| Edited by Toru Terachi. Hirokazu Handa: Corresponding author. E-mail: hirokazu@affrc.go.jp. Note: The nucleotide sequence reported here is available in the DDBJ/EMBL/Genbank databases under the accession number AB569606. |
The mitochondrial genomes of higher plants are reported to be larger and more complex than their counterparts in animals and fungi (reviewed in Kubo and Newton, 2008). The mitochondrial genomes of higher plants range in size from 200 kb in Brassica species to more than 2,400 kb in cucurbits, which is 10 to 100 times larger than mammalial genomes. In addition to being large, higher plant mitochondrial genomes are the most complex mitochondrial genomes currently known, and they have undergone frequent genome rearrangements over time. Due to their active recombination via large repeated sequences (longer than 1 kb), multiple molecules, including master chromosomes and subgenomes co-exist in the mitochondrion. This type of genomic arrangement is known as the multipartite structure (Fauron et al., 1995). Small sequence repeats (shorter than 1 kb, usually several dozen or hundreds of bp) also contribute to genome rearrangements, although the frequency of these events is not as high. Several nuclear genes are reported to affect the structure of the Arabidopsis mitochondrial genome, whose lesions result in increasingly irregular recombination events, especially in the case of Msh1 (Abdelnoor et al., 2003), RecA3 (Shedge et al., 2007) and OSB1 (Zaegel et al., 2006).
In some cases, recombination events generate a novel expressed ORF composed of chimeric sequences associated with CMS (cytoplasmic male sterility) (Chase, 2007; Hanson and Bentolila, 2004). These complex genome rearrangements can result in heterogeneous DNA molecules, one of which is the predominant DNA molecule. The other substoichiometric molecule is present at very low levels. The balance between the predominant and substoichiometric DNA molecules can be shifted by a process called substoichiometric shifting (Small et al., 1989). Bellaoui et al. (1998) conducted PCR analyses of the mitochondrial genomes of rapeseed cybrid plants and revealed substoichiometric molecules bearing different configurations of the gene (orf138) responsible for Ogura CMS in Brassicaceae. They reported that these substoichiometric molecules were produced through dynamic recombination, that their maintenance occurred via an active mechanism, and that changes in the dynamic equilibrium of molecules carrying the orf138 gene were related to the reversion of male-sterile phenotypes.
CMS rapeseed was produced by asymmetrical somatic cell fusion between the Brassica napus cv. Westar and the Raphanus sativus Kosena CMS line (Sakai and Imamura, 1992). This CMS phenotype with a rapeseed background is stable under various environmental conditions, and fertility is restored by the presence of the fertility-restoring gene (Rf) from Kosena radish, orf687 (Koizuka et al., 2003). The causal gene in this CMS rapeseed is orf125, which is homologous to orf138, the Ogura radish CMS gene, except for two amino acid substitutions and a 39 bp deletion in the orf138 coding region (Iwabuchi et al., 1999). Yet the structure of orf125-surrounding region in the mitochondrial genome of CMS rapeseed is different from that of orf138 shown in Bellaoui et al. (1998). To elucidate how the orf125 region in CMS rapeseed was created, we sequenced orf125 and its surrounding regions in CMS rapeseed, Kosena radish and Westar and carried out comparative studies of the genomic organization and genome rearrangements of these three lines using extensive PCR analyses.
The CMS rapeseed used in this study originated from the cybrid 18-3 by ‘donor-recipient’ protoplast fusion of iodacetoamide-treated protoplasts of Brassica napus cv. Westar (recipient) with X-ray irradiated protoplasts of the Raphanus sativus Kosena CMS line (donor) (Sakai and Imamura, 1992). The parental Westar (recipient) and Kosena (donor) lines of the protoplast fusion were also used.
