| Edited by Kazuo Yamamoto. Shogo Ikeda: Corresponding author. E-mail: ikeda@dbc.ous.ac.jp |
Endogenous and environmental alkylating agents generate alkyl base lesions, which may result in arrest of replication and transcription, mutagenesis, and/or cytotoxicity (for review see Friedberg et al., 2006). In humans these lesions could be involved in carcinogenesis, neurodegenerative disease and aging. Methyl methanesulfonate (MMS) is an SN2-type alkylating agent that predominantly methylates nitrogen atoms in purines (Beranek, 1990). The most frequent effect of DNA methylation is the generation of 7-methylguanine (7-meG) and 3-methyladenine (3-meA). 7-meG is relatively harmless, because this modification does not alter the coding specificity of the base. However, destabilization of the glycosidic bond and imidazole ring due to N7 substitution on guanine results in generation of a mutagenic apurinic (AP) site and ring-opened methyl-formamidopyrimidine, which efficiently inhibits DNA replication in vitro and in Escherichia coli (Tudek, 2003). 3-meA is a strong cytotoxic lesion that blocks both replication and transcription due to the aberrant methyl group protruding into the minor groove of the DNA double helix (Larson et al., 1985; Racine et al., 1993; Sedgwick et al., 2007). 3-meA is more susceptible to be hydrolyzed into the AP site than 7-meG (Beranek, 1990). While S-adenosylmethionine is the donor of methyl groups in enzymatic methylation events in vivo, it acts by the SN2 mechanism and yields the same products on alkylation of DNA as MMS (Sedgwick et al., 2007).
In many organisms alkyl base lesions such as 3-meA and 7-meG are mainly corrected by base excision repair (BER), in which damaged bases are removed by a lesion-specific DNA glycosylase, in this case 3-meA DNA glycosylase. BER engages other universal proteins, including AP endonuclease, AP lyase, DNA polymerase, and DNA ligase. E. coli possesses two 3-meA DNA glycosylases, Tag and AlkA. Tag only removes 3-meA and 3-methylguanine, whereas AlkA removes a wide variety of damaged bases including 3-meA and 7-meG (Friedberg et al., 2006). AlkA family homologs are not present in mammalian cells, but the budding yeast Saccharomyces cerevisiae has one homolog Mag1p, and the fission yeast Schizosaccharomyces pombe has two homologs Mag1p and Mag2p (Friedberg et al., 2006; Sedgwick et al., 2007). Mammalians possess another type of 3-meA DNA glycosylase with a broad spectrum of action toward alkylation-damaged DNA. Although inactivation of 3-meA DNA glycosylase genes in E. coli, S. cerevisiae, and mouse cells dramatically increased MMS sensitivity, loss of the S. pombe Mag1p and Mag2p does not confer MMS sensitivity (Memisoglu and Samson, 2000; Kanamitsu et al, 2007). This indicates that different species employ the 3-meA DNA glycosylase-initiated BER to different extents.
Genetic analyses of S. pombe showed that nucleotide excision repair (NER) and homologous recombination (HR) repair contribute greatly to alkylation resistance of S. pombe (Memisoglu and Samson, 2000; Alseth et al., 2005). Mutations in genes involved in NER (rad13 and rad16) and HR (rhp51 and rhp55) are much more sensitive to MMS than the mag1Δ mutant. However, mutants lacking the genes for AP endonuclease (apn2), DNA glycosylase/AP lyase (nth1) and flap endonuclease (rad2) are sensitive to MMS, indicating that BER is also important for the alkylated DNA repair (Osman et al, 2003; Alseth et al., 2004; Ribar et al., 2004; Sugimoto et al., 2005). Moreover, deletion of the rad16 (or rad13) gene from mag1Δ and mag2Δ strains greatly enhanced MMS sensitivity (Memisoglu and Samson, 2000; Alseth et al., 2005; Kanamitsu et al, 2007), suggesting that BER and NER synergistically function to sense alkylated bases in DNA. Genetic interaction of mag1 and rhp51 (or rhp55) also showed the synergism between BER and HR (Memisoglu and Samson, 2000; Alseth et al., 2005).
