| Edited by Hiroshi Iwasaki. Hiroyuki Araki: Corresponding author. E-mail: hiaraki@lab.nig.ac.jp |
DNA replication initiates from specific sequences, called replication origins. Eukaryotic cells have multiple replication origins that fire with various efficiencies and timings. To initiate DNA replication, many proteins assemble on, and dissociate from, origins (See for reviews DePamphilis, 2006; Masai et al., 2010). First, the six-subunit origin recognition complex (Orc, Orc1-6) associates with replication origins throughout the cell cycle in budding yeast, and the hexameric Mcm2-7 complex, which is a catalytic core of DNA helicase, is loaded onto the Orc-bound origins with the aid of Cdc6 and Cdt1 to form the pre-replicative complex (pre-RC) from late M to G1 phases when cyclin-dependent kinase (CDK) is not activated (Fig. 1a). The Sld3-Sld7 complex and Cdc45 form a complex and associate with the pre-RC-formed origins in a mutually dependent manner (Fig. 1b) (Tanaka et al., 2011). This origin association occurs in G1 phase for early firing origins, and in S phase for other origins, in a manner that depends on Dbf4-dependent kinase (DDK or Cdc7 kinase) but not on CDK (Yabuuchi et al., 2006; Heller et al., 2011; Tanaka and Araki, unpublished result). The activation of CDK at G1/S boundary leads to the assembly of other replication proteins on origins to form the pre-initiation complex (pre-IC) (Fig. 1c). Among the assembled proteins on origins, the heterotetrameric GINS complex, Cdc45, and Mcm2-7 form an active DNA helicase, the Cdc45-Mcm-GINS (CMG) complex (Moyer et al., 2006), which is a core component of the replisome progression complex at the replication forks (Gambus et al., 2006). The unwinding of origin DNA by the CMG complex (Fig. 1d) leads to the initiation of the synthesis of DNA strands by three DNA polymerases, α, δ, and ε (Polα, Polδ and Polε) (Fig. 1e). Polα associates tightly with primase and extends a short DNA strand after the primer RNA synthesized by the primase. Subsequently, the five-subunit clamp loader, replication factor C (RFC), binds to the 3’-OH end of newly synthesized DNA and loads Polδ or Polε with a proliferating cellular nuclear antigen (PCNA, homotrimer sliding clamp that increases Polδ and Polε processivity). At the replication forks, Polδ and Polε synthesize mainly the lagging and leading strands, respectively (Kunkel and Burgers, 2008).
 View Details | Fig. 1 Initiation of chromosomal DNA replication in budding yeast. (a) Formation of the pre-RC. (b) Association of Sld3-Cdc45 with replication origins. (c) CDK-dependent formation of the pre-LC and of the pre-IC. (d) Unwinding of origin DNA. (e) Start of DNA synthesis on unwound DNA. RFA, RFC, and Mcm10 are not depicted. See text for details. |
The genes encoding replication proteins have been identified using biochemical purification of replication proteins and subsequent reverse genetics, as well as genetic screenings. The identification of replication genes and the characterization of their products have elucidated the molecular mechanism of chromosomal DNA replication in eukaryotic cells. In this article, the identification of replication genes and the functions of their products will be described, as well as the rationale behind their isolation using genetic screenings.
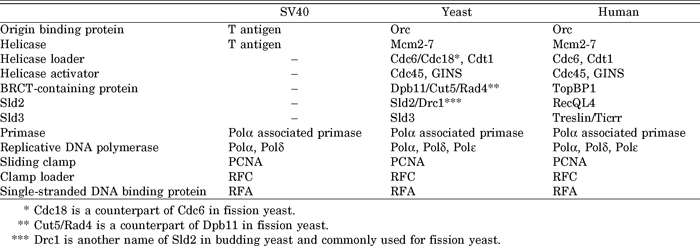
Reverse genetics has contributed mainly to the isolation of the replication genes required for the elongation step of DNA replication. The origin-specific cell-free replication system of the simian virus 40 (SV40) DNA was established in the early 1980s (See for a review Fanning and Zhao, 2009). This system requires the T antigen encoded by SV40 and host cellular factors, such as Polα, Polδ, PCNA, RFC, and replication factor A (RFA, Rfa1-3 complex, which binds to single-stranded DNA) (Table 1). The T antigen binds to the replication origin of the SV40 DNA and unwinds double-stranded DNA as a DNA helicase, and the cellular factors promote DNA synthesis. Counterparts of the cellular factors identified in SV40 replication were purified from yeast cells and their genes were cloned. The subsequent constructions and analyses of conditional mutations in these genes indicated that the cellular factors required for SV40 replication participate in chromosomal DNA replication. Instead of binding to the T antigen, chromosomal replication origins bind to Orc, a complex that is well conserved from yeast to human. This Orc complex was first purified from budding yeast, followed by the identification of the genes encoding its subunits (Bell and Stillman, 1992; Bell et al., 1993). Mutations in Orc subunits conferred a defect of DNA replication in non-permissive conditions, indicating that Orc functions in DNA replication as a cellular factor that binds to replication origins. These analyses shed light on the studies using yeast as a model system for eukaryotic DNA replication.
