| Edited by Yoshibumi Komeda. Hiro-Yuki Hirano: Corresponding author. E-mail: hyhirano@biol.s.u-tokyo.ac.jp |
Plant architecture depends on morphological features of shoots, leaves, inflorescences, and flowers. Morphologies of leaves have diversified in angiosperms, as those of inflorescences and flowers. For example, leaves in monocots have parallel veins, whereas those in eudicots have reticulate veins. Many monocots such as rice and tulip have long and large lamina. To support long leaves, some plants like a Kaffir lily have elaborated thick and rigid leaves. By contrast, in plants with large thin leaves such as rice and maize, a strong structure, called midrib, are formed in the central region of the leaves to support them in being erect.
The DROOPING LEAF (DL) gene in Oryza sativa has two distinct roles in rice development (Nagasawa et al., 2003; Yamaguchi et al., 2004; Bommert et al., 2005). First, DL acts in carpel specification; a loss-of-function mutation in DL causes homeotic conversion of carpels into stamens. Second, DL functions in midrib formation; the dl mutant fails to make midrib, resulting in drooping leaf phenotype. At the initial stages of leaf development in wild type, a group of cells in the central region of leaf primordia proliferates along the abaxial-adaxial axis. The midrib structure is formed from a large number of the cells produced by this specific cell proliferation. Midrib is comprised of a large vascular bundle, a small adaxial bundle, and two locules, called ‘clear cells’ (Yamaguchi et al., 2004). The locules are thought to be generated from enough cells accumulating in the central region through programmed cell death. In the dl mutant, however, the specific cell proliferation in the central region does not occur, and subsequent processes such as programmed cell death and differentiation of the small adaxial bundle are no longer generated, resulting in lack in midrib structure. Thus, the specific cell proliferation regulated by DL function is essential for midrib formation.
Molecular cloning of DL revealed that it encodes a putative transcription factor with a zinc finger domain and a YABBY domain (Yamaguchi et al., 2004). During leaf development, DL is expressed in the central region of the leaf primordia (P1–P4), the presumptive region where the midrib structure will be formed later. In contrast, constitutive expression of DL promotes cell proliferation along the abaxial-adaxial axis throughout the leaf primordia and generates ectopic midrib-like structures in the lateral region of the leaves, suggesting that DL is sufficient to elaborate midrib in rice leaves. These results indicate that DL specifies the fate of the cells in the central region of the leaf primordia to induce cell proliferation along the abaxial-adaxial axis.
The rice and Arabidopsis genomes have eight and six members in the YABBY gene family, respectively (Siegfried et al., 1999; Bowman, 2000; Toriba et al., 2007). In the YABBY genes of Arabidopsis, the CRABS CLAW (CRC) gene is most similar to DL (Bowman and Smyth, 1999; Yamaguchi et al., 2004). The crc mutant, however, causes neither homeotic conversion in the flower or lack of midvein in the leaf (Alvarez and Smyth, 1999). Carpel specification is regulated by the AGAMOUS gene that encodes a MADS-box transcription factor in Arabidopsis, whereas its orthologs, OsMADS3 and OsMADS58, may have lost the function of carpel specification in rice evolution (Yanofsky et al., 1990; Yamaguchi et al., 2006). In some grasses, such as Pennisetum americanum (pearl millet) and Panicum aestivum, a mutation in a single locus shows defects in the leaf and the carpel, which are similar to the those observed in weak dl mutants of rice, suggesting that the gene corresponding to DL is involved in midrib formation and carpel development in these plants (Rao et al., 1988, Fladung et al., 1991). Thus, the function of DL and its homologs may have been acquired the function for midrib formation and carpel specification before the divergence of grass species.
