| Edited by Fujio Kawamura. Kouji Matsumoto: Corresponding author. E-mail: koumatsu@mail.saitama-u.ac.jp |
Glycolipids (GL) appear in a remarkable variety of contexts in Gram-positive bacteria, cyanobacteria, and the chloroplasts of plant cells. Galactolipids play an important role in photosynthesis in cyanobacteria; the crystal structure of the photosystem I (PSI) complex from the thermophilic cyanobacterium Synechococcus elongatus reveals that its PSI contains one molecule of monogalactosyldiacylglycerol (MGalDG) (Jordan et al., 2001). In Arabidopsis, MGalDG is required for thylakoid biogenesis, photosynthesis and embryogenesis (Kobayashi et al., 2007). On the other hand, the viability of Staphylococcus aureus mutant cells lacking GL is reduced in phosphate-saline and their autolysis in the presence of Triton X-100 is faster than that of cells of the parental strain (Kiriukhin et al., 2001).
The Bacillus subtilis Marburg 168 cell contains three kinds of GL, monoglucosyldiacylglycerol, diglucosyldiacylglycerol (DGDG), and triglucosyldiacylglycerol, which amount to 1.2%, 9.8% and 0.3% of total membrane lipids (Kawai et al., 2006). These are synthesized processively by UgtP, which transfers glucose to diacylglycerol from UDP-glucose (Jorasch et al., 1998). The phenotype of cells with ugtP (ypfP) disruption has been reported to be shorter and rounder for the first time by Price et al. (1997). The ugtP mutant cells have also been reported to show strikingly abnormal cell morphology (Lazarevic et al., 2005). In addition, we have shown that the GFP fusion of UgtP, which is predicted to be a cytoplasmic protein with no transmembrane segment, is localized in the septal region in the form of an open-ring similar to the FtsZ-ring (Nishibori et al., 2005). Recently, Weart et al. (2007) have proposed that UgtP regulates the timing of cell division by adjusting the assembly of FtsZ protein based on their finding of dissociation in vitro of FtsZ polymer in the presence of UgtP.
In this study, we characterize B. subtilis mutant cells lacking GL. We describe a morphology of ugtP mutant cells that differs from what has been reported previously, and demonstrate the activation of three extracytoplasmic function sigmas (SigM, SigV and SigX) and of SigI. Furthermore, we show that the transcription of the genes of two mreB isoforms is increased, whereas GFP-MreB protein is reduced in the ugtP mutant cells.
The B. subtilis strains used in this study are listed in Table 1. All the strains used in this study were constructed by introduction of mutations of other strains into B. subtilis Marburg 168 by transformation.
 View Details | Table 1 B. subtilis strains used in this study |
Bacterial strains were grown aerobically in LB medium with shaking at appropriate temperature (37°C). Cell growth was monitored with a TAITEC mini photo 518R (at OD530) or Shimadzu Bio Spec-mini (at OD600). Cells of mreB and mbl derivatives were cultivated with supplementation of 10 mM MgSO4. Transformation of B. subtilis was carried out basically by the method of Spizizen (Anagnostopoulos and Spizizen, 1961). Transformation of E. coli and DNA manipulation were performed as described previously (Matsuoka et al., 2007). The following antibiotics were added to the media: for selection of B. subtilis transformants, chloramphenicol (5 μg/ml), kanamycin (5 μg/ml or 15 μg/ml for only mreB derivatives), neomycin (5 μg/ml), and spectinomycin (100 μg/ml).
After collection at room temperature, cells were centrifuged and resuspended in one tenth volume of LB medium. Then the cells were fixed on object slides coated with a layer of 1% (wt/vol) agarose gel in water. Fluorescence images were viewed with an ECLIPSE E600 fluorescence microscope (Nikon) and a Cooled CCD camera (ORCA-ER; Hamamatsu Photonics Co., Hamamatsu Japan). Green fluorescence from GFP (excitation at 495 nm; emission at 525 nm) was detected by using a standard GFP(R)-BP filter unit (460–500 nm excitation and 510–560 nm emission). To minimize the toxicity of high-energy excitation light, the focus was set under phase-contrast conditions and the fluorescence images were captured shortly after the shift to high-energy excitation light. The exposure time for green fluorescence of GFP was 2 sec. Captured images were processed with Adobe Photoshop 6.0. To measure cell length and width, cell membranes were stained with 1 μg/ml FM4–64 (Molecular probes, USA). The length was defined as the length of a center line which connects from a cell pole (or center of septum) to another on the picture. The width was measured at the septum. The values were converted to actual length using the micrometer on the microscope as a standard.