 View Details | Table 1 Replication proteins required for SV40, yeast and human |
Several yeast genetic screenings have been devised to identify DNA replication genes. In the case of Escherichia coli, DNA synthesis is easily monitored because radioactive deoxythymidine is incorporated specifically into DNA via the salvage pathway. However, laborious procedures were required to measure the synthesis of DNA in yeast cells, which do not have the salvage pathway. Thus, only a few mutations have been isolated by direct screening for mutants defective in DNA replication. Such examples are dna2, dna43 (= mcm10) and dna52 (= dbf4) (Dumas et al., 1982; Kuo et al., 1983; Budd and Campbell, 1995).
Instead of direct measurement of DNA synthesis, yeast screenings used various phenotypes caused by the mutations. Dr. Leland Hartwell’s group first isolated the cell division cycle (CDC) mutants of budding yeast (Hartwell et al., 1970; See for a review Pringle and Hartwell, 1981). In this mutant screen, they focused on the shape of yeast cells and analyzed mutant cells showing uniform shapes when cells were transferred to high temperature. Mutant cells that are defective in DNA replication continue growing and arrest the cell cycle with a large bud at high temperature. According to our current knowledge, this happens because the cells that replicate DNA partially or are exposed to damage caused by aberrant DNA replication arrest their division in G2/M phase via the cell cycle checkpoints (DePamphilis, 2006). Without checkpoints, cellular shapes are heterogeneous when they are arrested. Fission yeast CDC mutations were isolated mainly by Dr. Paul Nurse’s group (Nurse et al., 1976; Nasmyth and Nurse, 1981; Grallert and Nurse, 1996). It is noteworthy that the genes for which mutations were isolated are different in the yeast species, Saccharomyces cerevisiae (budding yeast) and Schizosaccharomyces pombe (fission yeast), both of which are used for extensive isolation of CDC mutants. Cdc2 (a catalytic subunit of Polδ), Cdc6, Cdc7 (Cdc7 kinase: a catalytic subunit of DDK), Cdc9 (DNA ligase), Cdc17 (a catalytic subunit of Polα), Cdc28 (a catalytic subunit of CDK) were identified from S. cerevisiae. Cdc1 (the second largest subunit of Polδ), Cdc2 (a catalytic subunit of CDK), Cdc6 (a catalytic subunit of Polδ), Cdc17 (DNA ligase), Cdc18 (counterpart of Cdc6 of budding yeast), Cdc19 (= Nda1: Mcm2), Cdc20 (a catalytic subunit of Polε), Cdc21 (Mcm4), Cdc23 (Mcm10), Cdc24 (RFC-like protein, an orphan in fission yeast), Cdc27 (the third largest subunit of Polδ), and Cdc30 (Orc1) were isolated from Sch. pombe. Cell shape and hydroxyurea, an inhibitor of DNA replication, were used for mapping their execution points during the cell cycle in budding and fission yeasts. However, the timing of gene function was not always accurate in such experiments. For example, this mapping suggested that Cdc17 of budding yeast works in G2 phase, because leaky mutations in the replication genes caused a partial defect, and the checkpoint arrested the cells in G2 phase, as described before.