The specific cell proliferation in the central region of leaf primordia is the first step for midrib formation. Cell differentiation and program cell death may be involved in the subsequent processes in the formation of midrib structure. It is of great interest to know what genes are regulated by DL and what genetic networks consisted of these genes are responsible for the early processes in midrib formation. Furthermore, it is also intriguing whether genes under the regulation of DL are shared by the genetic networks in midrib formation and by those in carpel specification. To address these questions, we compared expression profiles between the dl mutant and transgenic plants that are constitutively expressing DL by microarray analysis.
Taichung 65 (T65) was used as wild type for histological observation and for RNA isolation in RT-PCR analysis. The dl-sup1 mutant used in this study is a complete loss-of-function mutant that has a single nucleotide substitution at the RNA splicing site of the fifth intron of DL (Nagasawa et al., 2003; Yamaguchi et al., 2004). For overexpression, a construct (Act1:DL) in which DL cDNA was placed under the control of the rice Actin1 promoter was introduced into calli from the scutellum of T65 by Agrobacterium-mediated transformation (Hiei et al., 1994), and rice plants overexpressing DL were regenerated as described previously (Yamaguchi et al., 2004).
Total RNA was isolated from tissues described below using TRIZOL reagent (Invitrogen, Carlsbad) according to the manufacture’s instruction, and resulting RNA solution was treated with RNase-free DNase I (Takara, Kyoto). RNA was isolated from the shoot apices of about 1-cubic-millimeter including the leaf primordia from P1 to P6. The shoot apices were manually dissected from the seedlings of wild type, the dl-sup1 mutant (2 weeks after germination) and regenerated plants carrying Act1:DL (roughly similar size to that of the 2-weeks-old seedling of wild type).
To examine the expression patterns of DL in various tissues and organs, young panicles of 3–8 mm were used for inflorescences of wild type and the dl-sup1 mutant, in which floral organs are differentiating, and seedlings of 2 weeks after germination were used for the leaf and root of wild type. For RT-PCR analyses, first-strand cDNA was synthesized Superscript III (Invitrogen, Carlsbad) and the products were obtained by PCR amplification using primers listed in Table 1.
 View Details | Table 1 Primers used in this work |
Concentration of total RNA was determined with a spectrophotometer (Nano Drop ND-1000; Nano Drop, Wilmington) and integrity of total RNA was checked with Agilent 2100 Bioanalyzer (Agilent, Palo Alto). First-strand cDNA was synthesized from 400 ng of total RNA using MMLV-RT reverse transcriptase with an oligo (dT) primer containing a T7 polymerase promoter site at the 5’ end. Complementary RNA (cRNA) was synthesized and labeled with the fluorescent dyes Cy3 or Cy5 (Perkin-Elmer, Boston) using T7 RNA polymerase and the first-strand cDNA as a template. Agilent Low RNA Input Linear Amplification kit (Agilent, Palo Alto) was used to synthesize the labeled cRNA. The resulting cRNA was purified with Qiagen RNeasy Mini kit (Qiagen, Valencia) and use for hybridization.
For microarray analyses, Agilent 22K rice oligoarrays (Agilent, Palo Alto), which contain 22,000 features corresponding to the full-length cDNA of rice, was used (Kikuchi et al., 2003). Four sets of a color-swapping experiment were carried out using two independent biological samples. Hybridization was carried out with labeled cRNA at 60°C for 17 hours, and signals on the oligoarrays were scanned by Agilent Microarray Scanner (Palo Alto) after washing. Using Agilent Feature Extraction software (ver. 7.1), the signal intensity was measured from each spot in the array and normalized by the LOWESS method (Cleveland, 1979). Statistical analyses were carried out using Spotfire DecisionSite 8.0 package (Spotfire, Somerville).
To identify genes that are expressed under the control of DL and involved in early processes in midrib formation, we compared expression profiles in the shoot apex of the transgenic plants that strongly and constitutively expressed DL and those of the dl-sup1 mutants.
