| Edited by Kiichi Fukui. Masayoshi Shigyo: Corresponding author. E-mail: shigyo@yamaguchi-u.ac.jp |
Molecular markermaps are essential for the implementation of basic and applied research on genetics and breeding in edible Allium crops. Medium-density linkage maps exist separately for two economically important species, such as the bulb onion (A. cepa L.) and bunching onion (A. fistulosum L.), as shown in the review chapter prepared by McCallum (2007). Maps have been developed to date on the basis of RFLP (King et al., 1998; Martin et al., 2005) or AFLP (van Heusden et al., 2000a, 2000b) markers for the bulb onion and on the basis of AFLP (Ohara et al., 2005) and microsatellites (Tsukazaki et al., 2006) for the bunching onion. The two different bulb onion maps were subsequently anchored to the respective chromosomes via the use of the same complete set of A. fistulosum – A. cepa monosomic addition lines developed by Shigyo et al. (1996). Ensuing chromosomal assignments of bunching onion maps would be truly promising to launch an evolutionary study on the synteny between these two species.
Shigyo and co-workers demonstrated that chromosome manipulation techniques via the use of monosomic addition lines could be available to modify the endogenous chemical components of the bunching onion, such as flavonoids (Shigyo et al., 1997b; Masuzaki et al., 2006a, 2006b), carbohydrates (Hang et al., 2004a; Yaguchi et al., 2008b), chlorophyll (Dissanayake et al., 2008), or vitamin C (Yaguchi et al., 2008a). In most cases, alien chromosome addition lines induced the upregulation of these functional component productions. This finding might enhance the possibility of breeding for chemical components through the conventional chromosome manipulation technique.
We produced a population of shallot – bunching onion chromosome addition lines via the first and second backcrossings of their amphidiploid hybrids (♀) to shallot (♂) (Hang et al., 2004b). Forty-two hypo-allotriploids possessing 23 chromosomes, i.e., single-alien deletion plants (2n = 3x – 1 = 23, AAF-nF), were included in the BC1 progeny. On the other hand, eight monosomic addition plants (2n = 2x + 1 = 17, AA+nF) and 20 single-alien deletion plants were discovered in the BC2 progeny. In a previous study, the eight monosomic addition plants and the 62 single-alien deletion plants were analyzed by 6F chromosome-specific isozyme marker (Got-2) in order to identify its existence in their chromosome complements. The GOT isozyme analysis identified not only two monosomic addition plants possessing the 6F chromosome of A. fistulosum (AA+6F) but also two single-alien deletion plants missing 6F (AAF-6F). However, the remaining five monosomic addition plants and 60 single-alien deletion plants have not been identified.
The aims of the present study were to develop the A. fistulosum chromosome-specific genetic markers for identifying extra chromosomes in the remaining shallot – A. fistulosum monosomic addition plants and recognizing the deleted chromosomes in the single-alien deletion plants. Furthermore, the morphological and biochemical characteristics of the bulbs of single-alien deletion lines in dormancy were evaluated to determine the morphological effects of different numbers of extra chromosomes in an A. cepa diploid background and to reveal the availability for using those monosomic addition lines and single-alien deletion lines in place of existing shallot varieties.