Later, similar CDC mutations were isolated from the two yeast species as cold-sensitive alleles or suppressors of known cdc mutations. For example, CDC44 (= RFC1), CDC45, and CDC54 (= MCM4), which encode the largest subunit of RFC, the Cdc45 protein, and the Mcm4 protein, respectively, were isolated as genes responsible for cold-sensitive mutations of budding yeast, and CDC46 (= MCM5) and CDC47 (= MCM7) were isolated as genes responsible for suppressor mutations of the cdc45 mutations (Moir et al., 1982; Hennessy et al., 1991). Suppressor mutations often occur in genes that encode a protein that forms a complex with the original gene product. This was the case, here, as Cdc45 forms a complex with Mcm2-7 (Fig. 1b). However, their execution points were mapped in G2 phase (Moir and Botstein, 1982).
Similar mutations not named CDC were also isolated, including Dumbbell former (DBF) mutations. Dbf4 is a regulatory subunit of DDK of budding yeast (Johnston and Thomas, 1982).
Instead of cellular shape, Dr. Bik Tye’s group screened for minichromosome maintenance (mcm) mutations that reduce the stability of the plasmid (minichromosome), which has a centromere and a replication origin in budding yeast (Maine et al., 1984; See for a review Tye, 1999a). They isolated many mutations that confer defect of chromosome segregation or DNA replication. The latter mutations occurred in the genes encoding the Mcm2, Mcm3 and Mcm5 subunits of the Mcm2-7 complex, and Mcm10. This screening was expected to isolate leaky mutations occurring in the initiator proteins, as the tight mutations might not allow cell growth. Moreover, chromosomes have multiple replication origins, whereas minichromosomes have a single origin. This might also alleviate the growth defect caused by the mutations. Furthermore, the mcm mutations occurring in replication genes showed origin specificity; the plasmid bearing one origin is less stable than that bearing others in the mutant cells. This characteristic might also help the isolation of the mutations. This screening itself suggested that these proteins participate in the initiation step of chromosomal DNA replication.
Analysis of the Mcm5 (Cdc46) protein showed that it is localized in the nucleus from late M to G1 phase (Hennessy et al., 1990; Chen et al., 1992). This observation suggested that it would be a licensing factor, and highlighted the role of the Mcm proteins. The licensing factor model for the control of DNA replication was proposed by Drs. Julian Blow and Ron Laskey based on the study of DNA replication in Xenopus egg extracts (Blow and Laskey, 1988; Blow, 1993). In this model, DNA replicates only once after encapsulation of the chromosome by the nuclear membrane, because a putative licensing factor that is essential for the initiation and does not penetrate the nuclear membrane binds to chromatin during mitosis, a point at which the nuclear membrane is broken down. Mcm2-7 and Cdt1 were purified as licensing factors although they localize in the nucleus throughout the cell cycle (Chong et al., 1995; Kubota et al., 1995; Tada et al., 2001). In yeast, the pre-RC was proposed by observation of in vivo footprinting and was suggested as a licensing factor (Diffley et al., 1994). The pre-RC was later shown to contain Orc and Mcm2-7 (DePamphilis, 2006).
Three cycles of mcm screenings identified four replication genes, MCM2, MCM3, MCM5 and MCM10 (Tye, 1999b). However, other genes participating in the initiation step were not identified. As even the genes encoding other subunits of the Mcm2-7 complex were not identified, the mcm screening has not been saturated. I speculate on this subject as follows. A defective elongation step affects chromosomal DNA replication more than that of small minichromosomes (plasmids), because the intact chromosome requires longer DNA replication. Thus, the identification of mutations in the components of the replication forks is difficult. This suggests that mutations in the pre-RC and part of the pre-IC will be detected as mcm if they do not affect the function of the replication fork. However, mutations of Orc, which is a component of the pre-RC, were not identified as mcm. Mutations of subunits of Orc compromise not only the initiation of chromosomal DNA replication, but also silencing of the mating loci, which maintains the mating ability of haploid yeast cells (Foss et al., 1993). Thus, mutations of Orc are not amenable to genetic analysis and may be excluded from the analyses.