We produced transgenic rice plants (Act1:DL) overexpressing DL under the control of the rice Actin1 promoter that induce strong and constitutive gene expression in rice. RT-PCR analysis indicated that DL expression was enhanced in the Act1:DL plant compared to wild type (Fig. 1A). To examine the effect of DL overexpression, we observed the cross sections of the shoot apex of wild type, the Act1:DL plant, and the dl-sup1 mutant, which has null mutation (Fig. 1B). In wild type, the central region of the leaf primordia became thick gradually from P2 to P4, suggesting that the specific cell division occurred along the abaxial-adaxial axis in this region. In dl-sup1, however, thickness of the central region was reduced, suggesting that no such specific cell proliferation occurred. In contrast, constitutive expression of DL resulted in thickening leaf primordia not only in the central region but also in the lateral region. Together with the enhancement of DL expression, these observations indicated that the shoot apices of the Act1:DL plant are adequate enough to use subsequent analyses for expression profiles.
 View Details | Fig. 1 Molecular and phenotypic characterization of the plant overexpressing DL (Act1:DL). (A) RT-PCR analysis. Actin gene was used for a control. (B) Cross section of the shoot apex. The leaf primordia (P2–P4) and the shoot apical meristem (SAM) are indicated. Bars = 200 μm. |
For microarray analysis, RNA was isolated from the shoot apex that included the shoot apical meristem (SAM) and leaf primordia (including from P1 to P6). The shoot apices were dissected manually, and the dissected tissues were about one cubic-millimeter in size. Five shoot apices were used for a set of biological samples for RNA isolation. After conversion of mRNA to cDNA, RNA was transcribed with T7 RNA polymerase and labeled with the fluorescent dyes Cy3 or Cy5. Microarray experiments were carried out using rice 22K array (Agilent, Palo Alto). To assess the reproducibility of the signals, a pair of color swap experiment was repeated two times for two biological samples; in total, eight microarray data were obtained. Fig. 2 shows an example of the Scatchard plot of a pair of the color swap experiment. This pattern suggests that there is less effect on the color-labeling artifacts. Scatchard plots from the other sets of color swap experiments showed the pattern similar to that shown here. Microarray data are submitted to CIBEX (http://cibex.nig.ac.jp) with the accession number CBX59.
 View Details | Fig. 2 An example of Scatchard plot analysis comparing gene profiles in the shoot apices between the Act1:DL plant and the dl-sup1 mutant. |
Data obtained from microarray experiments were analyzed by the Spotfire DecisionSite 8.0 package (Spotfire, Somerville). To find significantly regulated genes, we used signals at the significant level of P<0.01 in t-test. These analyses identified 90 genes that showed at least two fold higher expression in the Act1:DL apices than in the dl-sup1 apices. By contrast, 250 genes were upregulated in the dl-sup1 mutant compared to the Act1:DL plant (Table 2). We counted the number of genes that are changed in the levels of the expression by in four classes; more than 2-, 3-, 5-, and 10-folds (Table 2). In any cases except for the case for more than the 10-fold change, the number of the genes that were upregulated in the Act1:DL plant was lower than those upregulated in the dl-sup1 mutant.
 View Details | Table 2 The number of genes upregulated or downregulated in the Act1:DL plant |
Cells in the leaf primordia differentiate along the central-lateral axes and different kinds of genes may be expressed depending on the cell types along this axis. The above results suggest that overexpression of DL may change the fate of all cells in the leaf primordia to that of cells in the central region in wild type, and may repress expression of genes specifically expressed in the cells in the lateral and peripheral regions. This repression, therefore, probably resulted in the reduction in the number of genes that are expressed in the Act1:DL plant.
Genes that changed their expression levels were categorized according to their functions (Fig. 3). In the genes downregulated in the Act1:DL plant, the genes encoding proteins related to transcription (mainly transcription factors) and those of enzymes increased in the number of genes and in the proportion to the total genes. Transcription factors and enzymes are characteristic to cell types and their function. Therefore, this result suggests that the shoot apex of Act1:DL has small number of cell types compared to that of dl-sup1 and supports the above inferences. In addition, whereas genes related to photosynthesis were upregulated in the dl-sup1 mutant, no such gene, except for one, is upregulated in the Act1:DL plant. This results suggests that cell fate toward photosynthetic cells is not specified in the leaf primordia if DL is constitutively expressed. Taking together these results, therefore, it is likely that overexpression of DL not only induces the genes related to midrib formation but also represses a number of genes in the cells other than those in the midrib region, resulting in cell specification restricted to midrib founder cells throughout the leaf primordia.
