| Edited by Yoshibumi Komeda. Bhavanath Jha: Corresponding author. E-mail: bjha@csmcri.org |
Abiotic stress is the primary cause of crop loss world wide, reducing average yields of major crop plants by more than 50% (Bray et al., 2000; Vinocur and Altman, 2005). The problem is huge, almost 1000 million ha of land is affected by soil salinity (Szabolcs, 1994), 7% of total land area. Of the 1.5 billion ha cultivated land, about 5% (77 million ha) is affected by salt (Munns et al., 1999). Plants growing in a particular habitat are often well adapted to the environmental stress conditions at that place. Understanding of the molecular processes regulating these metabolic adaptations will facilitate engineering of abiotic stress tolerance. The molecular mechanism of salt adaptation in halophytes is far improved compared to glycophytes. Salicornia brachiata is a highly salt tolerant plant than most other genera of halophytes. It is a leafless annual small bushy plant that grows up to 50 cm in height and has tiny flowers within green jointed succulent stem that forms seed-bearing spikes. S. brachiata can grow in wide range of salt concentrations, accumulating 40–50% salt of its dry weight. The vegetable salt from Salicornia is known as Saloni, contains 5–7% KCl along with NaCl, making it suitable for the high blood pressure patient. Calcium, magnesium and iodine micronutrients are naturally present in the Salicornia salt (Ghosh et al., 2005; US patent no. 6929809).
Expressed sequence tags (ESTs) analysis is an effective method, for getting information of gene expression and also provides an opportunity to identify new genes involved in biological functions. Various researchers have recently reported genes and ESTs, induced by cold, drought, high salinity stress and ABA in many plant species (Amtmann et al., 2005; Vinocur and Altman, 2005; Xu et al., 2007). There are few reports on ESTs generation and analysis in response to salt treatment on different halophytes like Suaeda salsa (Zhang et al., 2001), Tamarix androssowii (Wang et al., 2006), Thellungiella halophila (Wang et al., 2004), Mesembryanthemum crystallinum (Kore-eda et al., 2004) and Avicennia marina (Mehta et al., 2005).
The present study reports identification of genes in response to salinity in the extreme halophyte plant S. brachiata. The ESTs are analyzed and classified according to their putative functions. The expression of 90 unknown and hypothetical genes was studied by reverse Northern analysis. Transcript analysis of ASR1, PM-H+ATPase revealed its expression with salt stress.
Salicornia seeds were germinated in plastic pots containing garden soil. One set of two-month-old seedlings was exposed to 500 mM NaCl stress treatment for 24 h whereas a duplicate set of seedlings were maintained under control conditions. Total RNA was isolated from shoot material using Guanidium-isothiocyanate method. The cDNA from both control and NaCl treated plants were prepared using manufacture’s protocol (Invitrogen, USA). Subtractive hybridization was carried out to normalize the sequence abundance and to enrich the differentially expressed clones involved in salinity stress according to the Mishra et al. (2005). The 5’ end of each cDNA was ligated with an adaptor containing EcoRI site. cDNA was ligated unidirectionally into EcoRI and XhoI site of the phage lambda Zap expression vector according to the manufacturer’s instruction using a Uni-Zap-XR cDNA library construction kit (Stratagene, LaJolla, California, USA). Individual recombinant plaques from the primary cDNA library were manually transferred into 96 well format flat bottom storage plates containing 200 μl SM buffer (100 mM NaCl, 8 mM MgSO4.7H2O, 50 mM Tris-HCl pH 7.0, 0.04% gelatin) supplemented with 5% DMSO and 0.5% chloroform. The cDNA inserts from each recombinant phage were PCR amplified, with 150 ng of universal M13 forward and reverse primers (5’-GTAAAACGACGGCCAGT-3’ and 5’-CAGGAAACAGCTATGAC-3’), using 1 μl of recombinant phage suspension as a template in 100 μl reaction volume for 30 cycles with the following PCR conditions, 94°C for 1 min, 55°C for 1 min and 72°C for 2 min. PCR amplified cDNA inserts were purified using Strataprep 96 PCR purification kit (Stratagene) to remove unincorporated primers and nucleotides. The size and quality of cDNA inserts were monitored on agarose gels. All the PCR amplified products having a 200 bp or larger cDNA inserts were re-arrayed into a new 96-well PCR plate. One micro litre of purified PCR amplified cDNA inserts were directly used as template for cycle sequencing using DYEnamicTM ET terminator cycle sequencing kit (Amersham Biosciences) with T3 sequencing primers according to the manufacturers instructions and sequencing was performed on mega base sequencer. The raw sequencing data was processed to remove the poor quality sequences using PhredPhrap (Ewing and Green, 1998) software and vector and adaptor sequences by cross_match software. BLASTX 2.2.19+ programme of NCBI (Altschul et al., 1997) is used for searching sequence similarity between processed cDNA sequence outputs and non-redundant amino acid public database. The BLASTX hit lower than E value = 1e-2 was considered for the functional annotation of the ESTs sequence.