The synthesis of the SV40 DNA requires Polα and Polδ. These two DNA polymerases also function as replicative DNA polymerases for chromosomal DNA replication in budding yeast. The third replicative DNA polymerase, Polε, was first purified from budding yeast (Hamatake et al., 1990; Pursell and Kunkel, 2008). This polymerase consists of four subunits, Pol2 (256 kDa), Dpb2 (78 kDa), Dpb3 (23 kDa), and Dpb4 (22 kDa); Pol2 is the catalytic subunit. The genes encoding the subunits of this polymerase were cloned using antibodies and the amino acid sequence of purified Polε. The subsequent disruption of these genes revealed that the Pol2 and Dpb2 subunits are essential for cell growth (Morrison et al., 1990; Araki et al., 1991a), whereas Dpb3 and Dpb4 are dispensable (Araki et al., 1991b; Ohya et al., 2000). Moreover, the conditional mutations of POL2 and DPB2 compromise DNA replication in non-permissive conditions (Araki et al., 1991a, 1992). Surprisingly, although point mutations in the domain of Pol2 that is responsible for polymerase activity abolish cell growth (Araki et al., 1992; Dua et al., 1999), deletion of the same domain supports cell growth (Dua et al., 1999; Kesti et al., 1999). This is probably caused by compensation of Polε polymerase activity by Polδ. The C-terminal half of Pol2, which lacks any enzymatic activity, is essential for chromosomal DNA replication (Dua et al., 1999; Kesti et al., 1999).
We isolated the DPB11 gene as a multicopy suppressor of the dpb2-1 temperature-sensitive allele and found that multicopy DPB11 suppresses the pol2-11 and pol2-12 temperature-sensitive mutations (Araki et al., 1995). Polε purified from cells bearing the dpb2-1 mutation has trace amounts of the Dpb2 subunit (Araki et al., 1991a). The pol2-11 and pol2-12 mutations lack 32 and 28 amino acid residues from the C terminus of Pol2, respectively, and reduce the interaction between Pol2 and Dpb2 (Dua et al., 1998). Thus, Dpb11 was suggested as having a function that is closely related to the formation of the intact complex of Polε. Moreover, mutations occurring in essential genes are often suppressed by increased dosage (multicopy) of the related genes and cannot be combined with mutations in related genes. The dpb11-1 temperature sensitive allele cannot be combined with temperature-sensitive mutations in essential subunits of Polε (pol2-11, pol2-12, pol2-18 and dpb2-1), even in permissive conditions (Araki et al., 1995), or with deletions of nonessential subunits of Polε, Dpb3, and Dpb4 (Ohya et al., 2000).
Dpb11 is essential for cell growth and plays a dual role in chromosomal DNA replication and at the cell cycle checkpoints (Araki et al., 1995). The Dpb11 protein has two pairs of tandem BRCT repeats (for a total of four BRCT repeats), which bind to phosphopeptides (Glover et al., 2004; Garcia et al., 2005). The tandem BRCT repeats bind to two replication proteins, Sld2 and Sld3, and two checkpoint proteins, Rad9 and Ddc1 (Masumoto et al., 2002; Wang and Elledge, 2002; Tanaka et al., 2007b; Zegerman and Diffley, 2007; Granata et al., 2010). The C-terminal stretch located after the fourth BRCT repeat contains the activation sequence for a pivotal checkpoint kinase, Mec1, which is the counterpart of the ATR kinase of human cells (Mordes et al., 2008; Navadgi-Patil and Burgers, 2008).
The Dpb11 protein and its counterparts in other organisms, such as Cut5/Rad4 of fission yeast, Mus101 of Drosophila, and TopBP1 of human cells, constitute a protein family bearing multiple BRCT domains and function in DNA replication and at the cell cycle checkpoints (Garcia et al., 2005).
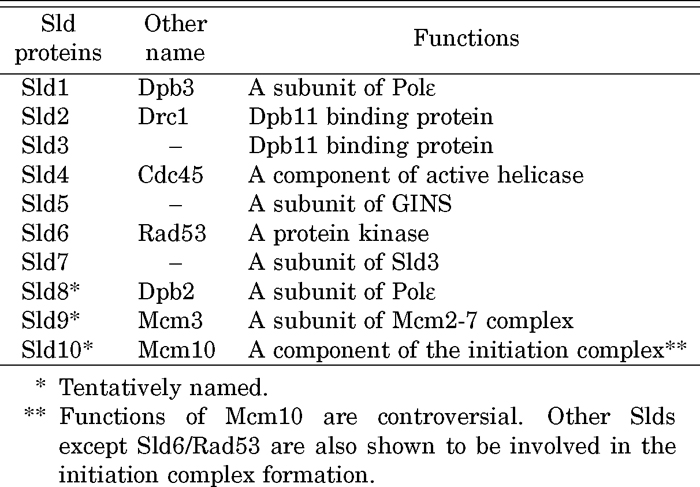
Although Dpb11 is essential for DNA replication, its role had not been elucidated. Thus, we isolated the synthetic lethal with dpb11 (sld) mutations (Kamimura et al., 1998; Tanaka et al., 2011). Two cycle screenings for sld mutations identified 16 mutations occurring in 11 genes (Table 2). Later studies revealed that the Sld proteins function in the formation of the pre-IC, together with Dpb11 and Polε (Fig. 1c).