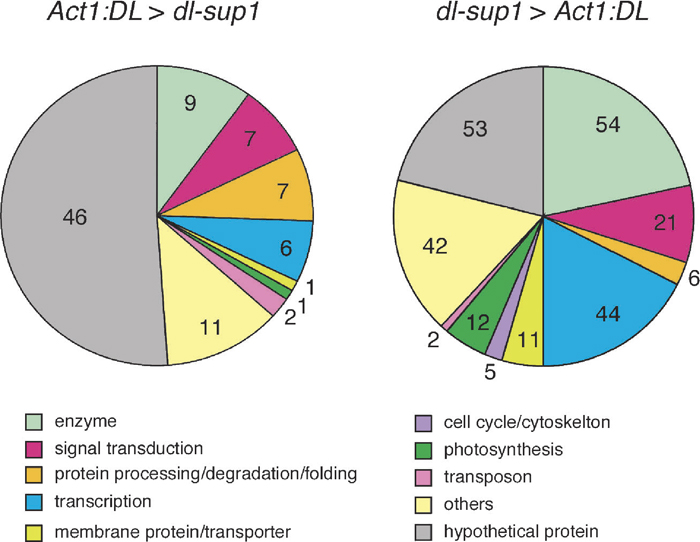 View Details | Fig. 3 The number of genes in various biological process categories. |
To get further insight for the genes that are upregulated in the Act1:DL plant, we chose arbitrarily 18 genes and examined expression patterns of them by RT-PCR analysis. First, we compared the expression levels of each gene in the shoot apex of three strains; Act1:DL, dl-sup1, and wild type. All genes examined were expressed higher in the Act1:DL plant than in the dl-sup1 mutant, suggesting that expression profiles from the microarray analysis are consistent with the expression levels of individual genes analyzed by RT-PCT. If expression of the chosen genes are enhanced by DL in the Act1:DL plant, the expression level of each gene obtained by the RT-PCR analysis is expected to be Act1:DL > wild-type (WT) > dl-sup1. The results obtained, however showed a few patterns in addition to the expected pattern (Fig. 4, Table 3). Therefore, we classified the genes into three classes according to their expression patterns; (A) Act1:DL > WT > dl-sup1 (expected), (B) Act1:DL > dl-sup1 > WT, (C) WT > Act1:DL > dl-sup1.
 View Details | Fig. 4 RT-PCR analysis of arbitrarily chosen 18 genes upregulated in the Act1:DL plant, compared to the dl-sup1 mutant. (1–3) Shoot apex of the Act1:DL plant (1), wild type (2), and the dl-sup1 mutant (3). (4, 5) Inflorescence of wild type (4) and the dl-sup1 mutant (5). (6) Leaf of wild-type seedling. (7) Root of wild-type seedling. The Actin1 gene was used for a control. |
 View Details | Table 3 Genes analyzed by RT-PCR in Fig. 3 |
The class A included genes encoding following proteins; galactose oxidase-like protein (Os01g0148500), O-methyltransferase (Os11g0303600), myo-inositol phosphate synthase (Os03g0192700), Myb-like transcription factor (Os04g0583900), Ankyrine-like protein (Os01g0188900), bZIP transcription factor (Os06g0662200), and OsEIL1 (Arabidopsis EIN3-related protein (Mao et al., 2006); Os03g0324200) (Table 3). Among these genes, the expression levels of the former four genes were considerably reduced in the dl-sup1 mutant and were moderately reduced in wild type (Fig. 4). This results strongly suggest that these four genes are under regulation of DL. It is likely that these four gene are involved in early process of midrib development.
