Abstract
With the aim of searching for potent diazotrophic bacteria that are free of public health concerns and optimize rice cultivation, the endophytic colonization and plant growth-promoting activities of some endophytic diazotrophic bacteria isolated from rice were evaluated. Among these bacteria, the emerging diazotrophic strains of the genus Novosphingobium effectively associated with rice plant interiors and consequently promoted the growth of rice, even with the lack of a nitrogen source. These results suggest that diazotrophic Novosphingobium is an alternative microbial resource for further development as a safe biological enhancer in the optimization of organic rice cultivation.
Rice (Oryza spp.) is an important crop for human nourishment and is grown worldwide. The cultivation of commercial rice often requires nitrogen fertilizers in order to maintain and increase rice yields. However, traditional agricultural practices for the cultivation of local rice for household consumption (e.g., without chemical input) also sustain reasonable rice yields each year (10, 13). The sustainability of rice yields involves various environmental factors, particularly soil-atmosphere nutrient cycles by beneficial microbes that occur in the rhizospheres of rice (6, 7, 13–15). The benefits of plant-microbe interactions are well-known factors that promote the growth of diverse rice cultivars to reach sustainable rice yields (6, 7).
Nitrogen-fixing microbes (so-called “diazotrophs”) offer significant benefits in the optimization of rice cultivation because they fix atmospheric nitrogen and supply usable forms of nitrogen to rice plants (14, 15). With the exception of obligate symbiotic diazotrophs (e.g., Rhizobium), many other diazotrophs fix an unlimited amount of nitrogen in the free-living and endophytic phases (e.g., Klebsiella and Sphingomonas) (14, 17). Attentive selection is essential for the further implementation of unlimited nitrogen-fixing microbes in rice cultivation because some of these microbes have been identified as health risks for farmers (e.g., Klebsiella pneumoniae).
We recently revealed the nitrogen-fixing potential of endophytic diazotrophic bacteria isolated from the plant tissues of many rice landraces grown in northern Thailand (14). These endophytic diazotrophic bacteria belonged to diverse bacterial taxa, while some exhibited relatively high nitrogenase activities in the free-living and endophytic phases. In the present study, we aim to screen for potent diazotrophs among our previous list of endophytic diazotrophic bacteria, which are free of public health concerns and enhance rice growth with the lack of a nitrogen input. We selected some distinctive strains of endophytic diazotrophic bacteria from our previous study (14) listed in Table 1 for further evaluations of their taxonomy, endophytic colonization, and plant growth-promoting activities.
Table 1
Distinctive strains of endophytic diazotrophic bacteria used in this study.
| Bacterial straina |
GenBank accession no. |
Closest species |
Identity (%)b |
Different nt/Total ntb |
| PR1 |
JX083379 |
Burkholderia kururiensis JCM 10599T |
100.00 |
0/1489 |
| PS5 |
JX083381 |
Novosphingobium sediminicola HU1-AH51T 9 |
7.03 |
40/1346 |
| SR6 |
JX083384 |
Novosphingobium sediminicola HU1-AH51T |
97.10 |
35/1207 |
| SS2 |
JX083385 |
Klebsiella pneumoniae subsp. pneumoniae DSM 30104T |
99.56 |
6/1351 |
| SS5 |
JX083386 |
Burkholderia kururiensis JCM 10599T |
100.00 |
0/1325 |
| SS6 |
JX083387 |
Novosphingobium sediminicola HU1-AH51T |
96.98 |
41/1356 |
a The origins of these bacterial strains are shown in (
13).
b Results are taken from the alignment search tool of EzTaxon (www.ezbiocloud.net, search on 26 May 2016).