NER is a versatile DNA repair system that eliminates a broad spectrum of DNA lesions that tend to distort the double helix (Friedberg et al., 2006; Sugasawa, 2008). There are two overlapping pathways, global genome repair (GGR), which can operate anywhere in the genome, and transcription-coupled repair (TCR), which is specialized to correct lesions from the transcribed strand of an active gene. These subpathways differ principally at the levels of DNA damage recognition. In human GGR, the XPC-HR23B complex is responsible for initial lesion recognition (Maillard et al., 2007; Sugasawa, 2008), whereas damage recognition during TCR is thought to occur when RNA polymerase II is stalled at the lesion (Friedberg et al., 2006; Scicchitano et al., 2004). TCR requires additional proteins that are not needed for GGR, including CSA and CSB. S. pombe possesses two homologs of the human XPC gene, designated rhp41 and rhp42 (Fukumoto et al., 2002; Marti et al., 2003). Rhp41p has a major function in removing cyclobutane pyrimidine dimers in the transcribed and non-transcribed strands, while Rhp42p is involved in the repair of lesions only on non-transcribed DNA. A defect of the S. pombe rhp26 gene, a homolog of human CSB and budding yeast RAD26, slightly increased the sensitivity to UV irradiation (Yasuhira et al., 1999). Similar to Rad26p and CSB, Rhp26p likely plays a key role in TCR.
In the present study, we investigated the contribution of NER and HR repair in S. pombe cells in preventing methylation damage-induced toxicity to DNA. In previous studies the rad16 or rad13 gene mutant has been used as a typical NER-defective mutant. These genes encode an endonuclease subunit required for dual incision of the damaged DNA strand, which is a critical step in both GGR and TCR. To clarify how MMS-induced damage is sensed in NER process, several genetic analyses using mutants with defects in lesion recognition steps (rhp41Δ, rhp42Δ and rhp26Δ) were performed. We show that the rhp41 and rhp26 mutants exhibited sensitivity to MMS to the same extent as the rad16Δ strain. Transcription of an inducible gene (fbp1) was strongly inhibited in MMS-treated rhp41Δ and rhp26Δ cells, suggesting the involvement of Rhp41p and Rhp26p in the TCR of MMS-damaged DNA. We also provide genetic evidence that NER and HR pathways are involved in repair of obligate intermediates during BER of MMS damage.
Yeast strains used in this study are listed in Table 1. Target disruption of rhp41, rhp42, and rhp51 genes was performed as follows. The genomic sequence of rhp41 was amplified with the following primers: 5′-CACAAATGATTCGGAACAAAACTTTCC-3′ and 5′-GTACTTCCGTTGAAAATCAAATCC-3′. The gene fragment was then subcloned into pGEM-T vector (Promega Corporation, Madison, WI, USA) by TA cloning. The kanMX6 marker was amplified from pFA6a-kanMX6 (Bähler et al., 1998) by PCR and inserted between XbaI and EcoRI sites of rhp41 cloned in pGEM-T. The rhp41::kanMX6 disruption cassette was amplified by PCR and used to transform haploid strain ED0668 with lithium acetate as previously described (Kanamitsu et al, 2007). The G418-resistant colonies were screened for the correct integration of kanMX6 marker within the rhp41 locus by PCR. The rhp42::kanMX6 and rhp51::kanMX6 disruption cassettes were constructed in a similar manner as above. The genome DNA of rhp42 was amplified with the primers 5′-GACGATTTTGACACTTGGGAACAAG-3′ and 5′-CCCGAAATTCGTGTTCACGC-3′, and cloned in pGEM-T vector. The kanMX6 marker was inserted between EcoRI and AflII sites of rhp42. The amplification of rhp51 genome DNA was performed with the primers 5′-GGCAGATACAGAGGTGGAAATG-3′ and 5′-CATACATATCTATCCTTACAAACTCATCCC-3′. The kanMX6 marker was inserted between EcoRI and AflII sites of the rhp51 gene. Double mutant strains rhp26Δ/nth1Δ, rhp26Δ/apn2Δ, rad16Δ/nth1Δ and rad16Δ/apn2Δ were constructed by random spore analysis followed by selection on appropriate media. Double mutants rhp51Δ/nth1Δ and rhp51Δ/apn2Δ were constructed by targeted gene disruption of the rhp51 gene in nth1Δ and apn2Δ strains, respectively, using an rhp51::kanMX6 disruption cassette. Multiple mutants rhp41Δ/rhp26Δ and rhp41Δ/mag1Δ/mag2Δ were made by disruption of the rhp41 gene in rhp26Δ and mag1Δ/mag2Δ strains, respectively, using the rhp41::kanMX6 disruption cassette. The genotypes of all mutants were verified by PCR.