 View Details | Table 2 Sld proteins and their functions |
The first screening used the dpb11-1 mutation occurring at the boundary between the fourth BRCT repeat and the C-terminal stretch and found 10 mutations in SLD1, -2, -3, -4, -5 and -6 genes. The sld1 mutation occurred in the third-largest subunit of Polε, Dpb3. The sld2 and sld3 mutations occurred in previously unidentified genes. Sld2 and Sld3 bind to the C-terminal and N-terminal tandem BRCT repeats of Dpb11, respectively, after phosphorylation by CDK (Masumoto et al., 2002; Tak et al., 2006; Tanaka et al., 2007b; Zegerman and Diffley, 2007). These phosphorylation-dependent interactions are essential and represent the minimal requirement for the CDK-dependent activation of DNA replication (Fig. 1c) (Tanaka et al., 2007a, 2007b; Zegerman and Diffley, 2007; Tanaka and Araki, 2010). sld4 is a mutation occurring in Cdc45, which is required for the initiation and elongation steps of DNA replication. Sld3 and Sld4 (Cdc45) form a complex throughout the cell cycle that associates with the pre-RC-formed origins in a mutually dependent manner (Fig. 1b). This association occurs with early-firing origins in G1 and with late-firing origins in S phase (Kamimura et al., 2001). In addition, Cdc45, but not Sld3, moves with the replication forks (Fig. 1d) (Kanemaki and Labib, 2006). The sld5 mutation also occurred in a previously unidentified gene. A combination of genetic and biochemical analyses identified a four-subunit replication factor, GINS, comprising Sld5 and partner of sld five (Psf) 1, 2, and 3. GINS was named after the number-based names of the genes Go, Ichi, Ni, and San; five, one, two, and three in Japanese. GINS functions in the initiation and elongation of chromosomal DNA replication (Takayama et al., 2003). Cdc45 and GINS associate with Mcm to form an active replicative helicase, the CMG complex (Fig. 1d) (Moyer et al., 2006). The genes encoding subunits of GINS were also isolated in the genome-wide screening for cell cycle genes using the temperature-degron system (Kanemaki et al., 2003). The sld6 mutation occurred in Rad53 kinase, which functions at checkpoints and in the regulation of the initiation of DNA replication (Dohrmann and Sclafani, 2006).
The second screening for sld mutations used the dpb11-24 mutation occurring in the first BRCT repeat and identified six mutations in five genes. Four of the five genes were known as SLD3, DPB2, MCM3, and MCM10. The remaining gene was the newly identified SLD7. The Sld7 protein forms a tight complex with Sld3 throughout the cell cycle; Sld3 and Sld7 are subunits of a complex. However, although Sld7 is not essential for cell growth, lack of this protein slows DNA replication and cell growth. This protein seems to promote the efficient separation of the replication machinery from origins (Fig. 1d) (Tanaka et al., 2011).
The repertoire of genes identified in the two screenings differed. This was probably caused by the dpb11 mutations used in the screenings. Interestingly, the most popular genes identified are related to the defect caused by dpb11 mutations. SLD2 (5/10) and SLD3 (2/6) were the most popular in the first and second screenings, respectively. The first screening used the dpb11-1 mutation, which reduces the interaction between Dpb11 and Sld2 (Masumoto et al., 2002), and the second screening used the dpb11-24 mutation, which reduces the interaction between Dpb11 and Sld3 (Tanaka et al., 2007b).
