Materials and Methods
The hind limb elements of KNM-BG 35250 are listed in Table 1. Since the pedal phalanges have been fully described elsewhere (Nakatsukasa et al., 2003), these elements are excluded from the list. Original specimens of N. kerioi and other comparative hominoids were studied in the National Museums of Kenya and the Uganda Museum. Only E. africanus specimens housed at the Natural History Museum (London) were examined using high-quality casts. Comparative extant primate samples were studied in the Osteology Department, the National Museums of Kenya, Powell-Cotton Museum, and the Anthropological Institute and Museum, University of Zurich. Since most elements of KNM-BG 35250 have been subjected to deformation, metric comparison was limited. Linear measurements were taken by using sliding calipers accurate to one-tenth of a millimeter. Angles were measured on digital photographs that were taken with settings to reduce parallax.
Table 1
Hind limb elements of KNM-BG 35250 described in this study
| Suffix |
Description |
| A |
right proximal 1/2 femur |
| B |
right distal 1/3 femur |
| D |
left proximal femur |
| E |
left proximal 1/3 tibia |
| F |
left distal fibula |
| G |
left ischium |
| H |
left distal tibia |
| I |
left talus |
| J |
left femoral shaft and distal end |
| K |
right distal fibula |
| L |
right distal tibia |
| O |
right calcaneus |
| Y |
right ischium |
| Z |
right patella proximal fragment |
| AE |
right proximal tibia |
| AF |
left tibial shaft |
| AG |
right fibula shaft |
| AH |
left fibula shaft |
| AI |
left medial cuneiform |
| AJ |
right first metatarsal proximal half |
| AK |
left second metatarsal proximal fragment |
| AL |
left fourth metatarsal proximal fragment |
| AM |
left fifth metatarsal lacking the head |
| AN |
metatarsal head with shaft |
Repeatedly used terms of anatomical orientation are abbreviated as follows: a-p = anteroposterior/-ly; m-l = mediolateral/-ly; s-i=superioinferior/-ly; p-d=proximodistal/-ly; d-v = dorsoventral/-ly; d-p = dorsoplantar/-ly.
Ischium
Description
The left ischial specimen (KNM-BG 35250G: Figure 1a) preserves the caudal half of the lunate surface and a short part of the ramus (18 mm long from the acetabulum). A fracture runs obliquely, crossing the caudal part of the acetabulum to the dorsal border of the ischial ramus around 1 cm below the ischial spine. The cross-section of the ramus is rather flat (8.8 mm thick and 22.6 mm wide at the break) probably due to postmortem d-v compression. From the acetabulum, neither diameter nor depth can be obtained reliably. The maximum width of the preserved part of the lunate surface is c. 14 mm. The ischial spine is prominent. It would have been about the level of the caudal acetabular border if the ischium were not subjected to fracture. Caudal to the spine is a well-defined marking by mm. gemelli and the tendon of m. obturator internus.

The right ischium (KNM-BG 35250Y: Figure 1b) is rather fragmentary. It is a c. 4 cm long piece, preserving the caudal part of the lunate surface and a shorter length of the ischial ramus. No additional information is added.
Comparative remarks
Where comparison is possible, the ischium of N. kerioi generally resembles that of Proconsul spp. Although it differs from that of P. nyanzae regarding a more cranial position of the ischial spine (see Ward et al., 1993), there is variability regarding this feature in P. heseloni. While it is caudally positioned in the KNM-KPS 8 individual, it is close to the caudal acetabulum rim in KNM-KPS 3. Although it was previously suggested that the lower position of the ischial spine was related to a greater development of tail muscles (McCrossin, 1994), it is now known that P. heseloni did not have a tail (Ward et al., 1991; Nakatsukasa et al., 2004). Therefore, the functional significance of this character remains an open question.
Femur
Description
Both the right and left femora consist of separated proximal and distal end pieces. The proximal piece of the right femur (KNM-BG 35250A) shows several important characters, though post-mortem cracking and deformation is apparent (Figure 2b). The left proximal piece (KNM-BG 35250D) lacks the head and most of the neck, and is crushed in the a-p direction (Figure 2c). However, KNM-BG 35250D preserves the greater and lesser trochanters, which are broken in KNM-BG 35250A. Thus, the morphology of the proximal femur is principally described based on the right piece and is supplemented by the left one. The distal piece of the right femur (KNM-BG 35250B) is crushed in the m-l direction and the left one (KNM-BG 35250J) in the a-p direction (Figure 3). The morphology of the medial and lateral epicondyles was observed in the former specimen and that of the condylar and patellar surfaces in the latter.


