私たちは最も視力の優れた中心窩で興味のある物体を見るために頻繁に眼球を動かしている。このような眼球運動により,網膜像も頻繁に動いているにも関わらず,私たちは安定した視覚世界を知覚する。このような安定した視覚世界を知覚するために,眼球運動時にどのように視覚情報は処理されているのか?本稿では,眼球運動時の視覚情報処理に関する二つの研究について紹介する。一つ目は,サッカードと呼ばれる眼球運動が生じているときのサッカード前後の視野統合に関する研究(Matsumiya, Sato & Shioiri, 2016)を,そして二つ目は,追従眼球運動と呼ばれる眼球運動を駆動する際に用いられる視覚的な動きの信号に関する研究(Matsumiya & Shioiri, 2015)を紹介する。
私たちは最も視力の優れた中心窩で興味のある物体を捉えるために頻繁に眼球を動かしている。例えば,スーパーマーケットで所望の品が商品棚にあるかどうかを探すときには,サッカードと呼ばれる跳躍的な眼球運動を行って,視線の位置をある品物から別の品物へと変えていく。また,人通りの多い人混みの中で友人を探すときには,歩行者の顔を追跡する。このときには,追従眼球運動と呼ばれる眼球運動を行う。この眼球運動により,動いている視覚対象が中心窩に固定され,その視覚対象が何であるかをはっきりと捉えることができる。ここでは,サッカード時8)および追従眼球運動時18)の視覚情報処理に関する著者らの研究を紹介する。
サッカード眼球運動が生じると,視覚世界から投影される網膜像が動く。それにも関わらず,我々は,視覚世界は安定していると知覚する。視覚系はこの視野安定をどのように実現しているのか?古典的な説明の一つに,視覚世界はサッカード中に変化しないことを視覚系が仮定しているという説がある1)。サッカード時には小さな標的の変位を検出できないことが知られており,サッカード変位抑制と呼ばれている(図1a)2)。この現象により,視覚世界が安定して見えることから,上記の説を支持していると考えられる。

サッカード眼球運動時の変位知覚。(a)サッカード中に標的が変位すると,変位を知覚できない。(b)ブランク効果。サッカード中に標的が消え,サッカードが終わった100 ms後に標的を異なる位置に再呈示すると,標的の位置変化を容易に検出できる。(c)ランドマーク効果。サッカード中に標的が消え,サッカードが終わった100 ms後に標的をサッカード前と同じ位置に再呈示する。一方,標的のすぐ上に呈示されたランドマークは,画面から消えることなく,サッカード中に位置が変わる。このとき,被験者は,ランドマークではなく,標的の位置が変化したと知覚する。
しかし,近年の研究により,視野安定機構はサッカード変位抑制だけではないことが指摘されている。サッカード時に標的を一時的に消失させると変位検出能力が改善する(ブランク効果;図1b)3)。また,サッカード時に標的を一時的に消失させ,周囲の刺激の位置をずらすと,周囲の刺激でなく標的の変位が知覚される(ランドマーク効果;図1c)4)。これら二つの効果は,サッカード後の標的の一時的消失による輝度情報の過渡的な変化によって引き起こされる。サッカード時は,通常,過渡信号に感度を持つシステムは抑制されていると考えられているが5–7),サッカード後の一時的消失がこの過渡信号に感度を持つシステムを活性化させている可能性がある。もし,これが本当であるならば,過渡信号に感度を持つシステムの活性化がサッカード時の標的の変位判断を改善させていることが予想される。
