for mitotic recombination by regulating the nuclease functions of the MRX complex (Rattray et al., 2001). These results suggest that Sae2p functionally cooperates with the MRX complex.
In this study, to elucidate how Sae2p is related to the MRX complex for DSB resection in meiotic recombination, we examined the role of Sae2p phosphorylation. We constructed sae2 mutants with substitutions of putative phosphorylation sites. We found that phosphorylation is necessary for both the initiation of resection and for efficient resection on meiotic chromosomes. The function of C-terminal phosphorylation is distinct from that of N-terminal phosphorylation. Phosphorylated Sae2p was shown to colocalize with the MRX complex. We discuss the role of Sae2p phosphorylation in the regulation of meiotic processes by collaboration with the MRX complex.
To create pCM136, a 1.6-kb SAE2 gene fragment of S. cerevisiae was amplified by PCR and cloned into pRS318. sae2 alleles bearing point mutations were subcloned into pYT1610 (Tsukamoto et al., 2005). pCM137 encodes full length Sae2p tagged with 9Myc at the C terminus.
To construct sae2 point mutation alleles in S. cerevisiae, the SAE2 gene was replaced with the URA3 marker by PCR-mediated one-step gene disruption. CMY165 is MATα sae2Δ::URA3, ho::LYS2, trp1, ura3, lys2, his4X::LEU2, arg4-nsp and CMY166 is MATa sae2Δ::URA3, ho::LYS2, trp1, ura3, lys2, his4B::LEU2, arg4-bgl. CMY165 and CMY166 are SK1 derivatives. The SAE2-9MYC and sae2 mutant alleles were targeted to the MET17 locus in CMY166 using the integration plasmid pYT1610.
The SAE2-9MYC allele was constructed by replacing the C terminus of the SAE2 gene in pYT1610 in CMY165 with a 9Myc-klTRP1 unit (Knop et al., 1999), using PCR-mediated one-step gene replacement. For construction of phosphorylation-defective mutants, pCM137 was used to create alanine substitutions (TAKARA). These sets of mutations were subcloned into the integration vectors and introduced into the sae2Δ strain.
The mre11-H125R mutant with separation-of-function phenotypes was isolated by screening a randomly mutagenized mre11 mutant gene pool. To replace the wild type endogenous MRE11 gene with mre11-H125R, a two-step integration method was used as previously described (Tsubouchi and Ogawa, 1998).
Approximately 3 × 108 cells per sample were collected. The samples were lysed as described (Burke et al., 2000), separated by 8% SDS-PAGE and analyzed by western blotting using ECL plus detection reagents (Amersham). For western blotting, anti-Myc monoclonal antibody (SIGMA) was used at 1/1000 dilutions. The Luminescent Detection kit (Roche Diagnostics) and a LAS3000 chemiluminescence image analyzer (Fujifilm) were used for detection.
Spread meiotic chromosomes were prepared as described (Bishop, 1994). Sae2p and Mre11p were detected using a mouse anti-Myc monoclonal antibody and a guinea pig anti-Mre11 antibody (Usui et al., 1998). Images were taken with a Zeiss epifluorescence microscope (Zeiss Axiovert 200, 100× objective with NA = 1.4) equipped with a cooled CCD camera (SPOT; Diagnostic Instruments). Image sources were recorded as monochrome data using CCD image capture with IP Lab-PVCAM software (Solution Systems). Random distribution was calculated as described in Novak et al. (2001).
Genomic DNA purified from each strain was digested with PstI, electrophoresed in an 0.8% TAE agarose gel, transferred to a nylon membrane (Amersham), and hybridized with a 1.6-kb probe bearing his4 DNA. The probe was labeled with digoxigenin, using a PCR DIG Probe Synthesis kit (Roche Diagnostics) and the primer pairs 5’-GATGGCCTGTTTACCGTCCTCCAT-3’ and 5’-GGACTCTGGACCTCTTGGATCTGT-3’. Southern blot signals were quantified with a DIG Luminescent Detection kit (Roche Diagnostics) and a LAS3000 chemiluminescence image analyzer (Fujifilm).