The eight shallot – A. fistulosum monosomic addition plants (2n = 2x + 1 = 17, AA+nF) and 62 single-alien deletion plants (2n = 3x – 1 = 23, AAF-nF), including two AAF-6F plants developed previously (Hang et al., 2004b), were used to identify the genomic constitution of each plant. A. fistulosum ‘Kujo-hoso’ and A. cepa Aggregatum group ‘Chiang mai’ were also used as controls. In all the plant materials, somatic chromosomes of root tips were observed by Feulgen nuclear staining followed by squash method. Five root tips were collected from young roots of each plant. Pretreatment of root tips by 0.05% colchicine in 3 h at 20°C. Cells of root tips were fixed in the mixed solution of acetic acid and ethanol (in the ratio of 1 to 3, v/v). Cells were then hydrolyzed in 1N HCl in 6 min at 60°C and stained by leucobasic fuchcine. Treated root tips were placed onto glass slice, squashed in 45% acetic acid and observed under the microscope. After counting more than 15 cells per plant, chromosome numbers were recorded. The karyotype analyses were according to the standard nomenclature system for the chromosomes of Allium (Kalkman, 1984), which was generally agreed in the Eucarpia 4th Allium Symposium (De Vries, 1990). A. fistulosum chromosome-specific genetic markers of five isozymes [chromosome 1F, Lap-1F; 2F, Got-1F; 6F, Got-2F (Shigyo et al., 1994); 5F, Pgi-1F (Shigyo et al., 1995b); and 8F, Gdh-1F (Shigyo et al., 1995a)], four EST markers [3F and 6F, ACAHN07F; 4F, ACABE16F; 5F, ACAEJ67F (Kuhl et al., 2004; Table 1) and ACM071 (Tsukazaki et al., 2007)], twelve SSR markers [1F, AFS015 (Ohara et al., 2005) and AFA08G10 (Tsukazaki et al., 2008); 2F, AMS14 (Fischer and Backmann, 2000) and ACE020; 4F, AFA11H10 and AFAT13H10; 6F, AFA02H08 and AFRT08C02; 7F, AFA06A08 and AFA15E08; 8F, AFA11E12 (Tsukazaki et al., 2008) and AFS096 (Ohara et al., 2005)] and one 5S ribosomal DNA (7F, Af5SS) were available for the identification of eight kinds of extra chromosomes in the monosomic addition plants and single-alien deletion plants. Shibata and Hizume (2002) named the 5S rDNA units of two different sizes (about 350- and 530-bp long) in A. cepa Ac5SS and Ac5SL, respectively. The 5% (w/v) denaturing polyacrylamide gel can detect the slightly smaller PCR-fragment size of A. fistulosum than Ac5SS. We named a section of approximately 350 bp in A. fistulosum as Af5SS after its predecessor in A. cepa. Analyses for the four isozyme markers were carried out according to the procedures of Shigyo et al. (1994, 1995a). For DNA analyses, total genomic DNA was isolated from fresh leaf tissue using a miniprep DNA isolation method (van Heusden et al., 2000b). PCR amplifications of the four ESTs, twelve SSRs, and the 5S rDNA markers were performed according to the procedures of Kuhl et al. (2004), Tsukazaki et al. (2008), and Shigyo et al. (1996), respectively. PCR products were separated on 5% (w/v) denaturing polyacryl-amide gels according to the procedure of Martin et al. (2005).
 View Details | Table 1 PCR primer sets used in this study |
The chromosomal locations in A. fistulosum have been already determined for six genetic markers, but not for Got-2F (Shigyo et al., 1994) or Af5SS (Hizume, 1994). Of the eight, we assumed their chromosomal locations in A. fistulosum from a homologous chromosome of the shallot chromosome to which the shallot allele corresponding to each of the six markers had been assigned because there is a close genetic relationship between A. fistulosum and shallot. For example, the chromosomal location of Lap-1F was postulated as 1F chromosome based on the fact that Lap-1A, a shallot allele of Lap-1F, was allocated to 1A chromosome (Shigyo et al., 1995a), which is homologous to 1F. As occasion arises, a number of plants identified the genomic constitution were used for the following experiments.
The single-alien deletion lines identified with the A. fistulosum chromosome-specific genetic markers were applied for morphological evaluation. Three characteristics of the leaf, i.e., number of tillers, number of expanded leaves per tiller, and length of the leaf blade, of hypo-allotriploids (2n = 23) described by Hang et al. (2004b) were redivided into five types of single-alien deletion lines (AAF-1F, AAF-3F, AFF-4F, AAF-7F, and AAF-8F). In addition, bulb characterizations in six types of single-alien deletion lines (the above-mentioned five lines plus AAF-6F) and two types of monosomic addition lines (AA+6F and AA+8F) were recorded after harvesting. The shallot and an allotriploid between shallot and A. fistulosum (AAF) were also used as controls. The bulb characteristics were evaluated by measuring the maximum bulb diameter, bulbing ratio, and bulb color in June 2005, 10 months after planting in August 2004. The bulbing ratio was estimated by dividing the maximum bulb diameter by the minimum neck diameter.
Bulbs of AAF-1F, AAF-4F, AAF-8F, and AA+8F, along with those of shallot and AAF as controls, were utilized to gauge carbohydrate contents. Sugar extraction from the pieces of bulbs was performed as described previously (Hang et al., 2004b). A 0.5 ml aliquot of 70% EtOH extracts was vacuum-evaporated to dryness and redissolved in 0.5 ml of Milli-Q water. The extract was filtered by passing through a 0.45 μm syringe-type filter HCL-Disk3 (Kanto Chemical Co., Inc., Tokyo, Japan) and analyzed by high-performance anion-exchange chromatography (HPAEC) according to the procedure of Shiomi et al. (1997).