この可能性を検証するために,標的の輝度コントラストがブランク効果とランドマーク効果に及ぼす影響を調べた8)。もし過渡システムがサッカード時の変位検出に関わっているのであれば,ブランク効果とランドマーク効果の強度が輝度コントラストに依存して変化することが予想される。図2に示されるように,サッカード標的がサッカード中に一時的に消失する場合(ブランクあり条件)は,サッカード標的の輝度コントラストが高いときに標的の変位検出率が高かった。サッカード標的がサッカード中に一時的に消失しない場合(ブランクなし条件)は,すべての輝度コントラストを通して標的の変位検出率は低かった。これより,ブランク効果の強度が輝度コントラストに依存して増すことがわかる。一方,ランドマーク効果への輝度コントラストの影響が図3に示される。ランドマーク効果を調べるときに用いられる被験者への課題では,画面に呈示された標的とランドマークのどちらが変位したかを応答させる。そのため,図3の縦軸は,標的の変位検出率ではなく,ランドマークが変位したすべての試行に対して標的が変位したと応答した割合(■)と標的が変位したすべての試行に対して標的が変位したと応答した割合(○)を示している。ランドマーク効果は,ランドマークが変位したにも関わらず標的が変位したと知覚する錯視であるため,ランドマークが変位したすべての試行に対して標的が変位したと応答した割合(■)が高いという結果は,ランドマーク効果が頻繁に生じていたことを示している。図3より,輝度コントラストが低いときに,ランドマーク効果が頻繁に生じており,輝度コントラストの増大に伴い,その頻度は低下していることがわかる。これらの結果は,ランドマーク効果の強度が輝度コントラストに依存して減少することを示している。
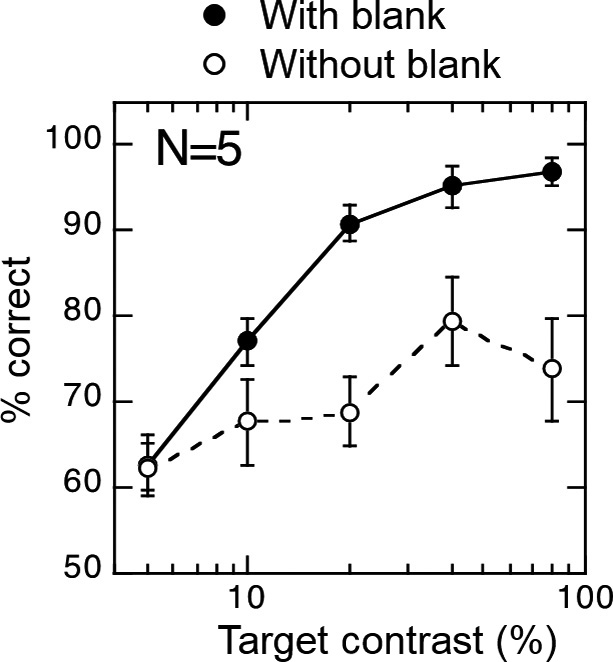
ブランク効果における輝度コントラストの影響。黒丸がブランクあり条件,白丸がブランクなし条件。N = 5。(Matsumiya et al., 2016)

ランドマーク効果における輝度コントラストの影響。黒四角がランドマーク変位条件,白丸が標的変位条件。N = 5。(Matsumiya et al., 2016)
ブランク効果とランドマーク効果は,ともに標的の一時的消失を含んでいるため,これらの二つの効果は共通した輝度情報の過渡システムの活性化に依存していることが予想される。このような共通した過渡システムが存在するかどうかを調べるために,ブランク効果とランドマーク効果の各々に対して信号検出理論9)を使って標的の検出感度と反応バイアスが輝度コントラストに応じてどのように変化するかを調べた。信号検出理論では,標的の検出感度を信号分布とノイズ分布のピーク間の差として定義し,反応バイアスを信号分布のピークとノイズ分布のピークの中間地点を基準にして,反応バイアスをその中間地点からのずれとして定義する。本実験では,標的の検出感度d′を算出するために,ブランク効果の実験において,標的の右(左)変位に対する正しい応答をhit,左(右)変位に対する間違った応答をfalse alarmと定義した。これらのhitとfalse alarmの割合から,ブランク効果におけるブランクあり条件とブランクなし条件に対するd′を計算した。ランドマーク効果の実験においては,標的変位に対する正しい応答をhit,ランドマーク変位に対する間違った応答をfalse alarmと定義した。これらのhitとfalse alarmの割合から,ランドマーク効果におけるブランクあり条件に対するd′を計算した。ランドマーク効果におけるブランクなし条件のd′を算出するために,Deubel et al.(1998)のデータを用いた4)。そして,ブランク効果とランドマーク効果の各々に対して,標的の一時的消失の効果を定量化するためにブランクあり条件となし条件のd′の差を算出し,この差を標的消失効果と定義した。