To measure transcriptional activities of ECF sigmas and SigI, B. subtilis cells were transformed with DNAs extracted from BSU15 (PsigI’-bgaB), BSU31 (PsigM’-lacZ), BSU32 (PsigV’-lacZ), and BSU34 (PsigX’-lacZ). To measure the promoter activities of mreB isoforms, strains were constructed as follows: the chromosomal DNAs were extracted from ASK4603 (PmreB’-lacZ), ASK4604 (Pmbl’-lacZ), ASK4605 (PmreBH’-lacZ), and ASK4606 (Pupstream mreB’-lacZ), these DNAs of lacZ fusions were then introduced into appropriate strains by transformation. LacZ was assayed as described previously (Miller, 1972). The BgaB assay was performed at 62°C.
Sodium dodecyl sulfate (SDS)-PAGE was performed with 10% polyacrylamide by the method described previously (Laemmli, 1970). Western blotting was performed by using anti-GFP antibody (Santa Cruz Technology) and anti-mouse antibody conjugated with horseradish peroxidase (HRP) (GE Healthcare) or anti-SigA antibody (Yee et al., 2011) and anti-rabbit antibody conjugated with HRP (Jackson ImmunoResearch Inc.). After electrophoresis, the gel was soaked in transfer buffer (25 mM Tris, 192 mM glycine, 10% methanol). Proteins in the gel were transferred onto a polyvinylidene difluoride membrane (GE Healthcare) using the semidry blotting method and then neutralized with phosphate saline buffer (PBS). Skim milk (1%) in PBS was used for blocking at room temperature for 1 h, and then anti-GFP antibody was added (1:10,000), and the mixture was incubated at room temperature for 2 h. The membrane was washed with PBS for 5 min three times. After washing, anti-mouse antibody conjugated HRP was added (1:10,000), and the mixture was incubated at room temperature for 2 h. To detect the signal, ECL-plus (GE Healthcare) was used. The signals were exposed on X-ray film (Fuji film) for appropriate times. The same membrane was used for detection of SigA after stripping the antibodies. To strip antibodies, the membrane was incubated into strip buffer (67.5 mM Tris-HCl pH 6.8, 2% SDS, and 50 mM 2-mercaptoethanol) with shaking at 50°C. Then the membrane was washed using TBS-T twice at room temperature.
The product of ugtP (ypfP) in B. subtilis has been identified as a UDP-glucose:diacylglycerol glucosyltransferase (Jorasch et al., 1998). Previously, ugtP mutant cells have already been reported to be shorter and rounder than the wild type (Price et al., 1997). However, we observed a different morphology. Both ugtP mutant strains, the one obtained by insertion of a pMUTIN plasmid (strain BFS2221) as well as the other obtained by replacement with a kan cassette (strain YTB019) (Fig. 1A), were quite bent out of shape and thicker during the logarithmic growth phase (OD530 at 0.3) and shorter and rounder only in the stationary phase (OD530 at 1.3). The length and width of YTB019 (ugtP::kan) cells, measured using the built-in micrometer on the microscope, in the mid-logarithmic growth phase (OD530 at 0.3), were obviously thicker and shorter than the wild type cells (Fig. 1B). Since UDP-glucose is needed as a glucose donor for the synthesis of GL, we examined the morphology of mutant strains in which a gene (pgcA or gtaB) involved in UDP-glucose synthesis was disrupted. Lack of GL in these disruptant cells was confirmed by thin layer chromatography followed by detection with orcinol reagent spray. These mutant cells were similar in shape to ugtP mutant cells (data not shown). The bent and distended cell phenotype of the ugtP mutants therefore, seems to be caused by the lack of GL.