The classification of the selected strains was amended by a comparison of their 16S rRNA gene sequence data to those in the publicly available database in EzTaxon (www.ezbiocloud.net). All related sequences were selected, consequently aligned with MUSCLE, and used to construct the phylogenetic tree by MEGA7 (www.megasoftware.net). Among the selected strains, a representative from each taxonomic cluster was selected for labeling with red and green fluorescence tracks using a cloning technique. The pBZ1::dsRed2 (11) and pRK404 (9) plasmids were constructed and served as red and green fluorescence tracks, respectively. Each plasmid was introduced into Escherichia coli S17-1 (pro recA RP4-2(Tcs::Mu) (Kms::Tn7); Mob+) by an electroporation protocol described by Noisangium et al. (9). The plasmids were extracted from the obtained E. coli S17-1 transformant and purified with the QIAprep Spin Miniprep kit (QIAGEN, USA). The purified plasmids were introduced into the competent cells of our selected strains, using the same electroporation protocol described above (9). A modified method was performed in order to prepare competent cells from our selected strains (Supplemental Materials, Method S1). Fluorescence-labeled strains were used in further evaluations.
The commercial Thai Jasmine rice cultivar (Oryza sativa var. KDML 105) served as a mutual plant in the present study, and its rice seeds were surface sterilized and allowed to germinate on modified semisolid (MS) medium in a 50-mL test tube, following the protocol described elsewhere (3). Experiments were conducted either in the presence or absence of a water-soluble nitrogen input (NH4NO3). Therefore, MS medium supplemented with NH4NO3 at a final concentration of 0.2 mM served as the N-available treatment, and non-supplemented MS medium acted as the N-free treatment. After germinating for 3 d, the seedlings were inoculated with 1 mL of each fluorescence-labeled strain (at a cell density of ~107 cells mL−1) and then allowed to grow at 27°C under the light-dark alternation (16 h in the light and 8 h in the dark). Colonized rice plant tissues were sampled and used for surface sterilization with a protocol described previously (14), and this was followed by cross-sectioning at a thickness of 100 μm using Microm HM 650V (Thermo Scientific, USA).
The endophytic colonization of fluorescence-labeled strains was tracked and observed with an inverted fluorescence microscope (Olympus IX51, USA). Viable counts of bacterial cells colonized at different rice plant interiors were also confirmed (Method S2). The heights of rice shoots represented a growth indicator of rice in this study; they were measured and calculated in percentages of increased heights over those measured in the absence of the bacterial inoculum (control). Any means±SDs or SEs derived from at least triplicate experiments were statistically compared using an analysis of variance (ANOVA) in SPSS 22.0 software (SPSS, Chicago IL, USA). Statistical results are indicated with F-distribution values, degrees of freedom, and significance (P) levels.
The phylogenetic reclassification with the Maximum Likelihood method revealed that potent endophytic diazotrophic bacteria belonged to the families Burkholderiaceae, Enterobacteriaceae, and Sphingomonadaceae (Fig. S1). The closely related phylogenetic species of these bacteria listed in Table 1 were different to our previous classification (14). Among all isolates, three were members of the genus Novosphingobium (formerly classified in the genus Sphingomonas [14]) and closely related to N. sediminicola HU1-AH51T. However, the % identities of these Novosphingobium isolates to their closest species were relatively small (96.98–97.10%), suggesting the high possibility of their species novelty. This range of % identities was similar to a recently released species, N. oryzae that was proposed as a novel species with ≤97.2% identities to other species in the same genus (18). Since the establishment of the genus Novosphingobium in 2001 (16), a few of its members have been identified as endophytes of rice (e.g., N. fluoreni from rice seeds [4] and N. oryzae from rice roots [18]). Previous studies revealed the plant growth-promoting potential of the genus Novosphingobium (1, 14, 18), while N. nitrogenifigens is so far the first and only diazotrophic strain of this genus (1).