図4より,輝度コントラストとともにブランクあり条件の標的の検出感度が増大することを示し,ブランク効果とランドマーク効果で類似した傾向を示した。この結果は,サッカード時に標的変位を検出する能力が輝度コントラストの増加に伴い促進され,このコントラストの効果はブランク効果とランドマーク効果の両方において類似していることを示しており,ブランク効果とランドマーク効果において,一時的消失による標的変位判断の変化は共通した処理機構であることを示唆する。従って,図3の結果の説明として,ランドマーク効果においては,標的の一時消失効果がより強くなることにより,視覚系は標的の位置を正確に判断できるようになるため,標的変位の錯覚であるランドマーク効果が減少したと考えられる。

信号検出理論により算出された標的の検出感度d′。(a)ブランク効果の実験データから算出された結果。(b)ランドマーク効果の実験データから算出された結果。(c)ブランク効果とランドマーク効果の比較。ブランクあり条件となし条件のd′の差を算出し,この差をブランク消失効果と定義した。(Matsumiya et al., 2016)
図5より,ブランク効果の実験から得られたブランクあり条件のデータから算出された反応バイアスは,ほぼゼロであった。一方,ランドマーク効果の実験から得られたブランクあり条件のデータから算出された反応バイアスは,ゼロではなく負の値となり,標的の輝度コントラストには依存しなかった。この結果より,ランドマーク効果が,ブランク効果と違い,物理的には変位していない標的を変位したと知覚する錯覚であることを考えると,この錯覚は反応バイアスとして説明できることが示唆される。

信号検出理論により算出された反応バイアスλ。丸印がブランク効果の結果。四角印がランドマーク効果の結果。(Matsumiya et al., 2016)
以上の結果は,サッカード後に呈示される輝度に基づいた過渡信号が標的の位置判断を正確にすることを示唆し,ブランク効果で見られた位置判断の向上はこの過渡信号の関与によると考えられる。一方,ランドマーク効果では,過渡信号により,正確に位置判断ができるようになったため,ランドマーク効果が減少したと考えられる。もしこの考えが正しいのであれば,輝度による過渡信号を含まない等輝度色刺激を用いると,ブランク効果は消失するが,ランドマーク効果は生じるかもしれない。予想通り,等輝度色刺激では,ブランク効果は消失し,ランドマーク効果は生じた(結果の詳細については文献8)を参照)。輝度情報を含まない等輝度色情報を用いるとブランク効果が消失することから,サッカード前後の視野統合による位置判断は,輝度に基づいた過渡信号により選択的に促進されていることが示唆される。
サッカードによる知覚抑制の生理学的機序は,外側膝状体の大細胞層経路の選択的抑制によると主張する研究がある6,7)。この大細胞層経路は,動きや位置の空間知覚に関わると考えられており,大細胞層経路の抑制がサッカードによって生じる網膜像の動き信号の知覚を防いでいるのかもしれない5)。一方,サッカード後に呈示される輝度に基づいた過渡信号により,サッカード時の変位検出が回復するため,この輝度に基づいた過渡信号はサッカードに伴う大細胞層経路の選択的抑制を回避していると考えられる。
また,外側膝状体には,大細胞層経路だけでなく,小細胞層経路も存在する10)。小細胞層経路は色情報の寄与が示唆されている11)。ランドマーク効果は等輝度色刺激でも生じることから小細胞層経路の処理が関わっているのかもしれない。等輝度色刺激を使ったときにサッカード時に生じる知覚現象には,ランドマーク効果以外に,視空間圧縮と呼ばれる現象も報告されている12)。この視空間圧縮はランドマーク効果と類似した特性を持ち13),輝度コントラストが低いときに視空間圧縮が強く生じる14)。ランドマーク効果も視空間圧縮も,背景に対する標的の位置判断という点で共通しており,このような相対的な位置判断には小細胞層経路が関与しているのかもしれない。そして,このような生理学的機序が反応バイアスを生み出しているのかもしれない。