Mitochondria were isolated from green leaves of 4-week-old plants, and mitochondrial DNA (mtDNA) was prepared using procedures described previously (Itani and Handa, 1998). Five μg of mtDNA was digested with NcoI, EcoRV and XbaI, separated by electrophoresis in 1.0% agarose gels and transferred to Hybond-N+ nylon membranes (GE Healthcare, UK). The ECL Direct Nucleic Acid Labeling and Detection System (GE Healthcare) was used for probe labeling, hybridization and signal detection. To isolate the upstream region of orf125 in CMS rapeseed, we performed a genome walk using gene-specific primers in accordance with Clontech’s Genome Walker™ Universal Kit user manual (protocol no. PT3042-1; Clontech, CA, USA). Nucleotide sequences were determined using the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, CA, USA) with an Applied Biosystems 3130xl genetic analyzer. For each PCR reaction, 1 ng mtDNA was used as template DNA and PCRs using TaKaRa ExTaq DNA polymerase (TaKaRa, Japan) were performed with a MyCycler thermal cycler (Bio-Rad, CA, USA) according to the following protocol: initial denaturation for 5 min at 94°C; 28 or 36 cycles of incubation for 30 sec at 94°C, 30 sec at 58°C and 1 min at 72°C; followed by a final extension for 5 min at 72°C.
Oligonucleotide sequences used for PCR analyses in this study are as follows.
atpA-F: 5'-TAGCCAGCTCACTTCCCTCGTTCG-3'
atpA-R: 5'-AATGTCATCATAAGGGTGTTGTTCC-3'
ccmFN1-F: 5'-AATGAGCTTTGCTCCTCTGGACGC-3'
Nco2.2-F: 5'-CCTTCCGCTTCAACCCAATCTTAC-3'
orf125-F: 5'-CCAAAGCTGAACATATCCGCACAG-3'
orf125-R: 5'-GCGCCAAATAGCTTCTAGTTAGGC-3'
orfB-F: 5'-AGGAGAAGGATCTTCGAGATCTGC-3'
orfB-R: 5'-CTCAGTGGGCATCATAAAGAGCTC-3'
The 7,550 bp DNA sequence of the mitochondrial genomic region that covers CMS rapeseed orf125 was determined (Fig. 1). As previously reported by Iwabuchi et al. (1999), this region includes a 378-bp open reading frame, which encodes the ORF125 protein, a Kosena CMS-associated mitochondrial protein, and a tRNA gene for initiator methionine (trnfM) located 182 bp upstream of the orf125 gene. Homology searches against the complete mitochondrial genome of normal rapeseed (Handa, 2003) determined that the region downstream (3,254 bp) of the 32 bp position before the termination codon of orf125 was identical to a region between exon C of the nad1 gene (nad1C) and ccmFN1. Conversely, the region upstream of orf125 (4,496 bp) showed no homology to the rapeseed mitochondrial genome sequence. The 3’ half of the orf125 upstream sequence was identical to the Kosena orf125/orfB region, as reported by Iwabuchi et al. (1999), and the remaining upstream sequence was postulated to derive from the mitochondrial genome of Kosena radish. Comparison of the orf125 region of CMS rapeseed to the normal rapeseed nad1C/ccmFN1 region and the Kosena orf125/orfB region revealed a 63 bp sequence commonly located in the rapeseed nad1C/ccmFN1 region and the Kosena orf125/orfB region. This result suggested that recombination occurred between the mitochondrial nad1C/ccmFN1 and orf125/orfB regions within this 63 bp repeat sequence, which produced the orf125 region in CMS rapeseed.
