| Edited by Masa-Toshi Yamamoto. Toru Shimada: Corresponding author. E-mail: toru@ss.ab.a.u-tokyo.ac.jp |
In the domesticated silkworm, Bombyx mori, one of the end products of nitrogen metabolism is uric acid. Uric acid is synthesized in the larva and transported to the epidermis; it subsequently accumulates as urate granules, which renders the larval skin opaque, or non-translucent (Tamura and Akai, 1990). Until date, more than 35 translucent mutants have been reported (Fujii et al., 1998). These mutants are caused by defects in the process of synthesis, transport, and accumulation of uric acid (Tamura and Sakate, 1983; Tamura and Akai, 1990). Translucent mutants have easily distinguishable phenotypical characteristics. Therefore, they are considered potential candidate marker genes in the construction of transgenic silkworms (Yasukochi et al., 1998). Thus far, responsible genes for the three translucent mutant, og (MoCoS) (Komoto et al., 2003), oq (BmXDH1) (Komoto, 2002), and od (BmBLOS2) (Fujii et al., 2008) were clarified.
The wild silkworm, Bombyx mandarina, is believed to be the ancestor of Bombyx mori. These two species can be crossed reciprocally and produce hybrid progeny (Kawaguchi, 1928; Murakami and Imai, 1974). Some genes such as wild melanism (Wm; Hirobe, 1947) and wild wing spot (Ws; Hirobe, 1947) which are derived from the Bombyx mandarina were introduced into Bombyx mori strain through the hybridization. In 1982, many individuals of Bombyx mandarina specimens were collected from Hangzhou, China, and reared in our laboratory. During the maintenance of their descendants, several larvae with translucent skin were found in a batch, whose female individuals deposited only unfertilized eggs, while the males were fertile. Therefore, males of the mutant were crossed with Bombyx mori. All the F1 individuals had non-translucent skin. On the other hand, the F2 obtained from sib-mating of the F1 resulted in non-translucent and translucent larvae at a ratio of about 3:1 in both female and male offsprings. These results indicate that the characteristic of translucency was inherited as an autosomal recessive mutation. The mutant was tentatively called ‘‘Ozaki’s translucent’’ and the gene was assigned the symbol oz; it is the first example of the translucent mutation originated from Bombyx mandarina. We have maintained the oz allele by repeated crossing of the +/oz females with either the oz/oz or +/oz males.
The oz/oz phenotype is characterized by highly translucent larval skin, female sterility, high mortality during pupal period and low viability of moths. In the translucent mutants, op, oq, and og locus cause sterility in Bombyx mori (Fujii et al., 1998). In the op strain, only the male moths of mutant homozygotes are sterile. In the oq strains, both the female and male moths of mutant homozygotes are sterile. On the other hand, og causes sterility in females, whereas og/og males are fertile. Additionally, many og individuals perish during pupal period similar to the oz homozygotes. Based on its resemblance to the og phenotype, we speculate that oz is the og gene of Bombyx mandarina.
Here we report genetic analysis of the oz mutant. Based on the crossing experiments, we confirmed that oz is the og gene of Bombyx mandarina. Moreover, sequence analysis revealed that exons 1, 2, and 3 are disrupted in the molybdenum cofactor sulfurase gene.
C108 is a standard strain maintained in our laboratory (Promboon et al., 1995). ogt homozygotes exhibit translucent larval skin (Fig. 1). The o35 strain bears ogt. Homozygotes of oz (ogZ) exhibit translucent larval skin (Fig. 1). The OZ strain having oz (ogZ) gene is maintained in our laboratory.
 View Details | Fig. 1 Larval characteristics of the ogt/ogt, oz/oz (ogZ/ogZ), and oz/+ (ogZ/+). ogt and oz (ogZ) homozygotes exhibit translucent skin. Larval skin of oz/+ (ogZ/+) larva is non-translucent (normal). |
Genomic DNA was extracted from the legs of the moths using InstaGeneTM Whole Blood Kit (Bio-Rad) as described previously (Fujii et al., 2006). Genomic DNA was also extracted from larval posterior silk glands as described by Abe et al. (1998).