我々は運動物体の動きの方向を知覚すると同時に,その動きを眼球運動により追跡することができる。このとき動きの方向知覚と眼球運動による動きの追跡は同じ視覚入力を受け取る。動きの方向知覚では知覚される方向を決定するために,眼球運動による動きの追跡では眼球運動を駆動するために,その視覚入力を用いることになる。このように同じ視覚入力でも,最終的な出力が知覚か眼球運動かに応じて,その視覚入力は別々の経路で処理されるのであろうか。
Goodale and Milner(1992)は,行動制御に用いられる視覚情報は知覚に用いられる視覚情報と異なった経路で処理されるという考えを提案している15)。この考えによると,動きの方向を知覚し眼球でその動きを追跡している場合,ここでの視覚入力は方向知覚と眼球運動で別々に処理されている可能性がある。この可能性を支持する研究はいくつか存在する。例えば,物理的な網膜像の動きと知覚される動きの間に矛盾があるような特殊な実験条件で追従眼球運動を測定すると,知覚される動きではなく,物理的な網膜像の動きを追跡するように眼球運動が発生する16)。しかし,多数の研究が動きの知覚と追従眼球運動の間で動き信号を共有していることを示唆しており17),追従眼球運動における知覚と行動の関係について,過去の研究の間でなぜ見解が異なるのか,その理由は明確ではない。
見解が異なる原因の一つとして,運動視の処理機構には二つの処理過程が存在することを考慮に入れていないことが挙げられる。知覚と行動で動き信号の乖離を報告した過去の研究では,運動刺激が運動視処理の一方の処理過程に関与し,知覚と行動で動き信号の共有を報告した研究では,運動刺激が運動視処理のもう一方の処理過程に関与していたのかもしれない。私たちはこの可能性を検討するために,運動視処理機構に存在する,遅い運動と速い運動の二つの処理過程に着目し,各々の処理過程と追従眼球運動がどのような関係にあるのかを調査した18)。
本研究では,低空間周波数と高空間周波数の刺激を空間的に重ね,各々を異なった方向に運動させた刺激に順応した後に生じる運動残効を用いた(図6)19)。5 Hzでドリフトする低空間周波数の刺激は速い運動成分を,5 Hzでドリフトする高空間周波数の刺激は遅い運動成分を持つ。この刺激の順応後に生じる運動残効では,時間周波数が0 Hzである静止テストを用いると,遅い運動である高空間周波数の順応刺激に対する運動残効が優位に現れ,時間周波数が4 Hzのフリッカーテストを用いると,速い運動である低空間周波数の順応刺激に対する運動残効が優位に現れる。これは,低時間周波数のテスト刺激による運動残効は遅い運動に感度を持つ運動検出器の応答を反映し,高時間周波数のテスト刺激による運動残効は速い運動に感度を持つ運動検出器の応答を反映していることを示している。すなわち,テスト刺激の時間周波数を変えることで,二つの運動視処理過程を独立して扱うことが可能となる。

刺激と実験手続き。低空間周波数と高空間周波数の刺激を空間的に重ね,各々を異なった方向に運動させた刺激に順応する。(Matsumiya & Shioiri, 2015)
運動残効では,テスト刺激は時間周波数によらず物理的に静止しているにも関わらず,知覚的に動きが見える現象である。そのため,運動残効で知覚される動きによって追従眼球運動を駆動できた場合は,眼球運動を駆動するために用いられる動きの信号は知覚と共有されていることになる。さらに,追従眼球運動と動きの知覚が動きの信号を共有しているのかどうかを正確に捉えるために,刺激の輝度コントラストを系統的に変化させ,知覚される運動残効の方向と運動残効時の追従眼球運動の方向を調べ比較した。眼球運動と知覚が動きの信号を共有しているならば,輝度コントラストが動きの方向に与える影響は眼球運動と知覚で一致するはずである。