Bulbs of AAF-1F, AAF-4F, AAF-8F, and AA+8F, along with those of shallot and AAF as controls, were available for investigations of the components of ACSOs. Ten grams of bulb tissues was microwaved for 2 min until the tissues were completely cooked to denature the alliinase. The cooked tissues were ground and extracted with distilled water. The extract was centrifuged at 2,000 × g for 10 min at room temperature, and the supernatant was saved. The supernatant was centrifuged at 15,000 × g for 1 min and filtered by being passed through a 0.45 μm syringe-type HCL-Disk3 filter (Kanto Chemical Co., Inc., Tokyo, Japan). A 50 μl filtered sample was injected into a high-performance liquid chromatography (HPLC) system and quantified. The HPLC system included a pump, a degasser, a column oven, a diode array detector set to 220 nm, a data collection system (EZchrom EliteTM, Hitachi High-Technologies Corporation, Tokyo, Japan), and an AQUASIL SS-1251-120 column (4.6 mm i.d. × 250 mm in length, Senshu Scientific Co., Ltd., Japan). The solvent used was 0.1% trifluoroacetic acid and flowed for 15 min at a flow rate of 0.6 ml/min. Standard compounds of S-methyl (Me)-, S-2-propenyl (allyl, Al)-, and S-1-propenyl (Pe)-CSOs were synthesized at the Somatech Center (House Foods Corporation, Japan). A series of standards was dissolved in Milli-Q water and analyzed as described above.
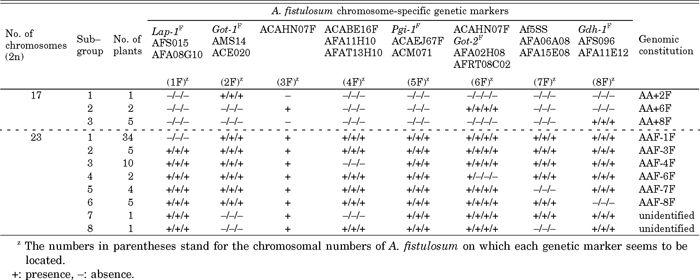
The somatic metaphase chromosomes of 70 plants were counted and shown in Table 2. Segregating isozyme and DNA markers could be used to detect the presence or absence of A. fistulosum chromosomes (1F–8F) under the diploid background of shallot. Fig. 1 and Fig. 2 display photographs of polyacrylamide gels stained for those isozyme and DNA markers with allelic segregation.
 View Details | Table 2 Segregation patterns of A. fistulosum-specific genetic markers in shallot – A. fistulosum monosomic addition lines (2n = 2x + 1 = 17, AA+nF) and the single-alien deletion lines (2n = 3x - 1 = 23, AAF-nF) |
 View Details | Fig. 1 Glutamate dehydrogenase (GDH) zymograms. From left to right lanes: A. fistulosum (FF), A. cepa Aggregatum group (AA), amphidiloid hybrid (AAFF), triploid hybrid (AAF), monosomic addition lines (I and II types) and single-alien deletion lines (III and IV types). (I) AA+8F; (II) AA+6F; (III) AAF-1F, AAF-4F, AAF-6F, AAF-7F; (IV) AAF-8F. The numeral data in parentheses indicate the numbers of plants obtained. |
 View Details | Fig. 2 Representative amplification profiles for A. fistulosum chromosome-specific DNA markers in A. fistulosum (FF), shallot (AA), two shallot – A. fistulosum monosomic addition lines (1 and 2) and four shallot – A. fistulosum single-alien deletion lines (3, 4, 5, and 6) on a denaturing polyacrylamide gel after silver staining. M, molecular size marker (100 bp DNA Ladder). Arrows indicate the chromosome-specific markers; a, ACAHN07F for chromosomes 3F and 6F; b, ACABE16F for 4F; c, ACAEJ67 for 5F; d, Af5SS for 7F. |
A. fistulosum and shallot showed multiple band patterns on GDH zymograms (Fig. 1). The specific activities of the bands declined gradually from cathode to anode. The band ladders of the two species did not stay in position relative to each other. The allopolyploids (AAFF and AAF) formed broad and blotted bands that migrated to the intermediate positions between the parental bands. However, the band ladders of AAF shifted slightly to the anodal side. Previous isozyme studies (Shigyo et al., 1995a, 1995c) demonstrated that the expression of GDH multiple band patterns in these two species was controlled by two different alleles (Gdh-1A, Gdh-1F) at the single locus Gdh-1 and that the allele Gdh-1A was located on shallot 8A chromosome. From the point of view of the close genetic relationship between A. fistulosum and shallot, there is a high probability that the gene locus Gdh-1 is located on the homoeologous chromosomes 8A and 8F. In this study, the allele Gdh-1F was, therefore, regarded as a chromosome marker for 8F. Furthermore, it was clarified that the dosage effect of the GDH genes appeared in the mobility of the band ladders.