To study the expression of ASR1 (EB484610) and PM-H+ATPase (EB484660) gene, two-month old seedlings were treated with different concentration of NaCl ranging from 0.01 M to 2 M for 24 h. In another experiment the plants were treated with 0.250 M NaCl for different time periods like 2, 4, 8, 12, 18, 24, 36, 48 and 72 h to study the time course transcript expression. Total RNA was isolated from the treated plants (shoot) using Guanidium – isothiocyanate method. Approximately 20 μg of total RNA was resolved on 1% formaldehyde agarose gel and transferred to positively charged Hybond N membrane. The blots were probed with [α32P]-labelled (nick translated) partial length of ASR1 and PM-H+ATPase ESTs. Hybridization and post hybridization washes were carried out as reported earlier (Reddy et al., 1998). The blots were autoradiographed. The signal intensity of RNA bands was quantified using Gene analysis tool software (Cambridge, U.K.), and plotted as relative value with respect to ribosomal RNA band as control for each treatment separately.
The selected ninety clones were PCR amplified (94°C for 1 min, 55°C for 1 min and 72°C for 2 min) using 1 μl phage stock of the hypothetical and unknown genes obtained during the subtractive hybridization. The concentration of the amplified product was determined using the Nanodrop spectrophotometer (ND1000, USA). Equal volume (2 μg) of each amplified product was electrophoresed on two different 1% agarose gels (0.5x TBE) simultaneously. The DNA from both the gels were transferred to positively charged Hybond N membrane and immobilized by UV crosslinking (U.V. 500 Crosslinker, Amersham Bioscience, USA). Total RNA was isolated from control and 500 mM NaCl treated (24 h) plants using Guanidium-isothiocyanate method. mRNA was isolated using PolyATract® mRNA Isolation Systems (Promega, USA). Two microgram of each mRNA from both control and salt treated plants were used for cDNA synthesis in the presence of [α32P] dCTP. One set of blots were hybridized with control cDNA probe and other with salt treated cDNA probe. Hybridization and post hybridization were carried out as mentioned earlier (Reddy et al., 1998). The blots were autoradiographed and the bands were compared to analyze up-regulated and down- regulated genes.
A total of 1200 clones were picked up and analyzed by the PCR. The PCR reaction with M13 forward and reverse primers confirmed 1002 clones to be recombinant. Poor quality sequences and the 5’ adaptor sequences if found were removed. Out of the 1002 sequences, 930 sequences were submitted to the NCBI database (GenBank accession number EB484528 to EB485289 and EC906125 to EC906292) while the remaining poor quality sequences, were discarded.
Out of 930 sequences 789 ESTs showed similarity with known or unknown category of genes in NCBI database. These ESTs were grouped into 13 functional categories including defense and cell rescue (4.8%), protein metabolism and sorting (3.0%), membrane transporters (3.8%), gene expression and RNA metabolism (11.2%), photosynthesis (10.9%), signaling component (4.3%), metabolism enzymes (14.8%), cell wall structure (2.5%), cell division, cycle and growth (3.8%), cytoskeleton (0.9%), energy (1.14%), miscellaneous (8.9%) and unknown or hypothetical (29.8%) (Fig. 1). Some of the genes in high abundance like ribosomal protein, Rubisco small subunit, metallothionein, Chl a/b binding are given in Table 1. Approximately 29% of ESTs did not exhibit homology with known genes, thus were classified in unknown category. In total population, 4.8% sequences showed similarity with salt stress related genes. The stress related genes are grouped into six different classes on the basis of their major function like general stress proteins, signaling components, reactive oxygen species scavengers, transmembrane transporters and ion homeostasis and transcriptional regulators (Table 2).
 View Details | Fig. 1 Functional classification of ESTs of S. brachiata, values against each classification are given in percentage. |
 View Details | Table 1 Most abundant mRNA |
 View Details | Table 2 Abundance profile of genes related to salt tolerance |
In our study, after 4 h of NaCl treatment, plants started to show signs of wilting but got resurrected after 24 h. The resurrection shows that the normal metabolism gets resumed in halophytic environment reflecting the unique property of halotolerant plants. The representation of high number of ribosomal proteins, Rubisco small subunit and Chl a/b binding genes indicate that plants remained healthy even in the presence of NaCl. Similarly a high number of these genes were also procured in ESTs analysis of other halotolerant plants like Suaeda salsa (Zhang et al., 2001), Tamarix androssowii (Wang et al., 2006), Thellungiella halophila (Wang et al., 2004) and Mesembryanthemum crystallinum (Kore-eda et al., 2004).