To examine the roles of Sae2p phosphorylation in the resection of DSBs, we constructed mutants with alanine substitutions of putative Mec1/Tel1 phosphorylation sites (Fig. 1). Sae2p has five (S/T)Q motifs that are putative Mec1/Tel1 phosphorylation sites. Three phosphorylation-defective mutants were made by alanine substitution of two sites near the N terminus (sae2-2A), three sites near the C terminus (sae2-3A) and all five sites (sae2-5A) (sae2-2A is abbreviated as 2A, and the other mutants are similarly abbreviated). The wild type and mutant proteins were tagged with 9Myc at the C termini. In the SAE2-9MYC strain, approximately 90% of spores were viable, indicating that the tag had little effect on meiosis (Table 1). In the phosphorylation-defective mutants, the mutant proteins could also be detected as readily as in the SAE2-9MYC strain (Fig. 2A). More slowly migrating bands were observed for SAE2-9MYC, 2A and 3A, but not 5A. These results are consistent with those of Cartagena-Lirola et al. (2006).
 View Details | Fig. 1 Diagrams of the constructed sae2 mutants and wild type strain. Asterisks on the gray bars indicate the putative phosphorylation sites in Sae2p. Crosses on the gray bars indicate mutations sites of these sites. Numbers show amino acid residue numbers for phosphorylation sites and for the C terminus of Sae2p. A wild type and 3 phosphorylation-defective mutants were tagged with 9Myc at the C termini, as indicated by ellipses. |
 View Details | Table 1 Frequencies of spore formation and viability |
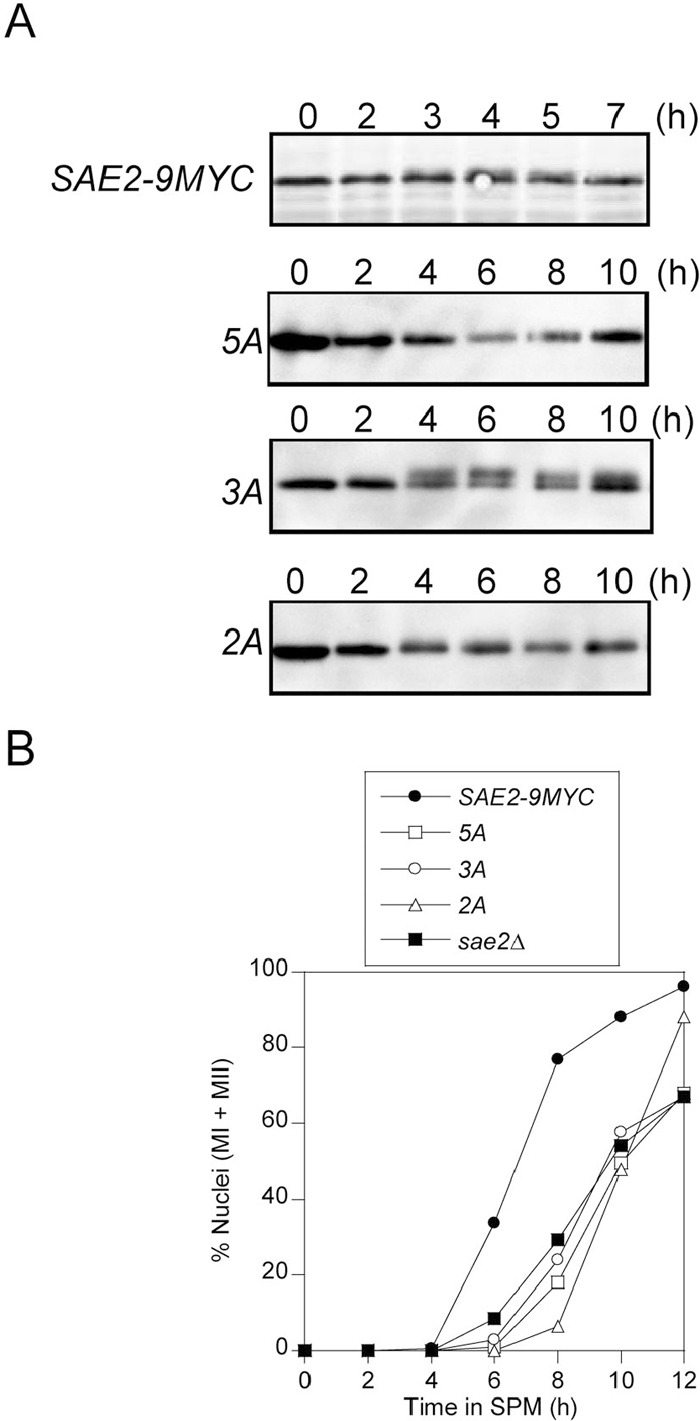 View Details | Fig. 2 (A) Expression of Sae2-9Myc and mutant proteins in meiosis. Meiotic cells were harvested at the indicated time, and cell extracts were prepared and analyzed by western blotting using an anti-Myc antibody. (B) The progression of meiotic nuclear divisions in the SAE2-9MYC strain and in sae2 mutants. Cells in SPM were collected at the indicated times, and those undergoing the first meiotic division (MI) or having completed both divisions (MII) were detected by staining with DAPI and are shown as percentages of total cells. More than 200 cells were examined at each time point. ●, SAE2-9MYC; □, 5A; ○, 3A; △, 2A; ■, sae2Δ. |
The efficiencies of the meiotic nuclear divisions (I and II) of these mutants were examined. Approximately 80% of SAE2-9MYC cells completed both meiotic divisions by 8 h, indicating that the progression of meiosis in this strain is similar to that in wild type. For the 3A and 5A mutants, meiosis was delayed by 3–4 h as compared with that of the SAE2-9MYC strain, and spores were rarely formed (Fig. 2B and Table 1). The progression of nuclear division and the extent of spore formation in these mutants were similar to those of a sae2Δ mutant. In contrast, although nuclear division in the 2A mutant was delayed by 2–4 h as compared with that of SAE2-9MYC cells, 85% of the cells underwent meiosis I and II by 12 h, and approximately 30% of the cells formed spores at 24 h (Fig. 2B and Table 1). Except for the delay in nuclear division in 2A cells, the properties of the phosphorylation-defective mutants are consistent with those observed by Cartagena-Lirola et al. (2006). Phosphorylation of the C-terminal region appears to be required for spore formation and meiotic nuclear divisions.