Next, we examined the expression of the chosen genes in the inflorescence at the floral organ differentiation stage because DL regulates carpel development as well as midrib formation (Nagasawa et al., 2003; Yamaguchi et al., 2004). Among the genes in class A, the gene Os01g0148500 was expressed at the higher levels in wild-type flower than in the dl flower (Fig. 4). Thus, this gene showed similar expression pattern both in the shoot apex and in the developing flowers; higher expression in wild-type flower than in the dl flower. Therefore, the gene Os01g0148500 may be involved in both midrib formation and carpel specification under the control of DL. By contrast, expression of the genes Os11g0303600 and Os03g0324200 in the inflorescence was not observed in both wild type and the dl mutant, suggesting that these two genes may be involved in only midrib formation under the control of DL (Fig. 4). In the case of the genes Os03g0192700, Os04g0583900, Os01g0188900, and Os06g0662200, unlike in the shoot apex, expression patterns of these genes in the inflorescence seemed to be unrelated to DL function. This indicates that these four genes would be induced by genes other than DL in the inflorescence. All genes in class A were expressed in the leaf and root of the wild-type seedling, suggesting that these gene are involved in various aspects of biological activities besides midrib formation and carpel development.
The expression patterns of the genes in the class B and class C was unexpected. In the class B, the expression of each gene in wild type was lower than in the dl mutant, whereas the expression in wild type was higher than in the Act1:DL plant in the class C. To produce Act1:DL plant, DL began to be strongly expressed from undifferentiated cells in calli before regeneration and it continued to be expressed constitutively and strongly in the seedlings until the shoot apices were dissected to isolate RNA. It is likely that the constitutive and strong expression of DL throughout the life cycle of the Act1:DL plant may cause various artifacts to disturb normal gene expression. It is likely that, for example, the constitutive and strong expression of DL induced the genes in the class C, which are otherwise repressed in the wild-type shoot apex. Anyway, the genes in these classes should be excluded from further analyses because changes in the expression of these genes in the two classes may be artifacts by overexpression of DL.
In this paper, we have tried to identify the genes that are involved in midrib formation under the control of DL. We have found several genes that are expected to be regulated by DL. Although only a small number of the genes are examined here in detail, genes regulated by DL are divided into two groups. Genes in a group seem to be regulated by DL in a similar way in both the shoot apex and the inflorescences, whereas genes in the other group seem to be specifically regulated by in only the shoot apex. Thus, some genes regulated by DL may be shared by the mechanisms that regulate midrib formation and carpel development. On the other hand, other some genes may be involved in only midrib formation. If microarray analysis would be carried out using RNA from the inflorescences, genes specifically involved in carpel development might be identified.
Unexpectedly, strong and constitutive expression of DL throughout all stages may cause severe perturbation of normal gene expression in the Act1:DL plant. To identify the genes certainly regulated under the control of DL, the results from other approaches should be incorporated for analyzing the results obtained here. One possible way is the comparison of expression profiles in wild type and the dl mutant. Furthermore, inducible gene expression system such as DEX induction would be more effective to identify the genes regulated by DL either directly or indirectly. Combination of these approaches will identify key genes under the control of DL and elucidate the genetic networks regulating midrib formation and carpel development in rice.
We thank Dr. Y. Nagamura and Ms. R. Motoyama (National Institute of Agrobiological Sciences) for technical helps and advice in the microarray experiments, and Dr. M. Nakazono and Ms. K. Ohsawa (Univ. Tokyo) for technical assistance in making transgenic plants. This research was partly supported by a grant from the Ministry of Agriculture, Forestry and Fisheries of Japan (Integrated research project for plant, insect and animal using genome technology IP-1005) and by Grants-in-Aid for Scientific Research (17208002) from the Ministry of Education, Culture, Sports, Science and Technology.
|