Whether halophytes have a distinct signaling pathway and regulatory network to impart the salt stress tolerance is not clear. Environmental signals are first perceived by specific receptors, which modulate expression and activation of different genes leading to stress tolerance. Many salt related genes involved in abiotic stress signaling pathways are found in S. brachiata ESTs (Table 3). Calmodulin, Ca2+ binding proteins and receptor like serine threonine kinase known in the literature as signaling components involved in various signal cascades, are present in the Salicornia ESTs. Serine threonine kinase is an important signaling molecule that transmits the signal to down stream transcriptional regulators and other stress related genes. Similarly, calmodulin and serine threonine kinase are also reported in Thellungiella halophila from salt treated seedlings (Zhang et al., 2008).
 View Details | Table 3 Genes related to stress tolerance |
Presence of NaCl imposes both ionic and osmotic stress on plants. Salt stress causes physiological drought to the plants and activates the drought responsive genes. The damage due to salt stress in plants is not only because of toxicity of Na+ ions but also a response to hyper osmotic stress. We found the drought resistance genes like Abscisic acid and stress induced protein, early response to dehydration (erd) proteins in Salicornia ESTs. This gene shows expression in drought condition in Arabidopsis (Kiyosue et al., 1994).
Ion transporters help in achieving greater tolerance by maintaining cytosolic ion homeostasis. Ions transporters selectively transport ions and maintain them at physiologically relevant concentration. Genes related to ion transporters like ABC transporters are reported in the present ESTs. Secondary active transport and electrochemical flux across the plasma membrane and tonoplast are driven by H+ pump and VATPase. Salicornia ESTs represents the plasma membrane associated H+ATPase and VATPaseC encoding genes, however, tonoplast H+-PPase was not present. Similary in Suaeda (Zhang et al., 2001), Thellungiella (Wang et al., 2004) the V-type ATPaseC subunits were also reported. The Salicornia PM-H+ATPase showed 75% identity with M. crystallinum. In Tamarix both plasma membrane H+ATPase and VATPaseC ESTs were present. In S. salsa Na+ accumulated seedlings showed increased V-ATPase activities induced by salt treatment and no clear pattern was detected for NaCl-dependent P-ATPase and V-PPase activity changes. Thus, P-ATPase and V-PPase seem to be less important physiologically than V-ATPase in S. salsa under salt stress (Wang et al., 2008). Salicornia plant similar to Suaeda accumulates salt in its succulent tissue. Previous studies with Salicornia demonstrated the accumulation of NaCl in pith region (Reddy et al., 1993), reflecting that the intercellular space provides more room for accumulating NaCl and reducing the toxicity in cytoplasm. In case of Salicornia, cell to cell transport and membrane transport seem to be a major phenomenon and that may be a possible reason for absence of H+-PPase from the EST in the present study. However, in future we will try to clone this gene using degenerate primers to ascertain its function in ion homeostasis.
The abiotic stresses lead to the production of ROS such as O2, H2O2 and OH– (Price et al., 1989; Moran et al., 1994; Mittler, 2002), which damage membranes and macromolecules. ROS scavenging is an important aspect to combat abiotic stress. Plants produce different antioxidant enzymes such as catalase (CAT), sodium dismutase (SOD), ascorbate peroxidase (APX), glutathione S-transferase (GST) and glutathione reductase (GSR). Salicornia stress induced ESTs also showed different genes related to ROS scavenging. The peroxisomal isoform of ascorbate peroxidase (APX) is a novel membrane isoform that functions in the regeneration of NAD+ and protection against toxic reactive oxygen species (Shigeoka et al., 2002). The intracellular localization and sorting of peroxisomal-APX has been reported and it has been shown to be involved in antioxidant defenses in different cellular compartments during development and abiotic stress (Teixeira et al., 2006). In our ESTs the peroxisomal ascorbate peroxidase is present. pAPX gene was also observed in Thellungiella (Wang et al., 2004) and Suaeda (Zhang et al., 2001). This gene showed high homology with Hordeum vulgare (Shi et al., 2001). We found the thioredoxin M and H3 type in our database, which have earlier been shown to be associated with tolerance to oxidative stress. Different homologs of thioredoxin were expressed in yeast to correlate their function and it was found that TRX m and TRX x, but not TRX f, affected the tolerance to oxidative stress induced by either hydrogen peroxide or an alkyl hydroperoxide (Issakidis-Bourguet et al., 2001). We have also found GST gene in Salicornia ESTs. GST is also reported in EST database of different halophytes, Aeluropus littoralis (Zouari et al., 2007), Thellungiella (Wong et al., 2005). The GST gene is made full length and the sequence has been submitted in the NCBI GenBank (Accession number EF373062) and further being analyzed for its potential role in salt tolerance.