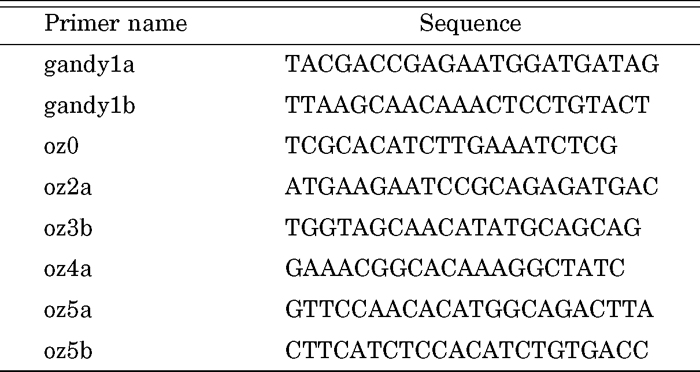
Genomic PCR was performed with ExTaq (TaKaRa) under the following conditions: 94°C for 3 min followed by 35 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 3 min. When the primer set (gandy1a/gandy1b) was used, the cycle number was increased to 40 and elongation time decreased to 1 min. PCR products were analyzed by electrophoresis on 1% or 2% agarose gels and stained with ethidium bromide. The PCR primer sequences used in this study are shown in Table 1. The sequence of a primer, MSR, has been described by Komoto et al. (2003).
 View Details | Table 1 Sequence of primers used in this study |
PCR fragments were purified from gels and cloned into a pGEM-T easy vector (Promega). Cloned DNA was sequenced using the ABI PRISM Big Dye Terminator Cycle Sequencing Ready Reaction Kit and analyzed by the ABI PRISM 3100 Genetic Analyzer.
Total RNA was extracted from the larval fat body using TRIzol reagent (Invitrogen) and treated with deoxyribonuclease (Wako) according to the manufacturer’s instructions. The total RNA isolated was reverse-transcribed, diluted, and used for PCR. First-strand cDNA reaction was performed using the TaKaRa RNA PCR Kit (AMV) Ver. 3.0 (TaKaRa) using an Oligo dT-adaptor primer. PCR was performed using ExTaq (TaKaRa) under the following conditions: 40 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min. The cycle number was decreased to 30 for the primer set (EFRTF/EFRTR). The elongation time was increased to 3 min for the primer set (M13 primer M4/oz4a), and (M13 primer M4/MSF). PCR products were analyzed by electrophoresis on 1% agarose gels and stained with ethidium bromide. Sequences of primers for the MoCoS gene are shown in Table 1. Sequences of the primer set (EFRTF/EFRTR) for the silkworm elongation factor 1α (EF1α) gene have been described by Komoto et al. (2003). The M13 primer M4 is a primer complement to the adaptor region of the Oligo dT-Adaptor primer used in the reverse transcription. It is also provided by the TaKaRa RNA PCR Kit (AMV) Ver. 3.0 (TaKaRa).
Nucleotide sequences flanked by a primer set (oz0/MSR) derived from the ogZ allele were deposited in the DDBJ, EMBL, and GenBank nucleotide databases under the accession number AB426481.
The allelism of oz and the og locus can be tested by crossing the ogt/+ females and oz /+ males. The ogt strain is maintained by crossing the ogt/+ male and ogt/+ female because both the males and females of the ogt homogygotes are sterile. Therefore, we cannot distinguish +/+ and +/ogt individuals in the ogt strain. In the ogt allele, a 551-bp miniature inverted-repeat transposable element (MITE) named Organdy is inserted in exon 5 of MoCoS (Komoto et al., 2003). To discriminate the +/+ individuals from the +/ogt ones in the ogt strain, we constructed a primer set (gandy1a/gandy1b) which amplified the 758-bp fragment containing Organdy from the ogt allele. DNA was extracted from six wild type moths of the ogt strain and the amplification pattern was analyzed using a primer set (gandy1a/gandy1b). The 758-bp band was amplified in the Nos. 1, 3, 4, and 5 moths (Fig. 2), which suggests that (1) the genotypes of Nos. 1, 3, 4, and 5 moths were +/ogt and (2) those of Nos. 2 and 6 were +/+. To confirm the allelism of oz and ogt, three ogt/+ females, and two +/+ females were crossed with oz/+ males obtained by crossing an oz/+ female and an oz/oz male. When oz/+ males were crossed with +/+ females, all larvae exhibited normal skin translucency (Table 2). In contrast, normal and translucent larvae occurred at the ratio of 3:1 in the crosses between oz/+ males and +/ogt females (χ2 = 0.09, P > 0.7) (Table 2). These results indicate that oz is the og gene of Bombyx mandarina. Hence, we revised the genetic symbol of ‘‘Ozaki’s translucent’’ from oz to ogZ.