 View Details | Table 1 Strains used in this work |
Cells were grown overnight in YE medium (0.5% yeast extract and 3% glucose) for survival studies. Cell density was determined by a Z1 particle counter (Beckman Coulter Inc., Brea, CA, USA), and adjusted to 1.0 × 107 cells/ml with YE medium. The cells were diluted 10-fold serially from 10–1 to 10–3 in YE medium. To measure the sensitivity of cells to chronic exposure to MMS, dilutents (3 μl) containing 3 × 101 to 3 × 104 cells were spotted onto YE medium containing MMS (Sigma-Aldrich, St. Louis, MO, USA; 0 to 0.01%) and incubated for 3 days at 28°C. For the UV sensitivity test, serial dilutions were spotted onto YE medium, and irradiated with 254-nm UV light (0 to 100 J/m2) using a FUNA-UV-LINKER (FS-1500; Funakoshi Inc., Tokyo, Japan) equipped with an internal photodetector for measuring the UV dose. Photographs of colonies were taken after 3 days of growth at 28°C.
S. pombe strains were cultured in 500 ml of YE medium at 28°C until OD600 reached between 1 and 2. Two 200-ml portions of the culture were transferred into two culture flasks. MMS was added to one of the two flasks to a final concentration of 0.1%. The cultures with or without MMS were further incubated at 28°C for 1 h with shaking. Four 45-ml portions of each culture with or without MMS were transferred into four 50-ml centrifuge tubes. The cells were washed once with 50 ml of YG medium (0.5% yeast extract and 3% glycerol) by centrifugation, resuspended in 50 ml of YG medium, and then transferred into four separate culture flasks. The four cultures (MMS-treated cells or non-treated cells) were further incubated at 28°C for 0, 1, 2, and 4 h (one flask per time treatment), respectively, with shaking. After incubation, the cells were collected by centrifugation, quickly frozen in liquid nitrogen, and stored at –80°C until RNA extraction.
The cells from each 50-ml culture were washed once with 10 mM Tris-HCl (pH 8.0) and 1 mM EDTA, and resuspended in 0.22 ml of a buffer containing 1 M sorbitol, 0.1 M EDTA, 14 mM 2-mercaptoethanol, and 100 units/ml Zymolyase-20T. After incubation at 30°C for 30 min, total RNA of the sample was isolated using a reagent ISOGEN (Wako Pure Chemical Industries, Osaka, Japan) according to the manufacturer’s instruction. The RNA samples were fractionated by electrophoresis on 1.2% agarose-2.1 M formaldehyde gels, followed by transfer to Hybond-N+ nylon membranes (GE Healthcare UK Ltd, Buckinghamshire, England). The genome sequence of the fructose-1,6-bisphosphatase gene (fbp1) was amplified with the primers 5′-GATCTCGACGAAATCGACAC-3′ and 5′-CCTCGACTTCATGTTTACTTCC-3′, and cloned in pGEM-T vector. Anti-sense RNA of fbp1 labeled with digoxigenin (DIG) was generated by T7 RNA polymerase using a DIG Northern Starter Kit (Roche Diagnostic GmbH, Mannheim, Germany). Hybridization was performed at 60°C overnight in a DIG Eazy Hyb solution (Roche) containing DIG-labeled probe RNA. The blots were washed twice with 2 × SSC-0.1% SDS at room temperature for 5 min and twice with 0.1 × SSC-0.1% SDS for 15 min at 60°C. The fbp1 mRNA was detected with alkaline phosphatase-labeled anti-Dig antibody and CDP-Star (Roche). Imaging of the chemiluminescence was performed with a LAS-1000 Plus luminescent image analyzer system (FUJIFILM, Tokyo, Japan).