Stress tolerance in plants is a multigenic trait and involves multiple genes and pathways to impart tolerance against the stress (Vinocur and Altman, 2005). Transcription factors are quite important to study the stress tolerance behavior in vivo since these can activate a whole array of genes involved in stress, therefore, can impart tolerance in a more effective way as compare to a single functional gene. The transcription factors are present in large number in the plants. Arabidopsis genome encodes approximately 5.9% transcription factors (Riechmann et al., 2000). In different halophytes transcription factors form an important component of EST database as evident in Aeluropus littoralis (Zouari et al., 2007), Mesembryanthemum (Kore-eda et al., 2004), Suaeda (Zhang et al., 2001), Thellungiella (Wong et al., 2005). In Thellungiella, Wong et al. (2005) have reported that 1% genes belong to transcription or signal transduction elements. In Salicornia ESTs, some important transcription factors are present, like Zn finger, DNA binding protein etc. Zinc finger protein 1 (ThZF1) from Thellungiella showed involvement in drought and salt stress (Xu et al., 2007).
Abscisic acid stress ripening gene (ASR1) was first cloned from tomato encoding a highly charged 13-kDa protein (Iusem et al., 1993) and thereafter isolated from several plant species of dicotyledonous and monocotyledonous plants (Maskin et al., 2001; Carrari et al., 2004). Being developmentally regulated during fruit ripening this gene has also shown the expression in both salt and drought. Over-expression of tomato ASR1 and an ortholog LLA23 from Lilium longiflorum in tobacco (Kalifa et al., 2004) and Arabidopsis (Yang et al., 2005), respectively, showed increased tolerance to salt and drought. We have also isolated full length ASR1 gene from Salicornia. This gene shows 74% identity with M. crystallinum (NCBI accession number AF053566) ABA-stress ripening protein. In Salicornia the ASR1 transcript was observed even without the NaCl treatment, reaching a maximum level at 250 mM NaCl. Time course analysis with 250 mM NaCl reveals that ASR1 transcript peaks at 12 h (Fig. 2a, b). The tomato ASR1 gene was expressed transiently in both roots and shoots, reaching a maximum 12–24 h after the application of salt and dehydration stress (Amitai-Zeigerson et al., 1995). In salt tolerant rice variety Pokkali the transcript was reported even under control conditions, and showed an increase at 6 h on 1% NaCl treatment and thereafter decreased (Vaidyanathan et al., 1999). Similarly, in the present study significant ASR1 transcript was observed under control conditions, indicating that in salt tolerant plants, the ASR1 transcript remains higher in unstressed conditions also.
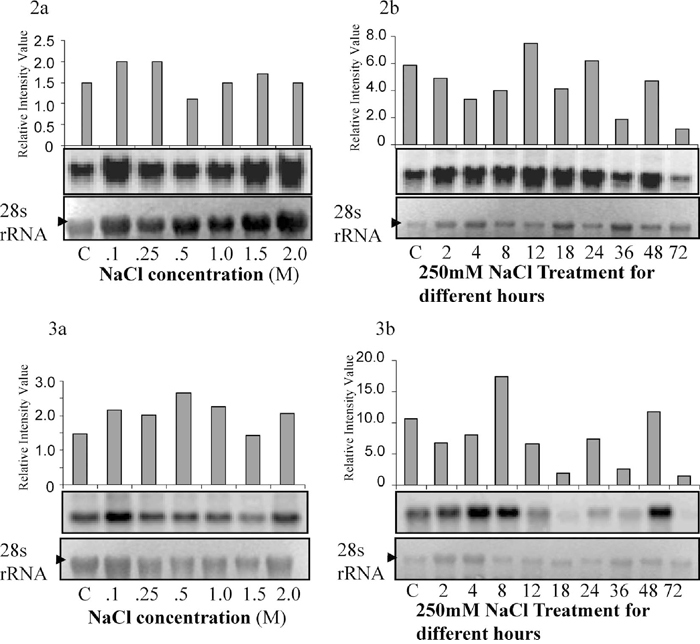 View Details | Fig. 2 Figs.2 and 3. Figures show the Northern analysis of ASR1 gene and PM-H+ATPase gene under different salt concentration (2a and 3a) and at 250 mM NaCl for different time period (2b and 3b). Approximately 20 μg of total RNA was loaded onto each well. Graphic representation over each radiograph shows the relative value of transcript against the 18S rRNA to show the quantitative increase in the expression levels. |
At high salinity plants maintain ion homeostasis to sustain themselves. Plants maintain a low concentration of Na+ in the cytosol by the active exclusion of sodium ions into the apoplast and vacuole by using specific plasma membrane and tonoplast Na+/H+ antiporters. The driving force for this secondary active transport is provided by the PM-H+ATPase and tonoplast H+ATPase (Palmgren, 1998; Ratajczak, 2000). It has been reported that salt treatment of plants induces the activities of the plasma membrane and tonoplast proton pumps (Reuveni et al., 1990; Niu et al., 1993; Perez-Prat et al., 1994; Binzel, 1995; Klobus and Janicka-Russak, 2004). Accumulation of both PM-H+ATPase (Niu et al., 1993; Perez-Prat et al., 1994) and V-H+ATPase (Narasimhan et