 View Details | Fig. 2 PCR diagnosis of the genotypes in the ogt strain. DNA extracted from six moths (1–6) of the ogt strain were used as template for PCR. The primer set (gandy1a/gandy1b) was used. M, size marker. Numbers at the left indicate base pairs (bp). |
 View Details | Table 2 Allelism of og locus and oz locus |
A 15-kb genomic region containing the MoCoS gene has been sequenced in the standard Bombyx mori strain, C108 (Komoto at al., 2003) (GenBank accession number AB090243). The gene consists of six or seven exons and there are four types of splicing around intron 1 (Komoto et al., 2003). A variant called type A (Komoto et al., 2003) was shown in Fig. 3. To analyze the genomic structure of the MoCoS gene, we performed genomic PCR analysis. In genomic PCR, two primer sets (oz2a/oz3b and oz5a/MSR) produced amplification products in the normal silkworm larva (C108 strain) (Fig. 4a and 4b), while in the ogZ homozygote, the primer set (oz2a/oz3b) did not produce an amplification product (Fig. 4a). These results suggest that there is deletion in the MoCoS gene. To reveal the structural abnormality of the MoCoS gene, we constructed a PCR primer set (oz0/oz5b) based on the sequence data of the C108 strain (AB090243) (Fig. 3). The oz0 primer is designed from 541 bp upstream of exon 1, and the oz5b primer from exon 5 of the MoCoS gene. We performed genomic PCR using the primer set (oz0/oz5b) in the C108 larvae and the ogZ homozygote. In the C108 larva, a 3.3-kb band was obtained, while a 1.2-kb band was amplified in the ogZ homozygote (Fig. 5). To determine the genomic structure of the MoCoS gene, the PCR products amplified by the two primer sets (oz0/oz5b and oz5a/MSR) were sequenced (AB426481). Subsequent sequence analysis of the fragment disclosed the 2.1-kb deletion containing exons 1 and 2, and the 5' end of exon 3 of the MoCoS gene (Fig. 3 and Fig. 5). The 5' breakpoint was assigned at 4,500 bp in AB020934, and the 3' breakpoint was mapped to 6,641 bp. An orphan sequence of 10 nucleotides (5'-AAAATTAATA-3') was present between the 5' and 3' breakpoints. The 5' flanking region of the breakpoint (1-469 bp in AB426481) had perfect homology with 4,032-4,500 bp in AB020943 except for a T-A substitution. The 3' flanking region of the breakpoint (480–2,426 bp in AB426481) was 98% homologous to the 6,641–8,584 bp region in AB020943. The 2,982–3,332 bp region of AB426481 had perfect homology with 8,907–9,257 bp in AB020943 except for a C-T polymorphism. Additionally, we found a 231-bp insertion sequence (2,535–2,767 bp in accession number AB426481) in intron 6 of the ogZ allele. The insertion makes the difference in the sizes of amplification products in the genomic PCR using a primer set (oz5a/MSR) between +/+ and ogZ/ogZ (Fig. 4b).