As shown in Fig. 1 the defects of 3-meA DNA glycosylases Mag1p and Mag2p did not affect MMS sensitivity of S. pombe cells, suggesting that alternative pathways are involved in recognition and correction of the methylated bases (Memisoglu and Samson, 2000; Alseth et al., 2005; Kanamitsu et al., 2007). Other BER-deficient mutants nth1Δ and apn2Δ showed MMS sensitivity, indicating that BER actually contributes to the repair of MMS damage. In S. pombe cells, AP sites generated by monofunctional DNA glycosylases are mainly processed by AP lyase activity of Nth1p to leave 3′-α,β-unsaturated aldehyde ends, which must be converted to 3′-OH ends by Apn2p (Nth1p-dependent BER) (Tanihigashi et al., 2006: Kanamitsu and Ikeda, 2010). Thus, nth1Δ and apn2Δ cells are thought to accumulate AP sites and 3′-blocked ends, respectively. The apn2Δ strain was more sensitive to MMS than the nth1Δ strain, indicating that the 3′-blocked end is a more deleterious intermediate than the AP site in S. pombe cells. An NER mutant rad16Δ exhibited moderate MMS sensitivity to same extent as nth1Δ. We previously showed that deletion of mag1 and mag2 genes from the rad16Δ strain enhanced sensitivity to MMS (Kanamitsu et al., 2007). This observation raised the possibility that some NER factors could recognize methylated bases such as 3-meA and 7-meG to initiate a repair process. An rhp51Δ strain was the most susceptible to MMS among the mutants with DNA-repair defects tested in Fig. 1. HR repair is essential to repair double strand breaks (DSBs) and single strand gaps produced by replication over blocking lesions including 3-meA and BER intermediates (Nowosielska et al., 2006; Wyatt and Pittman, 2006). Therefore, multiple DNA repair pathways are involved in the repair of MMS-induced damage in S. pombe cells.
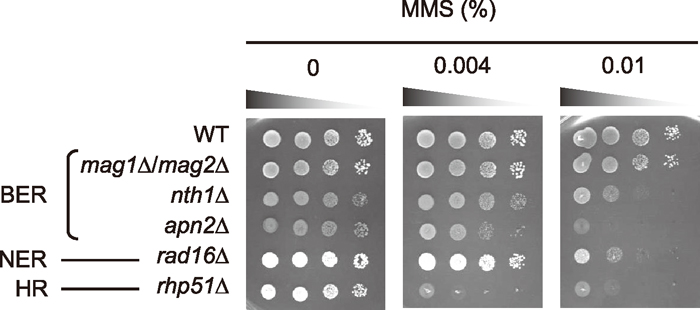 View Details | Fig. 1 Survival assays of various mutant strains for BER, NER, and HR to MMS. Wild-type (WT) cells and various mutants were grown overnight in YE. Survival assays for MMS were carried out as described in MATERIALS AND METHODS. |
S. pombe has two human XPC homologs rhp41 and rhp42, which contribute different roles in the repair of UV-induced DNA damage (Fukumoto et al., 2002; Marti et al., 2003). Consistent with previous studies, the rhp41Δ mutant but not rhp42Δ was sensitive to UV irradiation (Fig. 2A). The UV sensitivity of rhp41Δ was less than that of rad16Δ, which lacks strand incision activity during NER, indicating that Rhp41p recognizes a part of UV damage processed via NER. Surprisingly, rhp41Δ strain exhibited profound MMS sensitivity at or near the level of the rad16Δ strain, suggesting that Rhp41p is substantially involved in the initiation of MMS damage repair. rhp42Δ strain was not sensitive to MMS. Deletion of the rhp41 gene in a BER mutant mag1Δ/mag2Δ greatly increased MMS sensitivity, suggesting that Rhp41p-dependent NER synergistically corrects the MMS-induced damage with Mag1p/Mag2p-initiated BER (Fig. 2B). S. pombe rhp26 is a counterpart to the human CSB gene, and the deletion mutant rhp26 is slightly more sensitive to UV irradiation than wild type cells (Yasuhira et al., 1999). rhp26Δ strain also exhibited MMS sensitivity at the same level as the rhp41Δ strain (Fig. 2C). rhp26Δ/rhp41Δ strain turned out to be more sensitive than each single mutant, indicating that Rhp41p and Rhp26p independently contribute to repair MMS damage.