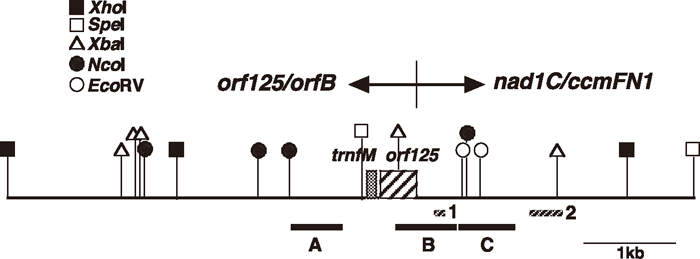 View Details | Fig. 1 A physical map of the orf125 region in CMS rapeseed. Coding regions are shown with boxes, the positions of genomic regions used as probes for Southern analyses are indicated with black bars, and the hatched bars indicate the positions of the genomic regions shown in Fig. 3. |
Southern hybridization analyses using three mitochondrial genomic regions of CMS rapeseed (see Fig. 1; probes A, B and C, which correspond to upstream, coding and downstream sequences of orf125, respectively) were performed to examine whether the corresponding sequences were present in CMS rapeseed, normal rapeseed (Westar) and Kosena radish. Probe A hybridized to the 6.1 kb and 16.8 kb fragments from CMS rapeseed and Kosena radish, respectively (indicated as a circle and a filled circle, respectively, in the EcoRV lanes in panel A of Fig. 2), but no hybridization signals were observed in normal rapeseed, demonstrating that this region originated from Kosena radish and not from rapeseed. Hybridization with probe B yielded one (4.2 kb) and two fragments (4.2 kb and 6.1 kb) in normal rapeseed and CMS rapeseed mitochondria, respectively (indicated as a triangle and a circle, respectively, in the EcoRV lanes in panel B of Fig. 2). Probe B covered the junction between the orf125 region and the nad1C/ccmFN1 region, which indicates the 4.2 kb hybridized fragment in rapeseed corresponding to the nad1C/ccmFN1 region. Two fragments were detected in CMS rapeseed corresponding to the orf125 region (6.1 kb) and the nad1C/ccmFN1 region (4.2 kb) (in the EcoRV lanes in panel B of Fig. 2). Probe B also identified two fragments, 4.2 kb and 16.8 kb, in Kosena radish, which likely corresponded to nad1C/ccmFN1 and orf125/orfB regions, respectively (indicated as a triangle and a filled circle, respectively, in the EcoRV lanes in panel B of Fig. 2).
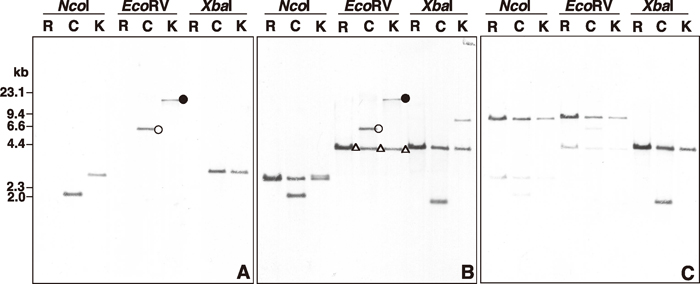 View Details | Fig. 2 Southern hybridizations to normal rapeseed (R), CMS rapeseed (C) and Kosena radish (K) mitochondrial DNAs. Mitochondrial DNAs were digested with NcoI, EcoRV and XbaI. Probes, A, B and C used in this study are shown in Fig. 1 and in MATERIALS AND METHODS. |
These hybridization results revealed that the nad1C/ccmFN1 region (4.2 kb) was located in all three mitochondrial genomes, which is supported by the hybridization data with probe C (corresponding to the nad1C/ccmFN1 region) in panel C of Fig. 2. In addition to the nad1C/ccmFN1 region, CMS rapeseed and Kosena radish have the orf125 region (6.1 kb) or the orf125/orfB region (16.8 kb) in their mitochondrial genomes, respectively.
Sequence analyses of the orf125 region in CMS rapeseed showed that the orf125 region in CMS rapeseed could have originated from recombination via 63 bp repeats between the nad1C/ccmFN1 and orf125/orfB regions, as shown in Fig. 1. In contrast, Southern analyses revealed that Kosena radish had both the nad1C/ccmFN1 and the orf125/orfB regions (the 4.2 kb EcoRV fragment in Fig. 2B and the 16.8 kb EcoRV fragment in Fig. 2A and Fig. 2B, respectively) in its mitochondrial genome, but no hybridization signal corresponding to the orf125 region was found in CMS rapeseed (6.1 kb EcoRV fragment in Fig. 2A and Fig. 2B). Why was the 6.1 kb EcoRV fragment corresponding to the CMS rapeseed-type orf125 region missing in Kosena radish despite the fact that it had the nad1C/ccmFN1 and orf125/orfB regions in its mitochondrial genome? To answer this question, we sequenced the nad1C/ccmFN1 regions in the mitochondrial genomes of normal rapeseed, CMS rapeseed and Kosena radish and compared the sequences of the three nad1C/ccmFN1 regions, the orf125 region in CMS rapeseed and the orf125/orfB regions in Kosena radish.