In the eight monosomic addition plants and 62 single-alien deletion plants, there were two types of band profiles on GDH zymograms. Profile I of the monosomic addition line and III of the single-alien deletion line were identical to that of the allotriploid AAF regarding the patterns and positions of GDH bands, as shown in Fig. 1. Profile II of the monosomic addition line and IV of the single-alien deletion line were identical to those of shallot. These results indicate that five of monosomic addition plants and 57 of single-alien deletion plants possess a genotype (Gdh-1A/Gdh-1A/Gdh-1F) and three of monosomic addition plants and five of single-alien deletion plants possess a genotype (Gdh-1A/Gdh-1A). It had been revealed that two SSR markers (AFS096 and AFA11E12) were allocated on the 8F chromosome of A. fistulosum (Tsukazaki et al., 2008). These SSR marker analyses showed that five of monosomic addition plants possessed the 8F chromosome and five of single-alien deletion plants lacked the 8F chromosome. Consequently, five of the monosomic addition plants could be identified as AA+8F and five of the single-alien deletion plants as AAF-8F (Table 2).
In a previous study, Shigyo et al. (1994) suggested that the two gene loci for dimeric GOT isozymes, Got-1 and Got-2, could be regarded as respective chromosome markers for 2F and 6F chromosomes. Hence, the allele Got-1F was employed as a genetic marker for 2F, and Got-2F, as a genetic marker for 6F. The results of GOT isozyme analysis are summarized in Table 2. One of monosomic addition plants and 60 of single-alien deletion plants possess a genotype (Got-1A/Got-1A/Got-1F) and seven of monosomic addition plants and two of single-alien deletion plants possess a genotype (Got-1A/Got-1A). Two of monosomic addition plants and 60 of single-alien deletion plants possess a genotype (Got-2A/Got-2A/Got-2F) and six of monosomic addition plants and two of single-alien deletion plants possess a genotype (Got-2A/Got-2A). Tsukazaki et al. (2008) revealed that four SSR markers AMS14, ACE020, AFA022H08, and AFRT08C02 were allocated on the 2F, 2F, 6F, and 6F chromosome of A. fistulosum, respectively (Table 2). Consequently, three out of eight monosomic addition plants were identified as AA+2F and AA+6F, and two out of 62 single-alien deletion plants, as AAF-6F. The two of single-alien deletion plants which did not have a 2F chromosome were not recognized as AAF-2F, because these two plants showed the lack of 4F or 7F chromosomes of A. fistulosum (as described below). Unfortunately, two of single-alien deletion plants belonged in the sub-group 7 and 8 showed disagreement between cytogenetics and molecular data.
A previous study (Shigyo et al., 1995a) revealed that monomeric LAP isozymes are controlled by two different alleles (Lap-1A, Lap-1F) at the single locus Lap-1 in A. fistulosum and shallot and that the allele Lap-1A is located on shallot 1A chromosome. The allele Lap-1F of A. fistulosum is regarded as a chromosome marker for 1F for the same reason as GDH. Furthermore, two SSR markers AFS015 (Ohara et al., 2005) and AFA08G10 (Tsukazaki et al., 2008) analyses revealed that Lap-1F negative plants also showed the negative patterns of the SSR profile. Consequently, 34 of 62 single-alien deletion plants are identified as AAF-1F (Table 2).
On 5% denaturing polyacrylamide gels, the three EST primer sets evaluated in this study (ACAHN07F, ACABE16F, and ACAEJ67F) showed clear banding differences between A. fistulosum and shallot (Fig. 2a–2c). Each primer set produces a single amplification product in both of these parental species, but the results clearly show that there were slight mobility differences. This co-dominant expression of bands seems to be regarded as allelic variation. A preliminary test with a complete set of A. fistulosum – shallot monosomic addition lines revealed that the EST markers ACABE16A and ACAEJ67A of shallot are located, respectively, on 4A and 5A chromosomes. Therefore, the EST markers ACABE16F and ACAEJ67F of A. fistulosum were used as chromosome markers for 4F and 5F, respectively. Previous research revealed that the EST markers AFA11H10, AFAT13H10, and ACM071 and the PGI isozyme markers were allocated on 4F, 4F, 5F (Tsukazaki et al., 2008) and 5F (Shigyo et al., 1995b) chromosome, respectively. Three 4F chromosome specific-markers were not detected in 11 of single-alien deletion plants in this study. Only one of these 11 single-alien deletion plants also showed the lack of 2F chromosome, therefore, this plant was classified into sub-group 7. Consequently, 10 of 62 single-alien deletion plants were identified as AAF-4F and no plant was identified as AAF-5F (Table 2).