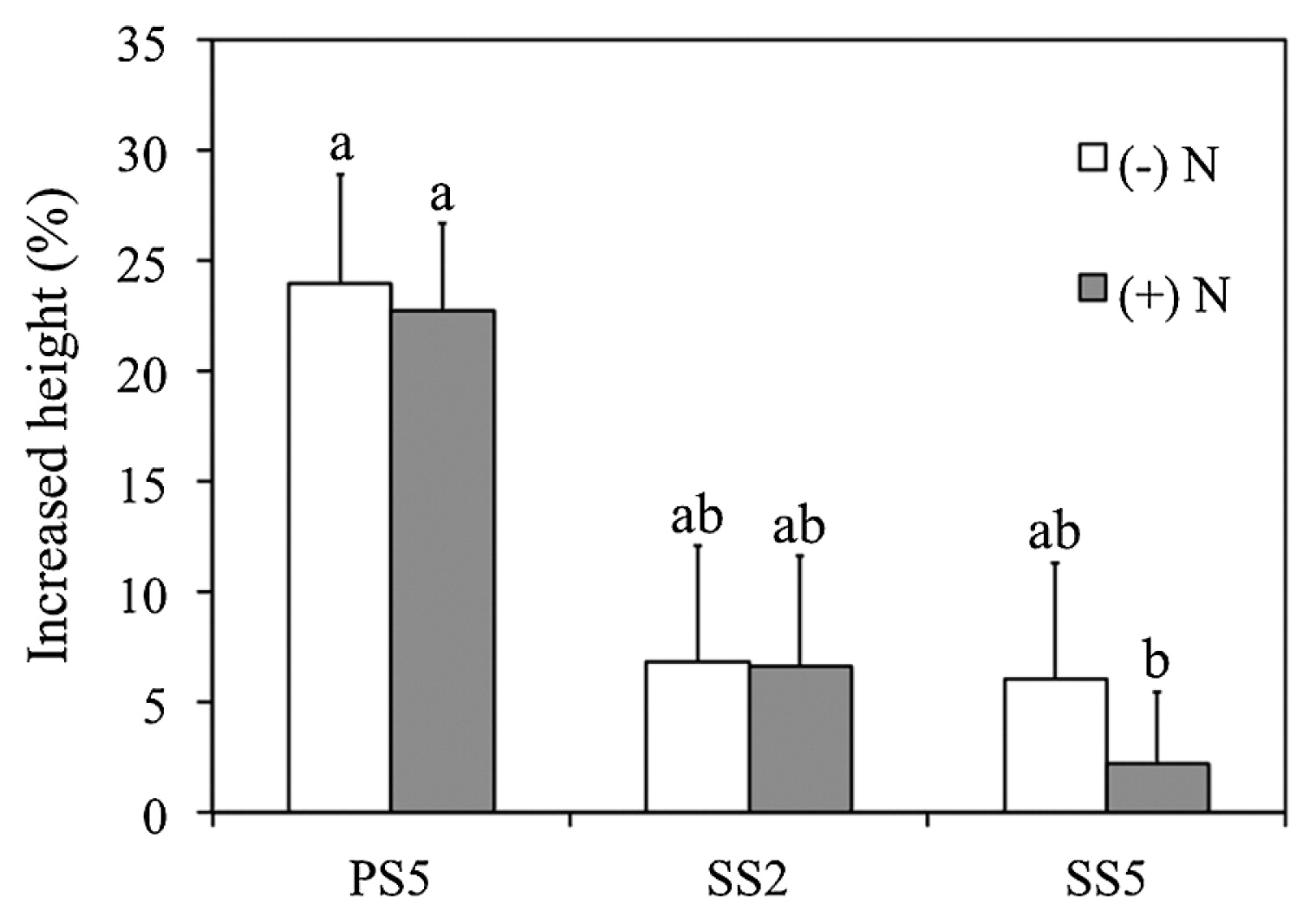
We selected Novosphingobium sp. PS5, Klebsiella sp. SS2, and Burkholderia sp. SS5 as a representative of each genus for further comparative studies of their endophytic colonization and plant growth-promoting potentials for optimizing rice cultivation. The endophytic colonization of rice interiors by Novosphingobium sp. PS5 is shown in Fig. 1. At 1 d after inoculation (DAI), strain PS5 colonized root hairs by forming microcolonies on the surfaces of epidermal cells, particularly around the emerging sites of lateral roots. Consequently, strain PS5 invaded the plant interiors at the parenchymatous root cortex, from which it migrated through the intercellular and intracellular spaces of the rice shoots, vascular bundles of leaves, and lateral roots (at 7 DAI). Similar colonizing patterns were observed with Klebsiella sp. SS2 (Fig. S2) and Burkholderia sp. SS5 (Fig. S3). We confirmed the abundant occupancies of the test bacteria colonized within the root and shoot interiors of rice with their viable counts (Table S1). The results obtained suggested that rice roots were an optimal habitat, housing large numbers of every test bacteria, while Novosphingobium sp. PS5 exhibited the highest abundance in the colonization of root and shoot interiors. The greater % of the increased height of rice shoots also corresponded to the abundant occupancy within the rice plant interiors of strain PS5 (Fig. 2).