DSB resection in the mutants was examined by Southern blotting. Genomic DNA was purified from the SAE2-9MYC strain and from mutant cells at various times after the induction of meiosis, and DSBs at the his4-LEU2 locus on chromosome III (Cao et al., 1990) were then visualized by Southern blotting (Fig. 3A). In SAE2-9MYC cells, smear bands were observed at 4 h and 6 h, and they disappeared at 8 h, similar to the pattern observed for the wild type strain. In both 3A and 5A mutants (Fig. 3B, C), discrete DSB bands were observed and these persisted for at least 12 h. On the other hand, the 2A mutant formed discrete bands at 4 h and 6 h, as observed for the sae2Δ mutant. These bands were smeared at 8 h and finally disappeared at 10 h (Fig. 3B and C). The patterns at 4 h and 6 h for 2A cells were distinct from those in SAE2-9MYC cells, which were characterized by smeared bands (Fig. 3D). These results indicate that the timing of DSB formation in the three phosphorylation-defective mutants was same as in wild type cells, but the timing of the initiation of resection was delayed by at least 4 h in the 2A mutant. These results reveal that phosphorylation of the C-terminal sites is critical for resection and that phosphorylation of the N-terminal sites increases the efficiency of resection initiation.
 View Details | Fig. 3 Detection of DSB bands at the his4-LEU2 locus in sae2 mutants. (A) Restriction maps of the his4-LEU2 region and locations of two DSB sites and the probe. (B) Meiotic cells were harvested at the indicated times, and genomic DNA was purified, digested with PstI and analyzed by Southern blotting. DSB bands in the SAE2-9MYC strain and in the phosphorylation-defective mutants 2A, 3A, 5A, and sae2Δ are shown. (C) Quantitative analysis of DSB bands shown in (B) for the SAE2-9MYC strain and the mutants. ●, SAE2-9MYC; □, 5A; ○, 3A; △, 2A; ■, sae2Δ. (D) Comparison of DSB bands between the SAE2-9MYC and 2A strains at 4 h. Smeared bands in SAE2-9MYC and discrete bands in 2A are indicated with solid lines and arrow heads, respectively. |
The MRX complex is observed as foci on chromosomes in the rad50S mutant, which accumulates DSBs, but it is not present in wild type strains (Usui et al., 1998). To determine whether Sae2p localizes on meiotic chromosomes and colocalizes with Mre11p, Sae2p and Mre11p focus formation was examined in rad50S cells and in the nuclease-defective mre11-H125R mutant, which is impaired in DSB resection. Nuclei from cells harvested at 4, 6, or 8 h in meiosis were examined by immunostaining of surface-spread chromosomes for Mre11p and Sae2p focus formation. Neither Mre11p nor Sae2p foci (Fig. 4A) were observed in SAE2-9MYC cells. On the other hand, Mre11p and Sae2p foci were detected in rad50S and mre11-H125R mutants (Fig. 4C, D). Thus, Sae2p foci are also detected in the resection-defective rad50S and mre11-H125R mutants. In addition, Mre11p focus formation was not dependent on the SAE2 gene (Fig. 4B).