Shibata and Hizume (2002) reported that A. cepa possesses 5S rDNA units of two different sizes with lengths of about 350 and 530 bp. According to them, A. cepa has two 5S rDNA loci in the proximal and distal regions (corresponding to the small and large units, respectively) of the short arm of chromosome 7. In this study, an interspecific polymorphism between A. fistulosum and shallot was detected around small subunits on the 5% denaturing polyacrylamide gel. Namely, the PCR-fragment size of A. fistulosum was slightly smaller than that of shallot (Fig. 2d). A section of about 350 bp of A. fistulosum was named as the Af5SS unit after its predecessor in A. cepa [Ac5SS in Shibata and Hizume (2002)]. Hizume (1994) also revealed that this unit was localized on the proximal region of the short arm of 7F chromosome in A. fistulosum. Based on the occurrence of the PCR product derived from the Af5SS unit, the presence or absence of 7F was recognized in the monosomic addition lines and the single-alien deletion lines (Table 2). Two SSR markers AFA06A08 and AFA15E08 were allocated on the 7F chromosome by Tsukazaki et al. (2008). These markers revealed that four single-alien deletion plants lacked the 7F chromosome. In brief, four of 62 single-alien deletion plants were identified as AAF-7F. One of 62 single-alien deletion plants which was the lack of 2F chromosome also showed the lack of 7F chromosome, therefore, was classified into the sub-group 8 (Table 2).
The preliminary test revealed that the EST marker ACAHN07A is located on both 3A and 6A chromosomes. Therefore, the four-step approach was conducted to identify the AAF-3F. First, all the single-alien deletion plants were analyzed by 1F, 4F, 7F, and 8F chromosomes specific-markers mentioned above. Seven plants were not identified the chromosome composition in this study. Second, the EST marker ACAHN07A was analyzed in those seven plants. Seven plants showed the positive pattern of ACAHN07A marker in this study. Third, all plants of ACAHN07A positive plants were applied to GOT isozyme and two SSR markers (AFA02H08 and AFRT08C02) analyses. If the negative pattern of these three markers of the 6F chromosome were detected in any plant, those plants can be identified as AAF-6F. Two of seven plants with the ACAHN07A positive pattern were eliminated as AAF-6F from the following step in this study. Finally, the all plants with positive pattern of 6F chromosome specific-markers were analyzed by 5F chromosome specific-markers mentioned above. This operation could separate AAF-3F and AAF-5F. In the present study, fortunately, five of 62 single-alien deletion plants were identified as AAF-3F (Table 2).
Eight monosomic addition plants consisted of one AA+2F, two AA+6F, and five AA+8F lines. Sixty of the 62 single-alien deletion plants could be identified as six different single-alien deletion lines (AAF-1F, AAF-3F, AAF-4F, AAF-6F, AAF-7F, and AAF-8F) out of the eight possible types. The frequencies of single-alien deletion lines found in this study are shown in Table 2. The somatic metaphase chromosomes of six types are shown in Fig. 3. There was a tendency for the numbers of plants of AAF-1F and AAF-4F to be much larger than those of other deletion types. Conversely, no plant has, thus far, been found for AAF-2F and AAF-5F. More sufficient numbers of single-alien deletion plants should be developed to identify these two single-alien deletion lines.