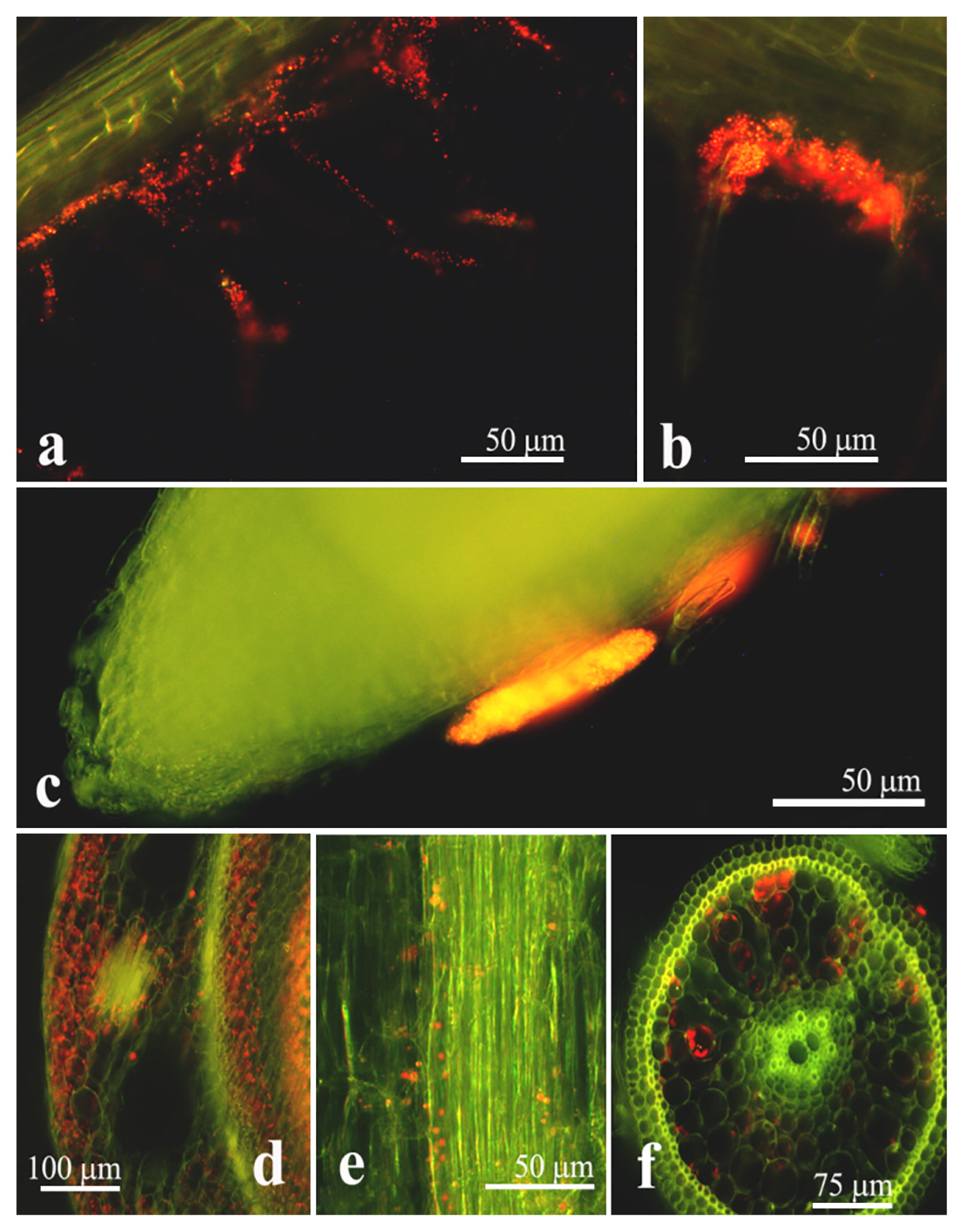
The mechanisms by which diazotrophic Novosphingobium invaded into and migrated within the rice interiors have yet to be elucidated. The plant rhizosphere and root exudates are considerable boundaries that offer an optimal ecological niche for synergistic interactions between plants and soil-dwelling microbes. Wound sites and openings that occur naturally by elongation and differentiation during rice growth and development are recognized as potential routes for plant invasion by endophytic diazotrophic bacteria (5, 7). This may support the abundance of bacterial cells that initially colonized these wound sites and openings in rice roots. The possession of cellulolytic enzymes has been proposed as a biochemical tool that endophytic diazotrophic bacteria use to invade the plant interior (12). The bacterial strains tested in the present study also exhibited the ability to form at least two cell wall lytic enzymes, i.e., cellulase, pectinase, and chitinase (14). Further studies on how these enzymes are expressed and function during the invasion and colonization of plant interiors will contribute to a better understanding of the biological processes involved in plant-microbe interactions. Based on our results, rice seedlings were healthy and there were no apparent disease symptoms detected after endophytic colonization by any bacteria tested, which supports mutual interactions between them and rice plants.