図7に,ある一試行においてテスト刺激が画面上に呈示されてから消えるまでの2秒間に計測された眼球運動データの一例を示す。眼球運動の移動方向は,眼球運動データ中の眼球位置の始点と終点を使って算出された。画面中央から下方向をゼロ度として,高空間周波数成分の縞の運動方向と反対方向を正の角度として定義した。図8に知覚された運動残効の方向と眼球運動の移動方向の結果を示す。縦軸は運動残効方向と眼球運動方向を示しており,正の値は高空間周波数成分の残効方向に重みがあることを示す。横軸は順応刺激のコントラストを示す。コントラストは各周波数単独で運動している条件による観察時の閾値を基準として,その倍数で設定した。静止テストを呈示するときの順応刺激は,低周波成分のコントラストをコントラスト閾値の30倍に固定し,高周波成分のコントラストを実験変数として閾値の7.5倍から60倍まで変化させた。フリッカーテストを呈示するときの順応刺激は,高周波成分のコントラストをコントラスト閾値の30倍に固定し,低周波成分のコントラストを実験変数として閾値の7.5倍から60倍まで変化させた。図8aに静止テストを呈示したときの結果を示す。高空間周波数のコントラストが閾値の30倍以上のときに,順応時の高空間周波数成分の動きと反対方向に近い方向を運動残効方向として知覚していたことがわかる。また,運動残効の方向が,相対的なコントラスト比に依存して低空間周波数成分に対するものから高空間周波数成分に対するものに連続的に変化した。一方,眼球運動の方向も,知覚された運動残効の方向とほぼ同じ傾向を示した。図8bにフリッカーテストを呈示したときの結果を示す。順応時の低空間周波数成分の動きと反対方向に近い方向を運動残効方向として知覚し,相対的なコントラスト比に依存して知覚方向が連続的に変化した。眼球運動の方向も,知覚された運動残効の方向とほぼ同じ傾向を示した。これらの結果は,遅い運動と速い運動の両者に対して,追従眼球運動と知覚は共通の運動処理機構で処理されていることを示唆する。

静止テスト観察時に生じた追従眼球運動の一例。(Matsumiya & Shioiri, 2015)

知覚された運動残効の方向と眼球運動の移動方向の結果。(a)静止テスト。(b)フリッカーテスト。N = 5。(Matsumiya & Shioiri, 2015)
本稿では,サッカード眼球運動時の視野安定機構に関する研究と,追従眼球運動と動きの知覚の関係に関する研究を紹介した。サッカード時の視野安定機構は未だに明らかにされていない視覚科学における重要な問題の一つである。我々の研究により,視野安定機構には,輝度の過渡信号を利用して正確に位置判断する機構と,背景に対する相対的な位置を判断する機構の二つが存在する可能性が示唆された。前者の機構は,過渡信号が呈示されることで,網膜位置信号に加えて眼球位置信号が用いられることで正確な位置判断を可能にしていると思われる。一方,後者の機構は,眼球位置信号を用いず,網膜位置信号だけを用いてサッカード前後の視野統合を行っているのかもしれない。これらのメカニズムの詳細については,今後の研究課題である。
また,追従眼球運動における知覚と行動の関係については,乖離した結果を見出すことはできなかった。過去の研究には,追従眼球運動と動きの知覚が乖離した証拠を示しているものもあるが,大多数の研究がこれらの間で動きの信号を共有していることを示している。少なくとも,遅い運動と速い運動の処理過程においては,追従眼球運動と動きの知覚は共通の運動処理機構で処理されていると考えられる。追従眼球運動における知覚と行動の関係に関する過去の研究に見られる結果の違いは,どのように説明できるのか? これからの研究課題として重要であると考える。