 View Details | Fig. 3 Somatic metaphase chromosomes of shallot (AA), A. fistulosum (FF), shallot – A. fistulosum monosomic addition lines (AA+2F, AA+6F, and AA+8A), allotriploid between shallot and A. fistulosum (AAF), and single-alien deletion lines (AAF-1F, AAF-3F, AAF-4F, AAF-6F, AAF-7F, and AAF-8F). Scale bar = 10 μm. |
The single-alien deletion plants which showed the vigorous growth were morphologically characterized, and the results are summarized in Table 3 and Table 4. Plants of AAF-6F were generated before the other type of single-alien deletion lines; therefore, the data in the seedling stage was not used for comparison. Five types of single-alien deletion lines showed differences from each other in the number of expanded leaves per tiller and length of the leaf blade but not in the number of tillers (Table 3). Seedlings of AAF-8F had many more expanded leaves (6 leaves per tiller) than the other type of single-alien deletion lines. Regarding leaf blade length, seedlings of AAF-7F grew more vigorously, as expressed in its long leaf blade (217.3 mm). All six single-alien deletion lines and two monosomic addition lines conformed to shallot regarding the habit of bulb formation, but they differed from each other and from the parents, the shallot and AAF, in several bulb characteristics (Table 4 and Fig. 4). The single-alien deletion line AAF-7F had the largest diameter of the six single-alien deletion lines and was bigger than that of the shallot and AAF, while AAF-4F had the smallest diameter in the six single-alien deletion lines. The bulb diameters of two types of monosomic addition lines AA+6F and AA+8F were smaller than that of the shallot and AAF. Four types of single-alien deletion liens (AAF-1F, AAF-3F, AAF-4F, and AAF-6F) and two types of monosomic addition lines (AA+6F and AA+8F) exhibited reddish-purple skin color and a purple outer scale, as did the shallot and AAF bulbs. On the other hand, yellow skin and light-purple outer scale were observed in the bulbs of AAF-7F. The colors of the bulb skin and outer scale of AAF-8F were also reddish-yellow and pink, respectively.
 View Details | Table 3 Leafing characteristics of five types of shallot – A. fistulosum single-alien deletion lines and allotriploids (AAF) |
 View Details | Table 4 Bulb characteristics of six types of shallot – A. fistulosum single-alien deletion lines, two types of monosomic addition lines, shallot (AA), and allotriploids (AAF) |
 View Details | Fig. 4 Bulb postures of shallot (AA), allotriploid between shallot and A. fistulosum (AAF), shallot – A. fistulosum single-alien deletion lines (AAF-1F, AAF-3F, AAF-4F, AAF-6F, AAF-7F, and AAF-8F), and shallot – A. fistulosum monosomic addition lines (AA+6F and AA+8F). Scale bar = 20 mm. |
Biochemical analyses were conducted three single-alien deletion lines (AAF-1F, AAF-4F, and AAF-8F) and one monosomic addition line AA+8F which obtained the number of lines with five or more for the analyses of bulb characteristics (Table 4). There was a significant difference in the total sugar content between shallot [73.9 mg/g fresh weight (FW)] and shallot carrying A. fistulosum chromosomes, i.e., three types of single-alien deletion lines (AAF-1F, AAF-4F, and AAF-8F), the monosomic addition line AA+8F, and AAF, in which sugars over 200 mg/g FW were detected (Fig. 5a). There were significant differences in the fructan content with the degree of polymerization (DP) 3 or higher between shallot and shallot carrying A. fistulosum chromosomes, i.e., AAF and AAF-1F. While there were no significant difference in the mono- and di-saccharides content between shallot and AAF, single-alien deletion lines, and AA+8F. Moreover, shallot could not produce fructan with DP 12 or more, although the single-alien deletion lines, the monosomic addition line AA+8F, and AAF produced fructan with DP 20 or more, especially AA+8F, which had the longest chains (Fig. 6).
 View Details | Fig. 5 Content of sugars (a) and S-alk(en)yl-L-cysteine sulfoxide (ACSO; b) in the bulb of shallot (AA), allotriploid between shallot and A. fistulosum (AAF), shallot – A. fistulosum single-alien deletion lines (AAF-1F, AAF-4F, and AAF-8F), and shallot – A. fistulosum monosomic addition line (AA+8F) in August 2005. Figures followed by the same letter in each plant materials are not significantly different in total sugars or total ACSO content at P < 0.05 according to Tukey’s multiple range test. AlCSO, S-2-propenyl (allyl) CSO; PeCOS, S-1-propenyl CSOs; MeCSO, S-methyl CSO. |
 View Details | Fig. 6 Chain-length distributions of fructan in basal leaf sheaths of shallot (AA), allotriploid between shallot and A. fistulosum (AAF), shallot –A. fistulosum single-alien deletion lines (AAF-1F, AAF-4F, and AAF-8F), and shallot - A. fistulosum monosomic addition line (AA+8F). |
The ACSOs were separated with baseline resolution. MeCSO was separated first (retention time, 6.3 min), followed by AlCSO (7.4 min) and PeCSO (8.5 min). There was a great difference in the total ACSO contents between shallot and AAF, the three types of single-alien deletion lines (AAF-1F, AAF-4F, and AAF-8F), and the monosomic addition line AA+8F (Fig. 5b). The contents of PeCSO, the primary ACSO of A. cepa and A. fistulosum, were almost identical in all the examined plants. On the other hand, the shallot showed a significant increase in the contents of AlCSO, the principal ACSO of garlic (A. sativum), compared with each single-alien deletion lines. In addition, MeCSO, the major ACSO of Chinese chives (A. tuberosum) and rakkyo (A. chinense), had a content in shallots that was two to four times as high as that in each single-alien deletion lines, AA+8F, and AAF.