 View Details | Fig. 3 Structure of the molybdenum cofactor sulfurase gene in the + and ogZ allele. a, + allele (C108 strain). b, ogZ allele. Primer positions are indicated by black arrowheads, the boxed area indicates exons, and the hatched box indicates the insertion sequence specific to the ogZ allele. The yellow box indicates an orphan sequence (5'-AAAATTAATA-3'), which is not present in the + allele. The red area indicates the deletion in the ogZ allele. Of the four variants involved in the splicing of intron 1 of MoCoS, a variant called type A (Komoto et al., 2003) is shown. |
 View Details | Fig. 4 Genomic PCR analyses of MoCoS in the + allele (C108 strain) and ogZ allele. M. size marker. Numbers at the right indicate base pairs (bp). a, A primer set (oz2a/oz3b) was used. b, A primer set (oz5a/MSR) was used. See Fig. 3 for the primer positions. |
 View Details | Fig. 5 Genomic PCR analysis of the deletion in the ogZ allele. M, size marker. Numbers at the left indicate base pairs (bp). C108 larva is used as a control. A primer set (oz0/oz5b) was used. See Fig. 3 for the primer positions. |
We presumed that MoCoS gene is not expressed in the ogZ homogygotes because the deletion identified in the ogZ allele contains transcript start site of the MoCoS gene identified by Komoto et al. (2003). To detect the transcripts of the MoCoS gene, we performed RT-PCR analysis. However, clear band was not obtained in the RT-PCR using primer sets (oz2a/oz3b and oz5a/MSR) in the C108 larva and the ogZ homozygote (data not shown). Therefore, we performed nested RT-PCR. In one experiment, first PCR was conducted using a primer set (MSF/M13 Primer M4). Second PCR is conducted using a primer set (oz2a/oz3b) (Fig. 6b). In the other experiments, first PCR was conducted using a primer set (oz4a/M13 primer M4). Second PCR is conducted using a primer set (oz5a/MSR) (Fig. 6c). The M13 primer M4 is a primer complement to the adaptor region of the Oligo dT-Adaptor primer used in the reverse transcription. The silkworm elongation factor 1α (EF1α) gene was used as the control for RT-PCR. Expression of the EF1α was detected in the C108 larva and the ogZ homozygote (Fig. 6a). However, expression of the MoCoS gene was not detected in the ogZ homozygote (Fig. 6b and 6c). These results indicates that MoCoS gene was not expressed in the ogZ homozygote due to the deletion containing exons 1 and 2, and the 5' part of exon 3 of the MoCoS. On the basis of these results, we concluded that ogZ is a null mutant of the MoCoS gene.
 View Details | Fig. 6 Nested RT-PCR analysis of the MoCoS in the C108 larva (+/+) and the ogZ homozygote. The Bombyx mori elongation factor 1α (EF1α) gene was used as the control. RNA was extracted from the larval fat body. Numbers at the left indicate base pairs (bp). a, A primer set (EFRTF/EFRTR) was used. b, In the first PCR, a primer set (MSF/M13 Primer M4) was used. In the second PCR, a primer set (oz2a/oz3b) was used. c, In the first PCR, a primer set (oz4a/M13 primer M4) was used. In the second PCR, a primer set (oz5a/MSR) was used. M13 primer M4 is a primer complement to the adaptor region of the Oligo dT-Adaptor primer used in the reverse transcription. See Fig. 3 for the primer positions. |
XDH catalyzes the oxidation of hypoxanthine to xanthine and xanthine to uric acid (Bursell, 1967). XDH requires the sulfide form of molybdenum cofactor (MoCo) for its enzymatic activity (Hill, 1996). MoCoS is responsible for converting the oxo form of Moco to its sulfide form (Amrani et al., 2000; Watanabe et al., 2000). In the ogZ homozygotes, we presumed that the (1) lack of MoCoS results in defective XDH activity leading to the translucent phenotype and (2) accumulation of hypoxanthine and xanthine, the substrates of XDH, lead to high mortality during pupal period and low viability of moths.
According to us, the ogZ homozygotes are fertile although they occasionally need assistance for copulation. Besides ogZ, there are two alleles in the og locus. Males of og homogygotes is fertile while that of ogt or ogk homozygotes are sterile. In the og homozygotes, XDHβ activity was absent while faint XDHα activity was present. In the ogt or ogk homozygotes, both XDHα and XDHβ activity were absent (Komoto et al., 2003). The oq locus encodes the XDH1 gene (Komoto, 2002). oq homogyzotes lack XDHα activity and are unable to synthesize uric acid (Tamura, 1977, 1983). Similar to the ogt or ogk homozygotes, males of oq homogyzotes is sterile. Male sterility of the ogt homozygotes and the oq homozygotes can be rescued by the injection of the xanthin oxidase which is an equivalent enzyme of xanthin dehydrogenase. It is concluded that male sterility of them are caused by the defect in the XDH activity (Tamura et al., 1999). Therefore, it is unclear why ogZ/ogZ males are fertile although MoCoS gene is not expressed. For the better understanding of the effect of the MoCoS and XDH on the male fertility, XDH activity should be analyzed in the ogZ homogygotes.
The ogt strain, o35, was kindly provided by Dr. Yutaka Banno, Silkworm Genetics Division, Institute of Genetic Resources, Graduate School of Bioresource and Bioenvironmental Sciences, Kyushu University, Fukuoka, Japan. This work was supported by Grant-in-Aid for Scientific Research on Priority Areas "Comparative Genomics" (MEXT), National Bioresource Project (MEXT/JST), Agrigenome Research Program (MAFF-NIAS), and Professional Program for Agricultural Bioinfomatics (JST), Japan.