2 計算方法
2.1 Wang-Landau法
タンパク質・ペプチド分子のエネルギー空間にはたくさんのエネルギーミニマムが存在し,それらのミニマムは高いエネルギー障壁で遮られている.Nakamuraは,このような複雑なエネルギー空間を効率的にサンプリングするためのエネルギー空間サンプリングアルゴリズムを提案した
[7].その際,Leeのエントロピーサンプリング法 [8]を用いて熱力学量を計算したが,ここでは,Wang-Landau (WL)の方法 [5]を用いて,エネルギーの低いミニマムをできるだけもれなくかつ効率的に探索するためのアルゴリズムを提案する.しかし,このアルゴリズムは,通常のWL法のように長いシミュレーションを行って状態密度を更新していき,最終的に熱力学量を求めるためのものでないことに注意すべきである.
いま,N個の2面角
θ
1
,
θ
2
,
⋯
,
θ
N
を持ったペプチド分子の配座をxとし,そのエネルギーをE(x)とする.ここでは,配座エネルギーE(x)をECEPP/2のポテンシャル
[9]を用いて計算する.つぎに,エネルギー空間E(x)を
|
E
'
(
x
)
=
α
ln
(
E
(
x
)
−
E
0
+
1
)
| (1) |
と変換し,いったん平坦化した空間でサンプリングすることにする.ここで,
α
は理論上のパラメターであり本研究では1.0とした.また,
E0はエネルギー空間
E(
x)におけるグローバルミニマムのエネルギーである.したがって,平坦化したエネルギー空間
E(
x)でのグローバルミニマムのエネルギー
E′は0である.一方,エネルギー
E0は通常不明であるので,サンプリングの開始時には十分高いエネルギーを与え,サンプリング中に
E0より低いエネルギーの配座を訪問するたびに
E0の値を更新していくことにする.このような手続きを繰り返すと,高速にエネルギーの低い配座がサンプリングされるので,エネルギー極小化法としても有効な方法となる.
WL法では,エネルギー空間E′を等確率,すなわち,
P
W
L
(
E
'
)
∝
1
/
Ω
(
E
'
)
でサンプリングするように,エネルギーE′の配座数Ω(E′)を与える.このΩ(E′)はエネルギーE'とE'+ΔE′のあいだにある配座数で,ここではΔE′
=
0.01とした.しかし,Ω(E′)はアプリオリにはわからないので,初期配座数Ω0(E′)として適当な値を仮定する.WL法では,エネルギーE′(x1)の配座からエネルギーE′(x2)の配座への遷移を,
|
T
(
x
1
→
x
2
)
=
[
1
,
Ω
(
E
'
(
x
1
)
)
/
Ω
(
E
'
(
x
2
)
)
]
| (2) |
によって判定する.エネルギー
E′の配座を訪問するたびに,Ω(
E′)に1より大きな
f(
f
> 1)を掛けて
|
Ω
(
E
'
)
n
e
w
=
f
Ω
(
E
'
)
o
l
d
| (3) |
によって配座数を更新していく.この手続きを繰り返すことによってエネルギーの確率分布
PWL(
E′)は一定値に近づいていき,エネルギー空間をランダムウォークするようになる.したがって,このアルゴリズムを使用すれば,ローカルミニマムに落ち込んでも必ず脱出できる.もしも
fに大きな値を与えるならば,局所ミニマムからの脱出が速まり,その結果,少ないステップ数で多くのミニマムを大まかに探索することができる.一方,
fに小さな値を与えるならば,エネルギー空間の狭い領域を細かくサンプリングし,局所ミニマムでのエネルギー極小化を行うことになる.ここでは,エネルギー空間を広く粗くサンプリングする場合には
f
=
eで行い,局所ミニマムでエネルギー極小化を行う場合には
f
と小さくしていくことにする.