It is conceivable that nitrogen input in the cultivation of rice may either interfere with the community of diazotrophic bacteria or suppress their nitrogen-fixing activities (8, 12). Furthermore, endophytic colonization is a mechanism by which diazotrophic bacteria protect themselves and maintain their nitrogen-fixing activities from surrounding stresses (e.g., oxygen-induced damage of nitrogenase, competition with other rhizobacteria) (2). Our previous findings showed that all bacteria tested were potent diazotrophs with the ability to fix atmospheric nitrogen either in the absence or presence of a water-soluble nitrogen input, and either in their free-living phase or in association with their mutual rice plant (14). Diazotrophic Novosphingobium exhibited the most distinctive endophytic colonization and plant growth-promoting potentials, as well as the free of public health concern, when compared to the other bacteria tested. We conclude that the diazotrophic Novosphingobium would be a promising microbial resource for further development as an efficient and safe biofertilizer for optimizing the organic rice cultivation.
Acknowledgments
The authors would like to acknowledge funding support by the Office of the Higher Education Commission, Thailand, under the National Research University Project and the Thailand Research Fund (Grant: RTA5880006).
References
- 1. Addison, S.L., S.M. Foote, N.M. Reid, and G. Lloyd-Jones. 2007. Novosphingobium nitrogenifigens sp. nov., a polyhydroxyalkanoate-accumulating diazotroph isolated from a New Zealand pulp and paper wastewater. Int J Syst Evol Microbiol. 57:2467-2471.
- 2. Danhorn, T., and C. Fuqua. 2007. Biofilm formation by plant-associated bacteria. Annu Rev Microbiol. 61:401-422.
- 3. Elbeltagy, A., K. Nishioka, T. Sato, H. Suzuki, B. Ye, T. Hamada, T. Isawa, H. Mitsui, and K. Minamisawa. 2001. Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Appl Environ Microbiol. 67:5285-5293.