 View Details | Fig. 1 Characteristics of the ugtP disruptant. (A) Morphology of ugtP mutant cells. The ugtP mutant cells of strain YTB019 (ugtP::kan) show distended and bent shapes during logarithmic growth, and are shorter in the stationary phase. The black bar indicates 10 μm. (B) Comparison of cell length and width of wild type with GL-lacking cells. Cells of YTB019 (ugtP::kan) were grown in LB to OD530 = 0.3 and cell length and width were measured (n = 100). WT: circle, ugtP: triangle. |
Reduction in the content of phosphatidylglycerol in the membranes affects the activities of extracytoplasmic function (ECF) sigmas, which are concerned with cell surface maintenance and stress response, and are tethered by their cognate anti-sigma factors to the membranes in non-stress conditions (Hashimoto et al., 2009). We expected therefore that some changes would occur in the membranes of cells lacking GL, and focused on the extracytoplasmic function sigmas. Examination of the levels of all ECF sigmas in GL lacking cells revealed that σM, σV and σX were activated (data not shown). The activation of σM attracted our interest, because the σM regulon is involved in cell wall synthesis and cell shape maintenance (Cao et al., 2002, Salzberg and Helmann, 2008), and continuous activation of σM is toxic to cells (Asai, K., et al., personal communication). Therefore, we disrupted the sigM gene of the ugtP::kan strain to see whether its phenotype resulted from continuous activation of σM. Unexpectedly, the phenotype of the constructed double mutant was even more degenerate, and the cells were shaped like balloons (Fig. 2), a shape that had been described as the phenotype of a ugtP mutant previously (Lazarevic et al., 2005). Disruption of the other ECF sigma genes, which were activated in GL lacking cells did not have a negative effect on cell shape (Fig. 2). The results indicate that the abnormal morphology of ugtP mutant cells was not caused by the activation of ECF sigmas, but a consequence of the change of membrane lipid composition because of lack of GL, to which ECF sigmas responded.
 View Details | Fig. 2 Morphology of double mutants of ugtP with defective mutations of ECF sigma genes. All strains were grown in LB medium to OD530 = 0.3. Left side panels show disruptants of single ECF sigmas. The right side shows double disruptants in which the defective mutation of one ECF sigma is coupled with the ugtP mutation. |
When they were grown in LB medium containing 1% sodium chloride, the shape of the ugtP sigM double mutant cells changed and became much more spherical (Fig. 3A). We further examined the characteristics of the double mutant and found that the ugtP sigM double disruptant was very sensitive to sodium chloride. The double mutant could not grow on LB agar plates containing 2% sodium chloride (Fig. 3B), in contrast to the ugtP and sigM single mutants, which show the same level of resistance to sodium chloride as the wild type. In the liquid medium the double mutant lysed during the stationary phase when the medium contained 1.5% sodium chloride (Fig. 3C).
 View Details | Fig. 3 Characteristics of the ugtP sigM double mutant. (A) Morphology of the ugtP sigM double mutant (strain MBS30) in LB medium containing 1% sodium chloride. Cells were grown to OD530 = 1.0 then harvested and observed under a phase contrast microscope. The black bar indicates 5 μm. (B) Growth of the ugtP sigM double mutant (strain MBS30) on LB agar plates containing 1%, 1.5%, and 2% sodium chloride. Overnight cultures in LB medium containing 1% sodium chloride were diluted tenfold serially down from 10–2 to 10–6 with saline. 3 μl each was spotted on LB agar plates containing 1%, 1.5%, and 2% sodium chloride. These plates were then incubated overnight at 37°C. (C) Growth of the ugtP sigM double mutant on LB medium containing 0.5%, 1%, and 1.5% sodium chloride. Overnight cultures of each strain were inoculated adjusting to OD600 = 0.05 in LB medium containing 0.5%, 1%, and 1.5% sodium chloride and cultivated at 37°C for 6 h. |
The abnormal morphology of the ugtP mutant was suppressed by addition of magnesium or manganese ions (Fig. 4). A similar observation (suppression in the presence of 1 mM MgSO4) has been previously reported for a ugtP single mutant (Lazarevic et al., 2005). This phenomenon reminded us of the characteristics of the mutants of mreB isoforms. MreB is one of the three MreB isoforms in B. subtilis and is important for maintenance of its rod shape and the other two, MreBH and Mbl, are also significant for the maintenance of the cell shape (Kawai et al., 2009b). Thus, we inspected the expression levels in the ugtP mutant cells of the genes for the isoforms via lacZ transcriptional fusions integrated at the amyE locus. The mreB gene is transcribed from two promoters, one is located at an internal site of maf (Jervis et al., 2007), designated here as Pupstream mreB, and the other is just on mreBCD, PmreB (Fig. 5A). The mbl and mreBH genes are monocistronic and the latter is transcribed from the promoter recognized by SigI (Tseng and Shaw, 2008). The LacZ reporter assays revealed that the transcription levels of mreB (Pupstream mreB) and PmreBH were ca.4.3- and 2.3 times as high as those in ugtP+ cells, respectively, but the others (PmreB and Pmbl) were unchanged in the ugtP mutant cells (Fig. 5B). Furthermore, we constructed double mutants having ugtP and one of the isoforms. A balloon-like form similar to that of ugtP sigM was observed with the ugtP mreB mutant cells, but not with the other double mutant cells (Fig. 6).