DNA sequences for regions upstream of the 63 bp repeats (including the orf125 coding sequence) are completely identical between the CMS rapeseed orf125 region and the Kosena radish orf125/orfB region (data not shown), and DNA sequences for regions downstream of the 63 bp repeats corresponding to the nad1C/ccmFN1 region are also completely identical between CMS rapeseed and normal rapeseed (Fig. 3A). However, several nucleotide changes and insertions/deletions were identified between the nad1C/ccmFN1 regions of CMS rapeseed and Kosena radish. These results indicate that an intermolecular recombination occurred between the orf125/orfB region of Kosena radish and the nad1C/ccmFN1 region of normal rapeseed via the 63 bp repeats and resulted in the creation of the orf125 region in CMS rapeseed.
 View Details | Fig. 3 (A) DNA sequence alignment of the nad1C/ccmFN1 region in CMS rapeseed, normal rapeseed and Kosena radish (corresponding to the region from 5806 to 6154 of CMS rapeseed; DDBJ accession number, AB569606). The genomic position of this DNA sequence is shown in Fig. 1 by the hatched bar and nucleotide differences among the three mitochondrial genomes are highlighted. (B) Sequence comparison of the part of fragment B (shown in Table 1) in CMS rapeseed and Kosena radish and the part of fragment A in normal rapeseed (corresponding to the complementary strand from 4714 to 4718 of CMS rapeseed; DDBJ accession number, AB569606). The fragment from each line was sequenced directly (without cloning). |
Bellaoui et al. (1998) reported that three tandem sequences in the orf138 region, a gene responsible for CMS in Ogura radish, were involved in dynamic and complicated recombination events that resulted in the creation of substoichiometric molecules and the reversion of the male sterile phenotype in some cybrid strains. orf125 is a form of orf138 with a 39 bp-deletion, and the genomic structures are conserved between the orf138 region in Ogura radish and the orf125/orfB region in Kosena radish. Considering this relationship, we analyzed the presence of three tandem sequences reported by Bellaoui et al. (1998) and the potential recombination events that may have been mediated by these repeats. We identified the presence of three tandem sequences, R1, R2 and R3, between orf125 and orfB in the Kosena radish (Fig. 4), which correspond to those identified by Bellaoui et al. (1998), although their sizes differ slightly from those reported in the previous study. R3 is located upstream of R1 and corresponds to the 63 bp repeat shown above, and R2 is an internal part of R1 (Fig. 4). The presence of these repeat sequences in Kosena radish shows that they were also involved in dynamic recombination events in our plant materials, as shown in the study of Bellaoui et al. (1998). To investigate such a possibility, we analyzed possible recombination events via R2 and R3 in CMS rapeseed, normal rapeseed and Kosena radish using PCR and sequence analyses. The results are summarized in Table 1, Fig. 5 and Fig. 6.