初期配座数Ω0(E′)は任意に与えることができるので,エネルギー空間を広く粗くサンプリングする場合には,Ω0(E′)=N
exp
(E′)を使用する.ここで,Nは2面角の総数である.一方,局所ミニマムでエネルギー極小化を行なう場合には,Ω0(E′)=bN1n
(E′+1)を使用する.ここで,bには100∼500の値を与える.このように,式(1)の平坦化したエネルギー空間でWL法を適用した場合,エネルギーE′の初期配座数Ω0(E′)および更新パラメターfの与え方によって配座空間の探索を効率よく制御できる.
2.2 α ヘリックス配座
DYNAとR6W-DYNAのように,比較的残基数の大きくフレキシブルなペプチドでは,配座空間全体をまんべんなく探索することは現段階ではまだ困難である.したがって,これらのペプチドが生体膜内で α ヘリックスを形成することを計算で示すことは実際上難しい.この研究では,DYNAとR6W-DYNAが膜内で α ヘリックスを形成した場合,どのような α ヘリックス配座が最もエネルギー的に安定となるのかを予測したい.すなわち,DYNAの α ヘリックス配座が関わる領域だけの探索に限定する.
そのために,DYNA の初期配座には少なくとも1本の α ヘリックスが含まれるように与える.すなわち,連続した4残基以上の主鎖の2面角(φ,ψ)に(−70°,−40°)
(以後 α Rと表記)の角度を与える.さらに,DYNAの10番目のPro残基が α ヘリックスブレーカーであることを考慮して,その前の残基Arg9の2面角には α R
を与えないという条件を課す.この条件によって,N末端側とC末端側のそれぞれの領域で α R
を連続してとれる残基数は4∼8残基までとなり,これは α ヘリックスの1巻き分から2巻き分に相当し, α ヘリックスの形成可能な本数も全体で1本か最大でも2本である.したがって,DYNAの主鎖の初期配座は,少なくとも1本の α ヘリックスを含み,かつ主鎖の α Rの含有率が88%,75%,63%,50%,38%,25%および13%となるように与えられた. α R以外の主鎖2面角
(φ,ψ)には,ラマチャンドランプロットの原子分子同士の衝突が生じない領域から7個の2面角,(−80°, 70°) ,(−80°, 140°),(−160°, −40°) ,
(−160°, 70°),(−160°, 160°),(70°, 40°),(−90°,
0°)が選ばれた.各 α R含有率に対して1000個ずつの初期配座をランダムに生成し,それぞれの初期配座の周辺領域を探索した.DYNAでは,Figure
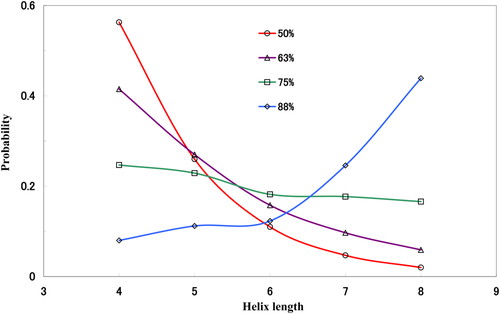
1に示すように,主鎖2面角の α R含有率が50%の場合には残基数が4個(1巻き分)の短い α ヘリックスが50%以上の確率で, α R含有率88%の場合には残基数が8個(2巻分)の長い α ヘリックスがほぼ50%の確率で生成される.図で示されていないが, α R含有率が50%以下の場合には,短い α ヘリックスの形成される確率がさらに高くなる.また,主鎖2面角の α R含有率が大きくなるほど,N末端側とC末端側の両方でそれぞれ α ヘリックスが1本ずつ形成されやすくなる.したがって,いろいろな主鎖の α Rの含有率で α ヘリックス配座を体系的に探索することによって,そのEMCをもれなく求めることが可能となる.各アミノ酸残基の主鎖の2面角ωと側鎖の2面角
χ
i
(
i
=
1
,
2
,
⋯
m
)
の初期値は180°とした.ECEPPでは,プロリン残基のφは−75°に固定されている.