 View Details | Fig. 4 Suppression of abnormal morphology of ugtP mutant by addition of magnesium and manganese. YTB019 cells were grown in LB medium without addition (left panel), with 10 mM MgSO4 (middle), or 0.6 mM MnCl2 (right). Cells were harvested at OD530 = 0.7 and subjected to microscopic observation. The black bar indicates 5 μm. |
 View Details | Fig. 5 Transcriptional activities of mreB isoforms in ugtP disruptant cells. (A) An illustration of neighboring genes of the mreB operon and promoters. The operon is transcribed from two promoters, Pupstream mreB and PmreB. (B) Transcriptional activities of mreB isoforms in ugtP cells. Cells were grown in LB medium to OD530 = 0.5. Cells were harvested and LacZ assays were performed by the method described in MATERIALS AND METHODS. Open bar indicates WT cells, filled bar ugtP mutant cells. Results are average values with S.D. from three independent cultures. |
 View Details | Fig. 6 Morphology of double mutants of ugtP coupled with a defective mutation of one of the mreB isoform genes. Overnight cultures of each strain were inoculated in LB medium, adjusted to 1/100 volume, then grown to OD530 = 0.5. Cells were harvested and observed by phase contrast microscope. |
Then we examined the localization of MreB in the cells lacking GL using the expression of GFP-MreB. To observe GFP-MreB, mreB on the chromosome was replaced with a kan cassette, and then Pxyl-gfp-mreB was introduced at the amyE locus. MreB has been believed to localize helically along the cell axis with MreBH and Mbl in wild type cells (Graumann, 2007). This apparently helical pattern has recently been shown to stem from a misinterpretation owing to the increased depth of field (Garner et al., 2011; Domínguez-Escobar et al., 2011). Discrete dots (in an apparently helical pattern) of GFP-MreB were observed in both ugtP+ and GL lacking cells at the beginning of the log phase (OD530 at 0.2), but at mid-log phase (OD530 at 0.5), such localization was not observed anymore and faint dots of fluorescence were scattered all over whole cells. (Fig. 7A). Actually, a western blotting experiment showed that the amount of GFP-MreB was considerably decreased, as seen in Fig. 7B. The results indicate that GFP-MreB was labile in the cells lacking GL. We suggest that GL is important for the localization of MreB especially during the mid-log phase.
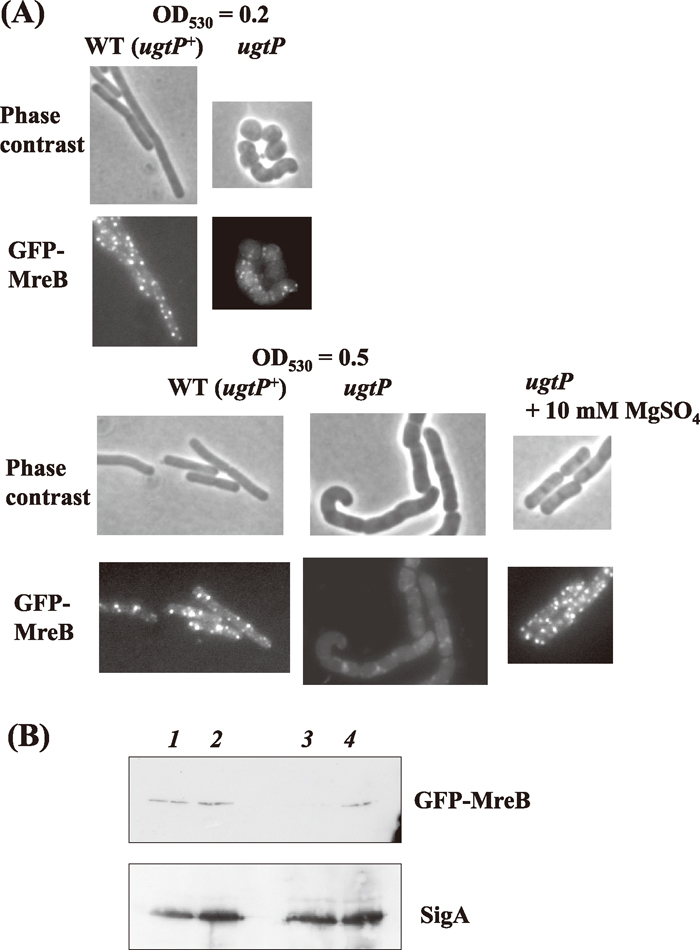 View Details | Fig. 7 Abnormal localization of MreB in cells with ugtP mutation. (A) Localization of GFP-MreB in MBS42 (ugtP+) and MBS43 (ugtP) mutant cells. Cells were grown in LB medium containing 0.01% xylose and were harvested at OD530 = 0.2 and OD530 = 0.5, then observed under a phase contrast microscope. (B) Detection of GFP-MreB by western blotting. MBS42 (ugtP+) and MBS43 (ugtP) mutant cells were grown in LB medium containing 0.01% xylose, and harvested at OD530 = 0.2 and 0.5. Western blotting was performed by the method described in MATERIALS AND METHODS. In the upper panel, signals of GFP-MreB protein were detected with α-GFP antibody, and in the lower panel signals were then detected by reprobing with α-SigA antibody (internal control). Whole cell lysates were used [MBS43 (ugtP), lanes 1 and 3; MBS42 (ugtP+), lanes 2 and 4],which were harvested at OD530 = 0.2 (lanes 1 and 2) and at OD530 = 0.5 (lanes 3 and 4). |