Synthetic-lethal screenings often identify genes that encode proteins that interact directly or form a complex. The Sld proteins are an example of this observation. The analysis of the physical interactions between the proteins during the cell cycle led to the identification of the pre-loading complex (pre-LC) containing Polε, GINS, Dpb11, and Sld2 (Fig. 1c) (Muramatsu et al., 2010). Moreover, purified these four proteins form a complex. Thus, the minimum core factors of the pre-LC seems to be Polε, GINS, Dpb11, and Sld2. Formation of the pre-LC depends on the phosphorylation of Sld2 by CDK, both in vitro and in vivo, but not on the pre-RC. Polε and GINS form a fragile complex throughout the cell cycle, which associates with the Sld2-Dpb11 complex. GINS does not associate with the Sld2-Dpb11 complex in the absence of Polε, suggesting that Polε mediates the interaction between GINS and Sld2-Dpb11. Although Sld2 binds to Polε, GINS does not bind to Sld2 efficiently, even in the presence of Polε. Thus, GINS requires the interaction between Sld2 and Dpb11 for stable association.
The pre-LC complex explains why mutations of Sld1 (Dpb3 subunit of Polε), Sld2, and Sld5 (a subunit of GINS) were isolated as a synthetic lethality with dpb11-1; Dpb11-1 partially disrupts the interaction between Dpb11 and Sld2 and, thus, the formation of the pre-LC. In addition, combination of this mutation with the sld mutation, which also disrupts partially the formation of the pre-LC, abolishes the formation of the pre-LC completely.
We proposed that the pre-LC is recruited to replication origins via an interaction between the N-terminal tandem BRCT repeats of Dpb11 in the pre-LC and phosphorylated Sld3 (Fig. 1c). The Sld3-Sld7 complex and Sld4 (Cdc45) associate with pre-RC-formed origins in a mutually dependent manner. Thus, these Slds are required for the association of Dpb11 with origins; thus, they are closely related to Dpb11 function. The Mcm2-7 complex, a component of the pre-RC, binds directly to the Sld3-Sld7 complex (our unpublished result) and Cdc45; thus, the mcm3 mutation might be isolated as sld. Although the function of another Sld, Mcm10, remains controversial, it may associate with origins, probably as a component of the pre-IC (Gambus et al., 2006). Thus, Mcm10 is also closely related to Dpb11 function. We further speculate that the pre-LC functions as a carrier of GINS to the pre-RC to form the CMG complex, which is thought as an active replicative DNA helicase (Muramatsu et al., 2010).
CDK is a key regulator of the cell cycle in eukaryotic cells. It regulates the initiation of DNA replication (DePamphilis, 2006). CDK phosphorylates Orc, the Mcm2-7 complex, Cdc6, and Cdt1. CDK phosphorylation reduces the interaction between Orc and Cdt1 (Chen and Bell, 2011), excludes the free Mcm2-7 complex from the nucleus, and enhances the degradation of Cdc6. These phosphorylation events prevent the formation of the pre-RC and consequently reinitiation (DePamphilis, 2006). Conversely, CDK phosphorylates Sld2 and Sld3, thus enhancing the formation of the pre-LC and its recruitment to replication origins. If this process is bypassed, DNA replication initiates in the absence of CDK activity. Therefore, CDK turns on S-phase by forming the pre-LC and recruiting it to origins (Tanaka and Araki, 2010). Therefore, genetic screening for sld mutations identified all the components that participate in the CDK-regulated initiation of chromosomal DNA replication.
With the expectation of functional elucidation of Polε at the elongation step in DNA replication, we isolated DPB11 as a multicopy suppressor of mutations of subunits of Polε and further obtained SLDs as causal genes of synthetic-lethal mutations with dpb11. However, most of these factors were involved in the initiation of chromosomal DNA replication. Interaction analysis of the proteins revealed that Polε functions in the initiation of DNA replication as a component of the pre-LC, in addition to its function in the synthesis of the leading strand at the replication fork.
The essential function of Polε in the initiation step of DNA replication is consistent with the requirement of SV40 replication (Zlotkin et al., 1996; Pospiech et al., 1999). In vitro SV40 DNA replication does not require the CMG helicase, because SV40 encodes its own DNA helicase, the T antigen. This implies that this replication system does not require the factors that function in the formation of the CMG helicase, the pre-LC and the related proteins, Cdc45 and Treslin/Ticrr (Kumagai et al., 2010; Sanchez-Pulido et al., 2010; Sansam et al., 2010), which is a counterpart of Sld3 in human cells. Thus, none of the components of the pre-LC, Polε, GINS, TopBP1 (a counterpart of Dpb11) (Garcia et al., 2005) and recQL4 (a counterpart of Sld2) (Sangrithi et al., 2005; Matsuno et al., 2006) are required for SV40 replication (Table 1).