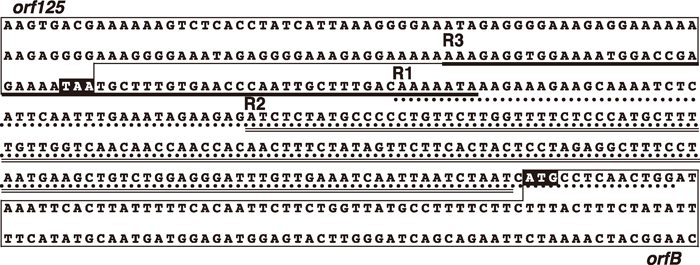 View Details | Fig. 4 Three repeat sequences between orf125 and orfB in Kosena radish (R1, R2 and R3 indicated by a hatched line, double line and thick line, respectively). The coding regions of orf125 and orfB genes are boxed with highlighted termination and initiation codons. |
 View Details | Table 1 Summarized results of recombination products by PCR |
 View Details | Fig. 5 Assessment of the recombination status around orf125 and their surrounding regions in normal rapeseed, CMS rapeseed and Kosena radish using PCR fragment analyses. The photograph in the upper row shows the results of 28 cycles of PCR amplification, and the results from 36 cycles of PCR amplification are shown in the lower row. A-K correspond to the fragments summarized in Table 1. m, molecular weight marker. |
 View Details | Fig. 6 Schematic representation of the R2 (A) or R3 (B) repeat-mediated recombination events in Kosena radish, rapeseed and CMS rapeseed, which yielded the fragments described in the text and Table 1. Solid horizontal lines indicate the fragments detected by PCR, and dashed lines indicate the fragments never found in this study. The boxes shown on the lines indicate the genes. The arrows between columns indicate a direction and a possible frequency of recombination. The boxes under the lines indicate the R2 or R3 repeat sequences. The primers are represented by horizontal arrowheads under the lines. |
Recombination events through R2 were observed in all three lines. In Kosena radish, R2 participated in the recombination between fragments C and I, which resulted in the emergence of fragments E and G at the substoichiometric levels (Fig. 5 and Fig. 6). Similarly, recombined fragments G and H and fragments G and K were detected at substoichiometric levels from fragments F and I in CMS rapeseed and fragments I and J in normal rapeseed, respectively (Fig. 5 and Fig. 6). These observations showed that sequence R2 was an active site of recombination in the mitochondrial genomes of the three lines under this study.
R3 could participate in recombination between fragments A and C in Kosena radish (Table 1 and Fig. 6). We detected the recombined fragments B and compared its intensity with those of fragment E and G using a software, NIH ImageJ (http://rsb.info.nih.gov/ij/) (Fig. 5). The amount of fragment B was quite low - about six times less than the levels of fragments E and G, which represent the substoichiometric products of a recombination event between fragments C and I via R2 in Kosena radish. Sequence analysis of a part of fragment B in Kosena radish and CMS rapeseed and comparison of them with the sequence of fragment A in normal rapeseed revealed that fragment B in Kosena was different from fragment B in CMS rapeseed and not related to fragment A in normal rapeseed (Fig. 3B). The other fragment, D, was never found in Kosena radish. In CMS and normal rapeseeds, we did not detect any recombination events via R3 (Fig. 6). These results indicate that the recombination via R3 was a rare event in the mitochondrial genome of Kosena radish, which is usually suppressed to a very low level.
Sequence analyses of the orf125 region in CMS rapeseed showed that the orf125 region in CMS rapeseed may have originated from recombination by way of R3 between the nad1C/ccmFN1 and orf125/orfB regions (Fig. 1). Precise sequence comparisons of the corresponding regions between CMS rapeseed and normal rapeseed or Kosena radish revealed that regions upstream of R3 (including the orf125 coding sequence) were completely identical to the Kosena radish orf125/orfB region, and that the region downstream of R3 in CMS rapeseed corresponding to the nad1C/ccmFN1 region was completely identical to the same region in normal rapeseed and not to that in Kosena radish (Fig. 3A). Southern hybridization analyses showed that Kosena radish had both the nad1C/ccmFN1 and orf125/orfB regions in its mitochondrial genome, but there was no hybridization signal corresponding to the orf125 region of CMS rapeseed (Fig. 2). These results indicate that the orf125 region in CMS rapeseed was generated from recombination between the orf125/orfB region of Kosena radish and the nad1C/ccmFN1 region of normal rapeseed via the R3 repeats during the cell fusion process.