As shown above, expression of mreBH was activated in the ugtP mutant cells (Fig. 5B). The promoter of mreBH has been reported to be recognized by SigI (Tseng and Shaw, 2008). We thus checked the promoter activity of sigI in the cells of ugtP background. As shown in Fig. 8A, there was no difference in the promoter activity of sigI transcription levels between the ugtP mutant and the wild type during both the log and the stationary phase. The transcription level of mreBH in ugtP and sigI mutant cells was then examined. The level of mreBH was 2.1-fold higher in the ugtP mutant than in the wild type (Fig. 8B). After disruption of sigI, no transcription of mreBH could be detected (i.e., the mreBH transcription was dependent solely on SigI), suggesting that SigI was activated by a post-translational mechanism or that an unknown transcriptional regulator acts on mreBH expression in GL lacking cells, although the mechanism of SigI activation is unclear.
 View Details | Fig. 8 Activities of PsigI and PmreBH in ugtP and sigI mutant cells. LacZ assays were performed by the method described in MATERIALS AND METHODS. Results are average values with S.D. of three independent cultures. (A) PsigI activity in WT and ugtP mutant cells. Cells of the strains harboring the transcriptional fusion PsigI-bgaB were grown in LB medium. Cells were harvested at OD530 = 0.3 (log phase) and OD530 = 1 (stationary phase), and LacZ activity was assayed. (B) Cells of the strains, with ugtP, sigI, and ugtP sigI mutations, harboring the transcriptional fusion PmreBH-lacZ were grown in LB medium. Cells were harvested at OD530 = 0.5 and LacZ activity was assayed. |
A study by Weart et al. (2007) indicated that UgtP inhibits FtsZ assembly in vitro, and it was proposed that UgtP regulates FtsZ assembly, but our data do not support that proposal. It is possible that the difference in results stems from the difference in genetic background of B. subtilis strains used. Whereas Weart et al. (2007) used strains of PY79 background, we used strains derived from 168 Marburg. Zeigler et al. (2008) have reported that PY79 has four large deletions and replacements of W23-like sequences in its genome compared with 168 Marburg. Changes of both cell length and width in the ugtP mutant compared with WT cells were observed in our study. We also noticed that despite a lack of UgtP, the FtsZ ring was normally localized at the septum (data not shown). Furthermore, the morphology of pgcA and gtaB disruptants was quite similar to that of cells lacking GL (data not shown). Lazarevic et al. (2005) reported balloon-like cell shapes for mutants with defects in genes involved in GL synthesis. However, the reported phenotypes may suggest that sigM was defective in these strains, since we also observed a similar balloon-like morphology in ugtP sigM and ugtP mreB double mutants. On the other hand, the abnormal morphology of ugtP cells was suppressed by MgSO4 (Fig. 4), and the same held for mreB and mbl mutants. We suspect that in these mutants, the cell surface structure is altered, causing an abnormality that a high concentration of MgSO4 is able to remedy. Of course, some other unknown factors may be involved.