 View Details | Fig. 4 (A)–(G): Localization of Mre11p and Sae2p on meiotic chromosomes in the mutants. Cells were harvested 4, 6 and 8 h after the introduction of meiosis; the chromosomes were spread on a glass slide and stained with anti-Mre11 and anti-Myc antibodies. Percentages of Sae2p (●) and Mre11p (▲) focus-positive cells represent the percentage of nuclei having more than five Mre11p or five Sae2p foci. At least 100 cells were scored at each time point. |
Mre11p and Sae2p focus formation was also examined in the phosphorylation-defective mutants. Mre11p and Sae2p foci increased from 4 h to 8 h in the 3A and 5A mutants, whereas Mre11p foci peaked at 6 h in the 2A mutant (Fig. 4E, F, G). Focus formation is independent of the phosphorylation of Sae2p, because Sae2p focus-positive cells were observed in all the phosphorylation-defective mutants.
Further, to clarify whether Sae2p phosphorylation is required for colocalization with the MRX complex, Sae2p and Mre11p foci on meiotic chromosomes were double stained in 2A, 3A, and 5A meiotic nuclei at 6 h and 8 h. At both time points, approximately 15% of 2Ap and 30% of 3Ap foci colocalized with Mre11p foci (random distribution: 6%) (Fig. 5A, B, C). However, only 5% of 5Ap foci colocalized with Mre11 foci, indicating that the distributions of the two proteins were independent (random distribution: 4%). These results demonstrate that phosphorylation of any Sae2p phosphorylation site is required for colocalization with Mre11p and that phosphorylation-negative Sae2p cannot colocalize with Mre11p. It is noteworthy that approximately 30% and 15% of Sae2p foci colocalized with Mre11p foci in mre11-H125R and rad50S mutants (random distribution: 4% and 6%), respectively (Fig. 5A, B, C).
 View Details | Fig. 5 Colocalization of Sae2p and Mre11p. (A) Representative chromosomes with Sae2p and Mre11p foci in the 5A, 3A, rad50S, and mre11-H125R mutants. Cells were harvested 8 h after the induction of meiosis; the chromosomes were spread on a glass slide and stained with anti-Mre11 (Green in Merged) and anti-Myc antibodies (Red in Merged). Arrowheads in Merged represent Sae2p foci colocalized with Mre11p. Bar; 5 μm. (B), (C) Percentage of colocalization of Sae2p and Mre11p foci in the mutants at 6 h and 8 h. At least 100 cells were scored. The frequencies are shown as means ± S.D. |
The pattern of resection of DSB ends was expected to vary among the different mutants analyzed here. We obtained information on DSB resection by examining whether the DSB ends could be utilized for recombinant formation in mitosis by a return-to-growth (RTG) experiment that assayed the arg4 locus. In rad50S and sae2Δ mutants, which are unable to remove Spo11p from DSB ends, no recombinants were formed and the recombination frequency did not increase in RTG. This result indicates that the DSB ends in these mutants are unsuitable structures for mitotic recombination. In SAE2-9MYC cells, recombinant formation increased with incubation time after introduction into meiotic conditions, and at 10 h it reached the maximum level of approximately 1000-fold of the spontaneous level.
Recombinant formation in 2A and 3A strains increased to 800- to 1000-fold of the spontaneous level in RTG, and the levels were similar to that of SAE2-9MYC cells. In contrast, recombinant formation increased by only approximately 30-fold in 5A cells. These results show that DSB ends in 2A and 3A cells, but not in 5A cells, can be utilized for recombinant formation in the mitotic cell cycle (Fig. 6A), which is consistent with the colocalization of these Sae2 proteins with Mre11p (Fig. 5A, B, C). The survival fractions of 3A and 5A mutants decreased gradually as meiosis proceeded and were similar to that of the sae2Δ mutant, while the survival fraction of the 2A mutant decreased moderately (Fig. 6B). These data suggest that both 2A and 3A, but not 5A mutants, have the potential to eliminate Spo11p from DSB ends. The 3A mutant is likely to be defective in recombinant formation after the removal of Spo11p.