与えられた α R含有率で生成された初期配座から出発して,長いシミュレーションを行うほどより多くのミニマムを求めることができる.しかし,限りあるシミュレーション回数では,探索領域の偏りのためエネルギー安定なミニマムを見逃す可能性がある.特に,残基数が大きくフレキシブルなペプチド分子ほどその可能性が高い.また,採用された初期配座によってはWL法でも局所ミニマムから容易に脱出できない場合もある.それらの危険性を防ぐには,たくさんの初期配座から出発してシミュレーションを行なう必要があるが,同時に1つの初期配座に対するシミュレーション回数を減らさなければならないであろう.どの程度の初期配座数を用意するか,そしてどの程度のシミュレーション回数で行うかという問題に直面する.もう1つの問題は,エネルギー空間にはエネルギー的に安定でないミニマムが非常にたくさん存在しており,それらを探索した分だけ計算効率が悪くなることである.したがって,どのような方法で探索されているミニマムのエネルギーの有利不利を判定するかである.ここでは,以下のような戦略で段階的にミニマムの探索と選別を行うことにする.
第1段階では,与えられた α R含有率において初期配座をたくさん生成し,それぞれの初期配座から出発して短い探索を行なう.このとき,1個の初期配座については,その近くにあるミニマムのエネルギーの高い領域を少し探索して終了することになる.このようにして探索されたミニマムの中から,相対的にエネルギーの低いミニマムだけを選別する.さらに,探索中に基準のエネルギー値を下回った配座だけを集めて,それらの構造情報(2面角の確率分布)を作成しておく.この第1段階の計算によって,エネルギー安定なミニマムの見逃しを十分防ぎ,同時に,エネルギーの高いミニマムの探索を早期に避けることができる.さらに,第2段階では,選別されたミニマムに対して,その周辺領域を少し広い範囲にわたって探索する.このとき,第1段階で得られた2面角の確率分布を用いることにより,低エネルギー領域を効率的に探索する.その探索の結果,再び相対的にエネルギーの低いミニマムだけを選別する.このような探索と選別を数回繰り返した後,十分にエネルギーの低いミニマムだけを残す.第3段階では,第2段階で最終的に絞られた局所ミニマムでのエネルギー極小化を行う.以下に,各段階での計算方法の詳細を記述する.
第1段階では,主鎖の α R含有率に基づいて生成された,1000個の α ヘリックス含有初期構造から出発して1000モンテカルロステップの短いシミュレーションを行い,2面角の確率分布を求める.シミュレーション中,エネルギー
E
'
(
x
)
を持つ配座
x
=
(
θ
1
,
θ
2
,
⋯
θ
k
,
⋯
,
θ
N
)
からエネルギー
E
'
(
x
'
)
の別の配座
x
'
=
(
θ
1
,
θ
2
,
⋯
θ
k
+
Δ
θ
k
,
⋯
,
θ
N
)
へ遷移する場合,(2)式の基準で受け入れるか否かを判定する.ここで,−180° ≤
Δθ ≤
180°である.もしも新しい配座が受け入れられた場合には,その配座の通常のエネルギー空間でのエネルギーE(x′)が平均エネルギー
E
¯
より低ければ,その2面角θk+Δθkを,そうでない場合には2面角θkを1°きざみでセーブする.ここで,平均エネルギー
E
¯
はあらかじめ予備的なシミュレーションで求めておく.1000個の初期配座についてシミュレーションを実行した後,セーブした2面角のヒストグラムから,主鎖の α R含有率における2面角の確率分布,
P
θ
1
,
P
θ
2
,
⋯
P
θ
i
,
⋯
,
P
θ
N
が得られる.ここで,
P
θ
i
は,i番目の2面角θiの確率分布である.ある初期配座の周辺を少し探索したとき,最もエネルギーの低くなった配座を,仮のエネルギー極小配座(仮のEMC)と呼ぶことにする.したがって,1つの α R含有率あたり1000個の仮のEMCが得られ,それらの中で最小のエネルギーを
E
m
(
1
)
とする.仮のEMCのエネルギーと最小エネルギー
E
m
(
1
)
との差が20 kcal/mol以内にあったM1個の仮のEMCだけをセーブする.