A function of Polε distinct from other replicative polymerases is also predicted from the evolution of DNA polymerases. Based on comparative analysis of the amino acid sequences of DNA polymerases, Tahirov et al. (2009) suggested that Polε and two other replicative polymerases, Polα and Polδ, originate from distantly related ancestral archaeon B-family DNA polymerases. Moreover, these authors revealed that the catalytic subunit of Polε consists of two tandem exonuclease-polymerase modules, the active N-terminal module and a C-terminal module, in which enzymatic domains are inactivated. According to their proposal, Polε evolved as a result of the insertion and subsequent inactivation of a distinct polymerase domain, probably from a DNA polymerase of an α-proteobacterium (proto-mitochondrion). These authors suggest further that the inactivated polymerase-exonuclease module of Polε may play an important role in the assembly of replication and repair complex. In DNA replication, Polε plays a role in the assembly of the pre-LC (Araki, 2010).
In addition to reverse genetics, classical genetic screenings identified many replication genes in yeasts and accelerated the elucidation of eukaryotic chromosomal DNA replication. Moreover, the genetic interactions used in screenings and analyses provide insight into the molecular mechanism of chromosomal DNA replication. The combination of genetic and biochemical analyses has revealed the molecular mechanism of DNA replication. Although it is widely believed that all proteins required for chromosomal DNA replication in budding yeast are already known, the genetic tools used remain important, as described below.
The study of eukaryotic DNA replication itself is moving toward more mechanistic approaches. We can now purify replication proteins and combine them in vitro to analyze the various steps of DNA replication. Moreover, a cell-free system of chromosomal DNA replication of budding yeast has been reported very recently (Heller et al., 2011), indicating that it will be possible to analyze the molecular mechanism of DNA replication more directly. Thus, we now envisage that most of the molecular mechanism of eukaryotic DNA replication will be illustrated in the next decade. However, cell-free systems often give rise to artifacts. We need to confirm the mechanism proposed by in vitro analyses using genetic tools; mutations that compromise specific steps must be used for this purpose.
Cell-free systems are not almighty. Eukaryotic cells have many replication origins on their chromosomes. Some origins fire stochastically early or late in S phase and some fire with predetermined timing. Although we do not know what decides the timing of origin firing during S phase, a higher-ordered nucleosome structure or the nuclear environment of replication origins may be important and cannot be assessed using a cell-free system. Thus, genetic approaches remain promising; mutations that abolish this timing of firing must be very useful, although no such mutation has been reported.
There are also many unsolved problems related to DNA replication in eukaryotic cells. Replication forks that face obstacles, DNA damage or shortage of DNA precursors, stall. Replication proteins and checkpoint factors work coordinately at the stalled forks. For example, Dpb11 associates with replication origins to initiate DNA replication and dissociates from origins when DNA synthesis starts. However, it is proposed that, when replication forks stall, Dpb11 associates with the forks via unknown mechanisms, to manage these problems (Granata et al., 2010). How these factors and structures work together is a future project in this field. In addition, the mechanism via which chromosomes duplicate during DNA replication remains unknown. Chromosomal DNA associates with various proteins and forms chromatin; histones form nucleosomes, which wrap the chromosomal DNA, and many regulatory proteins, such as transcription and epigenetic-related factors, bind to DNA or the nucleosomal histones. A cell-free system of DNA replication will be a useful tool to analyze this phenomenon and the genetic approach will facilitate this analysis.
Yeast is a good model for the study of DNA replication in eukaryotes. Replication proteins, especially machineries such as DNA helicase and polymerases, are well conserved from yeast to higher eukaryotes (Table 1). However, higher eukaryotes probably have a mechanism that is modified from that of the simple eukaryotes, yeasts, which may be connected to regulatory mechanisms acquired during evolution. These modifications will be addressed further in the near future.
I thank the members of my laboratory for helpful discussions, and Drs. K. Hizume, M. Kanemaki, and S. Tanaka for comments on this manuscript.
|