DGDG in cell membranes is also used as an anchor of lipoteichoic acids (LTA). In S. aureus, LTA is essential for normal cell growth (Gründling and Schneewind, 2007; Oku et al., 2009). Therefore, there might conceivably be a relationship between the phenotype of the ugtP mutant and LTA. B. subtilis has four LTA synthases (the products of yflE, yfnI, yqgS, and yvgJ), and two enzymes (the products of yflE and yfnI) are believed to be the major LTA synthases (Wörmann et al., 2011). The cells of a strain with a combination of ugtP and yflE yfnI double mutations exhibit a shape similar to that of the ugtP mutant (data not shown). Thus, there is no relationship between change in LTA structure and the morphology of the ugtP disruptant.
The sigma gene sigI of B. subtilis belongs to the group IV heat-shock response genes and seems to be negatively regulated by anti-SigI factor RsgI as are ECF sigmas in B. subtilis (Asai et al., 2007). In the cells lacking GL, some ECF sigmas (SigM, SigV, and SigX; data not shown) and SigI were activated (Fig. 5). The activation of ECF sigmas, but not SigI, in cells lacking GL has been reported (Salzberg and Helmann, 2008). In fact, we did not detect auto-regulation of SigI, but transcription from the SigI-dependent promoter mreBH was activated in the ugtP cells (Fig. 8). Asai et al. (2007) and Tseng and Shaw (2008) have examined the SigI regulon; however, the mechanism of SigI activation is still unclear. Our results indicate that SigI responds to the lack of GL, and is activated by an unknown mechanism. This result may serve as a clue for further studies to reveal the regulation mechanism of SigI and its regulon.
The cells of the ugtP sigM double mutant took on a balloon shape and their growth was sensitive to sodium chloride (Fig. 3). Horsburgh and Moir (1999) showed that a sigM mutant became sensitive to 2% NaCl in NB medium (unlike the wild type), whereas the sigM mutant in our experiments was still resistant to 2% NaCl in LB medium. This difference may be caused by the medium used. SigM is activated for restoring cell surface structures to respond to membrane stress caused by lack of GL, as the SigM regulon is involved in cell surface maintenance (Cao et al., 2002; Salzberg and Helmann, 2008). There would seem to be a close relation between cell surface structure and cytoskeleton to maintain cell shape, because the ugtP mreB double mutant exhibited morphology quite similar to that of the ugtP sigM double mutant. In ugtP mutant cells, mreB expression depending on SigM was increased (Fig. 5B). This may be so in order to compensate for the lability of MreB (Fig. 7). It may be that the cell morphology is prevented from getting worse as a consequence. Thus, once sigM or mreB is disrupted in the ugtP cells, the cell shape completely deteriorates. Kawai et al. (2009a) have reported that the localization of PBP1 depends on MreB. We observed lability of GFP-MreB in the ugtP disruptant at mid-log phase (Fig. 7). Recent papers report that the locations of MreB and cell wall machinery are closely coupled (Garner et al., 2011; Domínguez-Escobar et al., 2011). In the ugtP background, the coupling may be disordered in vegetative growth, and hence (GFP-)MreB may become unstable. We suspected that loss of UgtP might cause this phenomenon and checked interaction between UgtP and MreB; however, no positive interaction was detected by bacterial two-hybrid experiments (data not shown). It therefore seems that GL, the product of UgtP, has important roles in maintaining cell shape. GL may stabilize MreB, or protect MreB from proteolysis. Our results may suggest that GL is a key component connecting cytoskeleton and cell surface structure in B. subtilis. It is a question of some interest, whether this role of GL is specific to B. subtilis or common in GL-containing bacteria.
We are greatly thankful to J. Errington and K. Asai for providing strains involving mreB isoforms. This study was supported by a Grant-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science.
|