 View Details | Fig. 2 Survival assays of various mutant strains to UV and MMS. Wild-type (WT) cells and various mutants were grown overnight in YE. Survival assays for UV and MMS were carried out as described in MATERIALS AND METHODS. |
Rhp26p acts similarly to human CSB in the TCR of cyclobutane pyrimidine dimers (Yasuhira et al., 1999). Although the human XPC is required for recognition of UV lesions in GGR, S. cerevisiae Rad4p and S. pombe Rhp41p are also involved in preferential repair of the lesions on the transcribed strand (Verhage et al., 1994; Fukumoto et al, 2002; Marti et al., 2003). To specifically analyze repair of damage in the transcribed strand, we measured transcription of the inducible fbp1 gene after MMS treatment. Expression of fbp1 is induced when cells are shifted from a medium containing 3% glucose to a medium containing 3% glycerol (Vassarotti and Friesen, 1985). In a control experiment, no obvious difference of fbp1 expression in cells not treated with MMS could be observed between the strains (Fig. 3). This indicates that the transcription of fbp1 was not significantly affected by inactivation of BER and NER genes. In MMS-treated cells, however, the levels of fbp1 gene expression after 1 and 2 h were substantially lower in rad16Δ, rhp41Δ, and rhp26Δ strains than in the wild-type strain or mag1Δ/mag2Δ strain. After 4 h, accumulation of fbp1 mRNA in MMS-treated rad16Δ, rhp41Δ, and rhp26Δ strains was restored near the level of non-treated cells. The result indicates that the transcription of fbp1 was severely inhibited in rhp41Δ and rhp26Δ strains by MMS damage and that Rhp41p and Rhp26p are important for initiation of MMS-damage repair in the transcribed strand. A similar inhibition in the transcription of the fbp1 gene has been observed in rhp41Δ strain after UV irradiation (Marti et al., 2003). Inhibition of transcription in MMS-treated rad16Δ strain implies that the lesions are repaired via the NER pathway. A Mag1p and Mag2p defect did not affect transcription of the fbp1 gene, suggesting that Mag1p/Mag2p-initiated BER contributes to repair the lesions mainly on a non-transcribed gene.
 View Details | Fig. 3 Transcription of the fbp1 gene in MMS-treated wild-type (WT) cells and various mutants. Total RNA was extracted from the cells with or without MMS treatment at the indicated time points after induction of the fbp1 gene. Expression of the fbp1 gene was analyzed by Northern hybridization as described in MATERIAL AND METHODS. (Top) fbp1 mRNA detected by Northern hybridization. (Bottom) rRNA of the same sample stained with ethidium bromide, used as loading control. |
Cytotoxic intermediates such as the AP site and the 3′-blocked end are generated during the BER of MMS-induced damage. As described above, AP sites and 3′-blocked ends are thought to accumulate in the BER-deficient mutants nth1Δ and apn2Δ, respectively. Deletion of the rad16 gene from nth1Δ mutants greatly increased the sensitivity to MMS (Fig. 4A), indicating that NER is involved in repair of the AP site. Moreover, a double mutant nth1Δ/rhp26Δ but not apn2Δ/rhp26Δ was more sensitive to MMS than each single mutant (Fig. 4B). In mammalian cells the AP site in the transcribed strand is a strong block to RNA polymerase II, and may be sufficient to initiate TCR in vivo (Tornaletti et al., 2006). Therefore, the hypersensitivity of nth1Δ/rhp26Δ to MMS is probably due to the defect of TCR to process the AP site. The rad16 deletion from the apn2Δ mutant also enhanced sensitivity (Fig. 4A), suggesting that Rad16p provides a backup pathway for removal of 3′-blocked ends. In S. cerevisiae Rad1p-Rad10p nuclease (a counterpart of S. pombe Rad16p-Swi10p) functions in the removal of the 3′ blocked end in vivo and in vitro (Guzder et al., 2004).