In many instances, recombination and the resulting substoichiometric shifting (changes in the relative levels of molecules) were observed in cell culture and in regenerated plants (Hartmann et al., 1992; Kanazawa et al., 1994; Bartoszewski et al., 2004). Mackenzie and colleagues revealed that Msh1, a gene involved in the suppression of mitochondrial DNA rearrangements, appeared to be involved in the suppression of illegitimate recombination in plant mitochondria (Abdelnoor et al., 2003, 2006). Using an RNAi strategy, Sandhu et al. (2007) suppressed Msh1 expression in tobacco and tomato, resulting in reproducible mitochondrial DNA rearrangements and male sterility. Moreover, they observed that transgene segregation did not reverse male sterile phenotypes, which indicates that the mitochondrial DNA rearrangement induced by RNAi-mediated Msh1 repression was not reversible. Once rearrangement occurs, transgene removal can permit retention of the stably altered cytoplasm. The process of cell fusion is thought to be a severe manipulation for plant cells. During the cell fusion step, it is possible that the normal and tight regulation of mitochondrial DNA rearrangements is temporarily deregulated, which is similar to repression of Msh1 by RNAi. This deregulation can lead to mitochondrial DNA rearrangements that are prohibited under normal cell conditions.
To analyze the possibility of substoichiometric recombination more precisely, we carried out a PCR analysis to detect submoler recombined products. We detected very low amount of the fragment B, which corresponds to the orf125 region of CMS rapeseed (Fig. 5). To elucidate the mechanism by which the orf125 region in CMS rapeseed was generated from the rapeseed nad1C/ccmFN1 and the Kosena orf125/orfB regions, we performed direct sequence analyses (without cloning) of PCR-amplified fragments corresponding to the CMS rapeseed orf125 region, the normal rapeseed nad1C/ccmFN1 region and the Kosena orf125/orfB region. As shown in Fig. 3B, sequence comparisons revealed that the orf125 region in CMS rapeseed was comprised of the recombination between rapeseed nad1C/ccmFN1 and Kosena orf125/orfB regions, and there was no indication that the orf125 region was derived from the recombination between the Kosena nad1C/ccmFN1 and the Kosena orf125/orfB regions. These results clearly support the hypothesis that an intermolecular recombination event occurred between the orf125/orfB region of Kosena radish and the nad1C/ccmFN1 region of normal rapeseed, creating the orf125 region in CMS rapeseed.
If mtDNA rearrangements are deregulated during the cell fusion, there also will be a chance for recombination between two regions in radish mtDNA. Yet we did not detect any evidence for the presence of the orf125 region originating from recombination between the orf125/orfB of Kosena type and the nad1C/ccmFN1 region of Kosena type in CMS rapeseed. To explain this observation, we postulate two working hypotheses. First, nuclear factor(s) can detect the difference between the Kosena-type and rapeseed-type nad1C/ccmFN1 regions, and these factors are involved in selective rearrangements using only the nad1C/ccmFN1 region of rapeseed. The other hypothesis is that two type orf125 fragments, a Kosena-type and a rapeseed-type, are produced during cell fusion, but the Kosena-type fragment is eliminated by selelctive pressure after several back crosses with rapeseed.
In the present study we characterized the recombination status of two repeat sequences, R2 and R3, between orf125 and orfB in Kosena radish (Fig. 4), which were completely different. As discussed previously, the R3 sequence that recombined to generate the orf125 region in CMS rapeseed is recombination-inactive under normal cell conditions. In contrast, R2 appears to be an active site of mitochondrial DNA recombination, regardless of cell conditions.
Although R2 and R3 are located adjacent to each other, why are there differences in the recombination activity of these two repeat regions? One possibility is that recombination via each repeat is regulated by different nuclear genes. In Arabidopsis, Msh1 suppresses recombination via short repeat sequences ranging in size from 108 to 556 bp, while shorter repeats of less than 108 bp do not respond to Msh1 (Arrieta-Montiel et al., 2009). On the other hand, RecA1 functions as a suppressor of recombination involving repeats of less than 100 bp in Physcomitrella (Odahara et al., 2009). The size of R2 (143 bp) is within the size that Msh1 can mediate recombination (from 108 to 556 bp), while R3 (63 bp) may be a target of RecA1 regulation (less than 100 bp). It will be very interesting to reveal the relationships between the recombination activities of these repeats and nuclear factors such as Msh1 and RecA1 in the future.
This work was supported in part by a Grant-in-Aid for Scientific Research (B) from the Ministry of Education, Culture, Sports, Science and Technology of Japan (No. 18380007).
|