The present study has freshly identified three and six types of shallot – A. fistulosum monosomic addition lines and single-alien deletion lines, respectively. Six types of single-alien deletion lines, including AAF-6F described in our previous study (Hang et al., 2004b), have so far been obtained (Fig. 3). The two other types of single-alien deletion lines, in which either 2F and 5F chromosomes was absent, could not be produced. To facilitate the mapping and chromosomal assignment of genes in A. fistulosum, it is necessary that a complete set of the single-alien deletion lines is developed, i.e., production of AAF-2F or AAF-5F. However, six types of single-alien deletion lines and several monosomic addition lines, including AA+2F, could allocate the gene loci to the single chromosome, expect for the gene loci located on the both 2F and 5F chromosomes. Actually, fifteen linkage groups based on short sequence repeats, cleaved amplified polymorphic sequences, and insertion-deletion markers of A. fistulosum have been allocated to a single chromosome via the use of this chromosome addition set (Tsukazaki et al., 2008). They have started to integrate the A. fistulosum linkage map with the A. cepa map developed by Martin et al. (2005).
The frequency of homoeologous chromosome pairing and recombination is quite high in the meiosis of F1 hybrids between these two species (Emsweller and Jones, 1935; Maeda, 1937; Levan, 1941; Cochran, 1950; Tashiro, 1984; Peffley, 1986). In the present study, the two of single-alien deletion plants which belonged in the sub-group 7 and 8 showed disagreement between cytogenetics and molecular data. These plants perhaps resulted from the chromosomal substitution and recombination. However, this has yet to be confirmed by the fluorescent genomic in situ hybridisation (GISH) analysis. GISH has been shown to be valuable for discriminating between closely related genomes (Anamthawat-Jónsson et al., 1990) and for identifying alien chromosomes and chromosome segments (Schwarzacher et al., 1992). Shigyo et al. (1998) showed no clear exchanges of chromosome segments between A. cepa and A. fistulosum in the series of A. fistulosum – shallot monosomic addition lines by means of GISH analyses. Barthes and Ricroch (2001) revealed that four of 17 A. fistulosum – A. cepa monosomic addition plants possessed an A. fistulosum chromosome carrying a labelled signal indicative of chromosomal structural rearrangements involving the transfer of A. cepa chromatin onto an A. fistulosum chromosome by means of GISH analyses. GISH technique can fail to identify very short recombinant segments. In such cases, the use of chromosome-specific markers along the chromosome is required to confirm the identity of the monosomic addition lines and the single-alien deletion lines. Two single-alien deletion plants of the sub-group 7 and 8 in this study have been screened using the multiple molecular markers, therefore, the additive GISH analyses should reveal the recombination events of these plants.
From the point of view of a close genetic relationship between A. fistulosum and shallot, there is a high probability that a number of orthologues are located on a same group of chromosomes, namely homoeologous chromosomes, in these two species. As one case, AAF-8F showed the largest number of expanded leaves in the single-alien deletion lines and the controls (Table 3). A. fistulosum carrying 8A chromosome as an extra chromosome (FF+8A) showed slow expansion of leaf in the morphological characterization of a complete set of A. fistulosum – shallot monosomic addition lines (Shigyo et al., 1997a). These results indicated that anonymous factors related to inhibit the leaf expansion would be located on the 8F chromosome. Moreover, the line had yellow skin and a light-purple outer scale, differently from the reddish-purple skin and purple outer scale of the shallot, AAF, and other single-alien deletion lines (Table 4). The bulb of AAF-8F also turned reddish-yellow and pink in the skin and outer scale, respectively. The flavonoid 3’-hydroxylase (F3’H) gene to synthesize quercetin, which is a major flavonoid compound in the colored bulb of A. cepa, and the dihydroflavonol 4-reductase (DFR) gene to change a yellow bulb into a red one (Kim et al., 2004) have been assigned to 7A chromosome (Masuzaki et al., 2006a, 2006b). The F3’H gene, which would be located on 7F chromosome, cannot influence the bulb color of shallot because the gene seems to have a low expression level (Masuzaki et al., 2006b). Therefore, we hypothesize that the light bulb color of AAF-7F depends on the absence of the DFR gene of A. fistulosum; the gene, unlike the F3’H gene, seems to be functional. As chromosome 8F might have no flavonoid biosynthetic genes encoding from chalcone synthase to anthocyanidin synthase, as seen from the results of chromosomal assignment of the genes in shallot (Masuzaki et al., 2006a, 2006b), the light bulb color observed in AAF-8F was confounding. The reddish-purple skins of shallot complexify the bulb pigmentation of the single-alien deletion lines AAF-7F and AAF-8F. An anonymous factor related to pigmentation of shallot bulb other than the enzyme genes described above might be located on the chromosomes. Further analyses of the enzyme activity and gene expression related to pigmentation of shallot bulb in the single-alien deletion lines should reveal the pigmentation system of shallot.