al., 1991) mRNAs under NaCl-stress and the positive correlation with salt tolerance (Braun et al., 1986; Niu et al., 1993; Perez-Prat et al., 1994) are well studied. In the halophyte Atriplex nummularia Lindl., the proton transport capacity of the PM-H+ATPase has been reported by higher Na+ concentration (Niu et al., 1993). The Na+ stimulation of hydrogen pumping in Atriplex is attributed to increased PM-H+ATPase mRNA leading to higher protein level (Braun et al., 1986). In Salicornia the PM-H+ATPase transcript was found under control conditions, showing maximum expression at 500 mM NaCl. The highest expression was observed at 8 h (250 mM NaCl) treatment (Fig. 3a, b). In S. bigelovii the higher PM-H+ATPase enzyme activity was reported in 200 mM NaCl (Ayala et al., 1996). Whereas, in salt cress T. halophila no changes in the PM-H+ATPase mRNA expression level was observed under salt stress (Vera-Estrella et al., 2005). The time-course analysis of these genes validate that salt tolerance phenomena in Salicornia involve sequential events starting from the constitutive expression of salt responsive genes, the primary induction of key genes followed by expression of other genes.
 View Details | Fig. 3 Figs.2 and 3. Figures show the Northern analysis of ASR1 gene and PM-H+ATPase gene under different salt concentration (2a and 3a) and at 250 mM NaCl for different time period (2b and 3b). Approximately 20 μg of total RNA was loaded onto each well. Graphic representation over each radiograph shows the relative value of transcript against the 18S rRNA to show the quantitative increase in the expression levels. |
We have studied to identify the novelty of the unknown and hypothetical genes by reverse Northern analysis. 90 unknown and hypothetical clones were selected to validate their function in stress tolerance. 21 clones showed up regulation in salt stress, whereas 10 clones showed down regulation (Table 4, Fig. 4). Reverse Northern analysis of a few selected unknown genes from halophyte, smooth cordgrass (Spartina alterniflora Loisel.) revealed much higher abundance of transcripts in response to salt stress (Baisakh et al., 2008). Further analysis of these unknown/unique genes may provide additional stress-associated gene resources from Salicornia.
 View Details | Table 4 Regulation of unknown genes by salt stress |
 View Details | Fig. 4 Figure shows the reverse Northern analysis of the 90 hypothetical/unknown genes. The upper panel (A) is hybridized with salt treated plant’s cDNA and lower panel (B) with control plant’s cDNA. Clone numbers up or down regulated are mentioned on the top panel. |
It is important to isolate ESTs and study known salt responsive genes and unknown genes from halophytes as they have evolved unique mechanisms or regulatory pathways that are not found in glycophytes. Few salt stress related genes like PM-H+ATPase, Abscisic Acid and Stress Ripening Protein (ASR) have shown the alleviated level of expression even in control conditions, reflecting that in halophytes the salt induced genes are significantly expressed even at control conditions to maintain its normal growth and metabolism. It may be plausible that Salicornia recruits most of the known salt responsive genes for its survival in saline condition but some of the unique genes might also play crucial role in defying the high salt concentration. Further comprehensive profiling of unknown/hypothetical gene from the EST data base showed that many unknown genes also have function in salt tolerance mechanism in this extreme halophyte. Most of the stress related genes identified in the present study are consistent with previously published research in other species. In the long term, the most interesting contribution of this work may be the ‘unknown genes’ that have been identified, but of course it requires significant further experimentation.
The authors are thankful to DBT, New Delhi and CSIR, New Delhi (NWP 20) for financial assistance and support.
|