 View Details | Fig. 6 Return-to-growth assay of sae2 mutants. Meiosis was induced in the indicated sae2 mutants. At various times after induction, cells were returned to vegetative growth media, and the fraction of Arg+ prototrophs was measured. (A) The fractions of recombinants at the arg4 locus were determined by dividing the number of colony-forming units (CFUs) on SC–Arg medium by the number of CFUs on yeast peptone dextrose (YPD) medium. (B) The fractions of survival were determined by dividing the number of CFUs on YPD at each time point by that of 0 h. ●, SAE2-9MYC; □, 5A; ○, 3A; △, 2A; ■, sae2Δ. |
Comparisons of the results obtained from cross-sectional experiments with each mutant are summarized in Table 2.
 View Details | Table 2 Comparisons of the results obtained from cross-sectional experiments with each mutant |
Sae2p is phosphorylated by the Tel1 and Mec1 protein kinases, and phosphorylation is involved in DNA damage signal transduction in mitosis and DSB repair in meiosis (Baroni et al., 2004; Cartagena-Lirola et al., 2006). In this study, we demonstrated that phosphorylation of Sae2p was essential for the progression of meiosis through cooperation with the MRX complex.
Sae2p localized on meiotic chromosomes when the resection of meiotic DSB ends was arrested in the rad50S and mre11-H125R mutants, and in the phosphorylation-defective sae2 mutants (2A, 3A and 5A) (Fig. 4). Sae2p colocalizes with the MRX complex in the 2A, 3A, rad50S and mre11-H125R mutants, although the frequencies in the 2A and rad50S mutants were low, suggesting that colocalization is relatively unstable in these mutants. Moreover, colocalization is disrupted by mutations affecting all phosphorylation sites (Fig. 5), indicating that phosphorylation of Sae2p regulates the function of the MRX complex at meiotic DSB ends because Mre11p colocalizes with Spo11p (Prieler et al., 2005). Sae2p might collaborate with the MRX complex to induce Mre11p nuclease activity for the removal of Spo11p. Alternatively, Lengsfeld et al. (2007) reported that Sae2p itself has an endonuclease activity that cooperates with the MRX complex, and that the presence of both Sae2p and the MRX complex enhances the endonuclease activity of Sae2p in vitro. Our data supports the above conclusion that Sae2p functions cooperatively with the MRX complex.
Phosphorylation of Sae2p at sites near the C terminus appears to have a role different from that played by phosphorylation near the N terminus. Phosphorylation of C-terminal sites is essential for the resection reaction. On the other hand, efficient initiation of the resection reaction requires phosphorylation of the N terminus. Moreover, the frequency of colocalization between Sae2p and Mre11p was high in C-terminal phosphorylation-defective mutants (3A), but it was lost in mutants with substitutions at all five phosphorylation sites (5A). This result suggests that phosphorylation of the N terminus enhances interactions with the MRX complex and improves resection initiation. In the C-terminal phosphorylation-defective mutant (3A), recombinants could be formed under RTG conditions, but the resection reaction was rarely promoted, suggesting that this mutant has residual activity for resection or that it can remove Spo11p from DSB ends, but with poor progression in resection. Thus, C-terminal phosphorylation may be required to initiate resection.
We have also constructed N- and C-terminal truncated mutants. Truncations of 70 amino acid residues at the N terminus and of 39 amino acid residues at the C terminus impaired DSB resection (unpublished data). RTG analysis of the truncated mutants showed that deletion of the C-terminal region severely reduced recombinant formation, similar to what is observed for the sae2Δ mutant, whereas recombinant formation in the N-terminally truncated mutant was at nearly wild type levels (unpublished data). These results suggest that the C-terminal region plays a critical role for the resection reaction in vivo.
It is interesting that the C-terminal region of Sae2p, including the phosphorylation sites, has homology with human CtIP, fission yeast Ctp1, and novel proteins in plants and nematode (Limbo et al., 2007; Penkner et al., 2007; Sartori et al., 2007; Uanschou et al., 2007). Human CtIP interacts with the Mre11-Rad50-Nbs1 complex and BRCA1, which are associated with genetic disorders, and it functions in DSB resection (Yu et al., 2006; Sartori et al., 2007). Our genetic analysis of Sae2p should contribute to the understanding of the general mechanism of homologous recombination and genetic diseases.
We thank Dr. Oshiumi for generously providing strains and discussions. We are grateful to Ms. C. Mitsuoka and Y. Fujikawa for technical assistance. This work was supported by a Grant-in-Aid for Specially Promoted Research to TO (14780530) and Grants-in-Aid for Young Scientists to YT (1770150) and Exploratory Research to MT (17657058) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
|