AAF, the three types of single-alien deletion lines, AAF-1F, AAF-4F, and AAF-8F, and a monosomic addition line AA+8F differed widely from shallot regarding their components of sugars and ACSOs (Fig. 5). Regarding sugars, AAF, AA+8F, and the single-alien deletion lines carrying chromosomes of A. fistulosum showed higher contents of fructans, which are oligosaccharides with chain lengths higher than DP 2, than shallot (Fig. 5a), and the chain lengths in shallot were the shortest (Fig. 6). These results indicated that the chromosomes derived from A. fistulosum in the diploid background of shallot may contribute to an increase in the fructan production in shallot bulbs. The previous study revealed that the important quantitative trait locus (QTL) Frc affecting fructan content and the major enzyme gene sucrose phosphate synthase (SPS) related sucrose synthesis were allocated on chromosome 8 of A. cepa (McCallum et al., 2006; Yaguchi et al., 2008b). AA+8F showed higher fructan content than shallot, indicating that anonymous factors related to produce the sugars would be located on the 8F chromosome. Fructan biosynthesis is initiated by the enzyme sucrose:sucrose 1-fructosyltransferase (1-SST), which catalyses the formation of 1- kestose from sucrose (Vijn et al., 1998). It has been reported that 1-SST was induced by high sucrose contents in barley leaves (Muller et al., 2000; Wang et al., 2000). These indicate that the enzyme genes which catalyze sucrose biosynthesis are also important for fructan productions. The interpretation of biochemical and morphological phenotypes of alien monosomic addition lines and single-alien deletion lines is challenging since genes on the alien chromosome are expressed in the diploid genetic background of a divergent parent (Chang and de Jong, 2005). Support for the idea that heterozygosity or polyploidy in sucrose metabolism genes such as SPS and sucrose synthase (SuSy) can induce marked changes in sugar metabolism is provided by studies in maize. Causse et al. (1995) observed significant heterosis for SPS activity in maize hybrids and subsequently reported co-location of the QTL for SPS activity with the structural gene (Prioul et al., 1999). More recent studies of gene expression in diploid (Auger et al., 2005) and triploid (Swanson-Wagner et al., 2006) maize hybrids have also revealed non-additive expression of SPS and SuSy. The additive experiment of the enzyme activities related to fructan and sucrose biosyntheses in the shallot, the single-alien deletion lines, and the monosomic addition lines should reveal the gene expression event on the alien chromosome of A. fistulosum in shallot.
In ACSOs, Yoo and Pike (1998) reported that the total ACSO content of A. fistulosum was lower than that of shallot. AAF and the single-alien deletion lines showed much lower contents of total ACSO than shallot in the present study (Fig. 5b). This result suggested that chromosomes derived from A. fistulosum in the diploid background of shallot might carry anonymous factors to inhibit the synthesis and/or to promote the degradation of ACSOs in the bulb of shallot. Since 1 mol of pyruvic acid is produced for each mole of ACSO hydrolyzed by alliinase, enzymatically produced pyruvic acid has long been used as a proxy for measuring pungency (Schwimmer and Weston, 1961). Crowther et al. (2005) showed a linear relationship between the content of ACSOs and pyruvic acid. In addition, they showed a negative correlation between the sweetness and pyruvic acid content. These results indicated that the AAF and several types of single-alien deletion lines could be a mildly pungent, sweet variety of the shallot. The similar bulb size between each single-alien deletion lines and shallot might promote the conversion of using shallot into single-alien deletion lines.
|