 View Details | Fig. 4 Survival assays of various mutant strains to MMS. Wild-type (WT) cells and various mutants were grown overnight in YE. Survival assays for MMS were carried out as described in MATERIALS AND METHODS. |
Previous studies showed that the hypersensitivity of rhp51Δ (or rhp55Δ) to MMS is restored by deletion of the mag1 gene in the mutant strain (Memisoglu and Samson, 2000; Alseth et al., 2005). The results raise the possibility that DNA strand breaks generated during BER could be substrates for HR. Genetic interactions among rhp51 and BER genes (nth1 and apn2) were examined to confirm if HR is involved in repair of the BER intermediates (Fig. 5). Deletion of either the nth1 or the apn2 gene from rhp51Δ strain greatly increased sensitivity to MMS, but did not affect UV sensitivity. This indicates that DSBs and single strand gaps produced by DNA replication at the AP site and 3′-blocked end resulting from BER are the substrates for HR repair.
 View Details | Fig. 5 Survival assays of various mutant strains to UV and MMS. Wild-type (WT) cells and various mutants were grown overnight in YE. Survival assays for UV and MMS were carried out as described in MATERIALS AND METHODS. |
Here we provide the genetic evidence that TCR plays a vital role for correction of MMS-induced DNA damage in S. pombe cells. Inactivation of NER factors Rhp41p and Rhp26p caused moderate MMS sensitivity, and transcription of the fbp1 gene was strongly blocked by MMS in the rhp41Δ and rhp26Δ mutants (Fig. 2 and Fig. 3). TCR is thought to be initiated by stalling RNA polymerase II at a lesion, and in a subsequent step CSA and CSB are required for release of the transcription machinery (Friedberg et al., 2006; Sugasawa, 2008). 3-meA seems to be a probable candidate for activation of TCR, because the lesion is able to hinder translocation of T7 RNA polymerase along the DNA template during chain elongation (Racine et al, 1993). The effect of 3-meA on other RNA polymerases is unknown. The AP sites located in the transcribed strand are potent blocks to eukaryotic RNA polymerase II, and repaired by transcription-coupled NER (Tornaletti et al, 2006; Kim and Jinks-Robertson, 2010). Enzymatic removal of 3-meA and 7-meG by Mag1p and Mag2p produces AP sites. Methylated purine base destabilizes the glycosidic bond and also yields AP site (Tudek, 2003). Consequently, the lesions 3-meA and AP site may initiate TCR in MMS-treated S. pombe cells (Fig. 6).
 View Details | Fig. 6 A schematic illustration of pathways for repair of MMS-induced damage in S. pombe cells. Alkylated bases such as 3-meA are removed by Mag1p and Mag2p, or tend to be spontaneously hydrolyzed into the AP site. In the BER pathway, repair of the AP site is initiated by incision of Nth1p leaving the 3′-block (Tanihigashi et al., 2006: Kanamitsu and Ikeda, 2010). Apn2p functions primarily in the removal of the 3′-block. 3-meA and the AP site inhibit the elongation of transcription by RNA polymerase II. Rhp41p and Rhp26p independently recognize these lesions and initiate TCR. Rhp41p perhaps contributes to GGR of MMS-induced damage. Rad16p-Swi10p nuclease possibly repairs the 3′ blocks as a backup of Apn2p. HR is necessary for repair of DSBs and single strand gaps produced by DNA replication at the lesions such as 3-meA, AP sites and 3′-blocked ends. Details are provided in the text. |
Mammalian XPC associated with HR23B interacts preferentially with the undamaged strand opposite the lesion and is able to indirectly detect a broad spectrum of DNA lesions (Maillard et al., 2007). Unlike XPC, S. cerevisiae Rad4p and S. pombe Rhp41p function in both GGR and TCR for repair of UV-induced damage (Verhage et al., 1994; Fukumoto et al., 2002; Marti et al., 2003). Rhp41p probably contributes to both GGR and TCR of MMS-induced DNA damage including 3-meA and the AP site in fission yeast (Fig. 6). Although Rhp23p, a homolog of human HR23B, has been shown to function as an accessory factor for Rhp41p in GGR and TCR (Lombaerts et al., 2000; Fukumoto et al., 2002), the precise mechanism of damage recognition in Rhp41-dependent TCR is not yet understood.