- 4. Gao, S., Y. Zhang, N. Jiang, L. Luo, Q.X. Li, and J. Li. 2015. Novosphingobium fluoreni sp. nov., isolated from rice seeds. Int J Syst Evol Microbiol. 65:1409-1414.
- 5. Hardoim, P.R., L.S. van Overbeek, and J.D. van Elsas. 2008. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 16:463-471.
- 6. Isawa, T., M. Yasuda, H. Awazaki, K. Minamisawa, S. Shinozaki, and H. Nakashita. 2010. Azospirillum sp. strain B510 enhances rice growth and yield. Microbes Environ. 25:58-61.
- 7. Mano, H., and H. Morisaki. 2008. Endophytic bacteria in the rice plant. Microbes Environ. 23:109-117.
- 8. Muthukumarasamy, R., U.G. Kang, K.D. Park, et al. 2007. Enumeration, isolation and identification of diazotrophs from Korean wetland rice varieties grown with long-term application of N and compost and their short-term inoculation effect on rice plants. J Appl Microbiol. 102:981-991.
- 9. Noisangiam, R., K. Teamtisong, P. Tittabutr, N. Boonkerd, U. Toshiki, K. Minamisawa, and N. Teaumroong. 2012. Genetic diversity, symbiotic evolution, and proposed infection process of Bradyrhizobium strains isolated from root nodules of Aeschynomene americana L. in Thailand. Appl Environ Microbiol. 78:6236-6250.
- 10. Oupkaew, P., T. Pusadee, A. Sirabanchongkran, K. Rerkasem, S. Jamjod, and B. Rerkasem. 2011. Complexity and adaptability of a traditional agricultural system: case study of a gall midge resistant rice landrace from northern Thailand. Genet Resour Crop Evol. 58:361-372.
- 11. Piromyou, P., T. Greetatorn, K. Teamtisong, et al. 2015. Preferential association of endophytic bradyrhizobia with different rice cultivars and its implications for rice endophyte evolution. Appl Environ Microbiol. 81:3049-3061.
- 12. Prakamhang, J., K. Minamisawa, K. Teamtaisong, N. Boonkerd, and N. Teaumroong. 2009. The communities of endophytic diazotrophic bacteria in cultivated rice (Oryza sativa L.). Appl Soil Ecol. 42:141-149.
- 13. Rangjaroen, C., B. Rerkasem, N. Teaumroong, R. Sungthong, and S. Lumyong. 2014. Comparative study of endophytic and endophytic diazotrophic bacterial communities across rice landraces grown in the highlands of northern Thailand. Arch Microbiol. 196:35-49.
- 14. Rangjaroen, C., B. Rerkasem, N. Teaumroong, R. Noisangiam, and S. Lumyong. 2015. Promoting plant growth in a commercial rice cultivar by endophytic diazotrophic bacteria isolated from rice landraces. Ann Microbiol. 65:253-266.
- 15. Santi, C., D. Bogusz, and C. Franche. 2013. Biological nitrogen fixation in non-legume plants. Ann Bot. 111:743-767.
- 16. Takeuchi, M., K. Hamana, and A. Hiraishi. 2001. Proposal of the genus Sphingomonas sensu stricto and three new genera, Sphingobium, Novosphingobium and Sphingopyxis, on the basis of phylogenetic and chemotaxonomic analyses. Int J Syst Evol Microbiol. 51:1405-1417.
- 17. Videira, S.S., J.L.S. De Araujo, L.D.S. Rodrigues, V.L.D. Baldani, and J.I. Baldani. 2009. Occurrence and diversity of nitrogen-fixing Sphingomonas bacteria associated with rice plants grown in Brazil. FEMS Microbiol Lett. 293:11-19.
- 18. Zhang, L., J.S. Gao, S.G. Kim, C.W. Zhang, J.Q. Jiang, X.T. Ma, J. Zhang, and X.X. Zhang. 2016. Novosphingobium oryzae sp. nov., a potential plant-promoting endophytic bacterium isolated from rice roots. Int J Syst Evol Microbiol. 66:302-307.