第2段階では,主鎖の α R含有率ごとに,第1段階でセーブされたM1個の仮のEMCから探索が開始される.いま,探索中に,エネルギーE(x)を持つ配座
x
=
(
θ
1
,
θ
2
,
⋯
θ
k
,
⋯
,
θ
N
)
から別の配座
x
'
=
(
θ
1
,
θ
2
,
⋯
θ
k
+
Δ
θ
k
,
⋯
,
θ
N
)
へ遷移する場合,Nakamuraの方法に従って,まず新たな2面角θk+Δθkが許容されるか否かが
T
'
(
θ
k
→
θ
k
'
)
=
(
1
,
P
θ
k
/
P
θ
k
'
)
によって判定される [10].許容された配座に対してのみエネルギーE(x′)が計算され,遷移確率T(x
→ x′)) =
[1,Ω(E(x))/Ω(E(x′))]によって受け入れるか否かが判定される.このような探索を104
モンテカルロステップだけ行ない,この局所ミニマムでの仮のEMCを求める.同様な探索を第1段階でセーブされた仮のEMC周辺すべてに対して行うならば,M1個の仮のEMCが得られる.この探索での最小エネルギーが
E
m
(
2
)
ならば,それと仮のEMCのエネルギーで差が20 kcal/mol以内にあるM2個の局所ミニマムが選ばれる.以下同様の手続きで,第2段階のシミュレーションを2∼5回繰り返して,エネルギーの低い仮のEMCを選別していく.最小エネルギー
E
m
(
n
)
の収束状況によって,第2段階での計算の打ち切りの判断を行う.
第3段階では,第2段階で絞られた局所ミニマム内にある仮のEMCから出発して,(3)式の更新パラメターfの値をe,初期配座数をΩ0(E′)=bN1n
(E′+1)としてエネルギー極小化を行う.このエネルギー極小化では,初期配座の近傍で細かく探索されるように,配座
x
=
(
θ
1
,
θ
2
,
⋯
θ
k
,
⋯
,
θ
N
)
から別の配座
x
'
=
(
θ
1
,
θ
2
,
⋯
θ
k
+
Δ
θ
k
,
⋯
,
θ
N
)
への遷移が−30° ≤ Δθk ≤
30°の比較的狭い角度範囲内で行われる.ただし,このエネルギー極小化では2面角の確率分布を用いた許諾の判定は行わない.エネルギー極小化は5 ×
104モンテカルロステップだけ実行される.もしもこのミニマム内でのエネルギー極小化の結果,(1)式の最小エネルギーE0が104モンテカルロステップの間0.01 kcal/mol
以下で変化しない場合,EMCに到達したと判定する.もし到達していないならば,Ω(E′)をΩ0(E′)にリセットし,更新パラメターを
f
と小さくして2回目のエネルギー極小化を続ける.この手続きを繰り返すことによって,最終的なEMCが得られる.
2.4 溶媒中での自由エネルギー
この研究では,真空中で得られた最終的なEMCをシクロヘキサン溶媒へ移行したときの溶媒からの間接的なエネルギー寄与gcをEisenbergとMcLachlanに従って,
|
g
c
=
∑
i
M
σ
i
A
S
A
i
| (4) |
によって計算する [
11].ここで,
ASA1はi原子のASA,
σ1はi原子のASP,
M
は移行自由エネルギー に寄与する原子数である.原子のASAは,FraczkiewiczとBraunの方法 [
12]を用いて計算し,原子のASPはEfremovらの値 [
6]を使用する.ここでは,溶媒での自由エネルギー
Gcを単に,
で見積もることができると仮定する.ここで
Eは真空中で得られたEMCの配座エネルギーである.