S. cerevisiae Rad26p is a TCR-specific factor with DNA-dependent ATPase activity, which is recruited to the site of DNA lesion in an elongating RNA polymerase II-dependent manner (Selby and Sancar, 1997; Malik et al., 2010). S. cerevisiae RAD26Δ strain exhibited moderate MMS sensitivity, and its sensitivity to MMS was greatly enhanced by deletion of both NER and BER pathways required for removal of 3-meA and 7-meG (Lee et al., 2002). The transcription of GAL genes through these lesions was severely inhibited in rad26Δ cells lacking NER and BER (Lee et al., 2002). S. pombe rhp26Δ strain also exhibited MMS sensitivity (Fig. 2C). Interestingly, the rhp26Δ/rhp41Δ double mutant was more sensitive to MMS than the NER-defective mutant rad16Δ. This observation implies that Rhp26p functions in part independently of NER. Indeed, budding yeast Rad26p could promote transcription by RNA polymerase II through MMS-induced lesions independently of NER and BER, although Rad26p acts with the NER machinery for the TCR of UV photodamage (Lee et al., 2002).
Synergism between NER and BER in the protection of alkylation damage has been observed in budding yeast (Xiao and Chow, 1998; Torres-Ramos et al., 2000) and mammalian cells (Plosky et al., 2002; Sitaram et al., 1997), even though the 3-meA DNA glycosylase-initiated BER plays a major role in the repair of MMS in these organisms. 3-meA and 7-meG were removed without strand bias from active genes of 3-meA DNA glycosylase-deficient mouse cells (Plosky et al., 2002). In contrast to the clearance of N-methylpurines, N-ethylpurines were removed preferentially from the transcribed strand in NER-proficient mammalian cells, perhaps because a larger sized adduct impeded RNA synthesis (Sitaram et al., 1997).
In the Nth1p-dependent BER pathway of S. pombe, repair of the AP site is initiated by incision of Nth1p leaving the 3′-block (Fig. 6). Apn2p functions primarily in the removal of the 3′-block, and poorly in the incision of AP sites. A genetic interaction between rad16 and apn2 suggests that the Rad16p-Swi10p endonuclease (a counterpart of human ERCC1-XPF) provides a backup pathway for removal of 3′-blocked ends (Fig. 6). In this case the NER protein plays a NER-independent role. Similarly, flap endonuclease activity of the budding yeast Rad1p-Rad10p complex is required for removal of 3′ blocked ends from DNA strand breaks induced by reactive oxygen species (Guillet and Boiteux, 2002; Karumbati et al., 2003; Guzder et al., 2004). The S. pombe NER genes rad16, swi10 and rhp14 (XPA homolog) also have a function in msh2/pms1-independent mismatch repair (Fleck et al., 1999).
3-meA itself efficiently blocks DNA replication (Larson et al., 1985), and may provoke DSBs when a replication fork encounters these lesions. The AP site and 3′-blocked single-stranded breaks generated during BER of 3-meA and 7-meG could also block DNA replication (Nowosielska et al., 2006; Wyatt and Pittman, 2006). Ring-opened methyl-formamidopyrimidine derived from 7-meG potently inhibits DNA replication (Tudek, 2003). Additionally, AP sites give rise to DNA strand breaks due to their inherent chemical instability. Synergism between HR and BER genes in repair of MMS-induced DNA damage suggests the necessity of HR to prevent the toxicity of BER intermediates. Therefore, S. pombe HR acts on three different types of damage, 3-meA, AP sites and 3′-blocked ends (Fig. 6).
We would like to thank to Dr. Shinji Yasuhira, Kyoto University, for generously providing the rhp26Δ strain. This work was supported in part by the Ministry of Education, Culture, Sports, Science and Technology of Japan through a Financial Assistance Program of the Social Cooperation Study (2006–2010).
|