The head of KNM-BG 35250A displays strong anteversion. In superior view (Figure 2a), the center of the head is weakly displaced anteriorly relative to the neck axis. Furthermore, the head is strongly rotated to face anteromedially. Although a moderate head displacement is not unusual in extant anthropoids, such an emphasized rotation is anomalous and should be regarded as deformation. Both the s-i and a-p diameters of the head are 22.1 mm. Relative to the neck thickness the head is large but less so than in extant apes (Table 2), suggesting a moderately wide range of movement at the hip joint (Ruff et al., 1989; Ward et al., 1993; MacLatchy et al., 2000). The articular surface of the head is preserved except for the posterior margin. The articular surface is extensive and mushrooms over the neck except the abraded posterosuperior part (the articular border is indicated by an arrow in Figure 2d). In superior view, the articular surface depth (measured from the line passing through the anterosupeior and posterosuperior margin in Figure 2a) is 73% of the head diameter. The fovea is oblong along the a-p axis (6.1 mm a-p, 3.4 mm s-i) and situated posteroinferiorly. The head extends proximally 9 mm above the most inferior level of the superior border of the neck. By matching this specimen with the left counterpart (Figure 2d and e), it is estimated that the superior surface of the head was 1.5 mm above the tip of the greater trochanter (Table 2).
Table 2
Metric comparison of the femur
| Taxon |
|
Sex |
SIFD |
%SIFD |
GTPR |
%APFN |
%LCW |
| Nacholapithecus |
KNM-BG 35250A & D |
M |
22.1 |
159.8 |
−1.5 |
72.8 |
98.8 |
| Nacholapithecus |
KNM-BG 17821 |
M |
— |
|
— |
77.5 |
— |
| Nacholapithecus |
KNM-BG 17778 |
F |
17.6 |
|
— |
|
— |
| Proconsul nyanzae |
KNM-MW 13142A |
M |
28.7 |
166.2 |
−1.0 |
78.5 |
— |
| Proconsul nyanzae |
KNM-RU 5950 |
M |
25.3 |
157.9 |
— |
77.5 |
— |
| Proconsul nyanzae |
KNM-RU 5527 |
?M |
25.5 |
158.3 |
— |
73.4 |
76.6 |
| Proconsul nyanzae |
KNM-RU 1753 |
M |
26.7 |
— |
— |
— |
— |
| Proconsul heseloni |
KNM-RU 14227 |
F |
— |
— |
— |
— |
89.6 |
| Proconsul heseloni |
KNM-KPS 3 (right) |
F |
16.7 |
— |
— |
— |
— |
| Proconsul |
Mean |
|
— |
160.8 |
— |
76.5 |
83.1 |
| Equatorius |
BM-M16331 |
?M |
22.8 |
164.4 |
— |
76.1 |
— |
| Morotopithecus |
MUZM 80 |
? |
26.5 |
139.8 |
−1.5 |
88.1 |
81.1 |
| Ugandapithecus |
NAP IX B’64/65 |
M |
30.9 |
152.3 |
−4.0 |
77.8 |
— |
| Turkanapithecus |
KNM-WK 16950I |
?M |
17.6 |
149.7 |
−1.2 |
78.2 |
— |
| Pan troglodytes |
Mean |
F |
31.14 |
169.73 |
−2.44 |
85.29 |
81.14 |
| 9 females |
SD |
|
1.84 |
12.34 |
2.99 |
5.42 |
4.18 |
| Pongo pygmaues |
Mean |
F |
31.50 |
182.51 |
−13.50 |
73.60 |
90.85 |
| 4 females |
SD |
|
1.07 |
13.06 |
1.73 |
4.59 |
4.78 |
| Hyobates lar |
Mean |
M |
16.30 |
191.94 |
−1.75 |
82.11 |
75.67 |
| 10 males |
SD |
|
0.58 |
7.27 |
1.18 |
4.81 |
4.59 |
| Papio cynocephalus |
Mean |
M |
23.36 |
151.85 |
9.20 |
73.97 |
88.20 |
| 10 males |
SD |
|
1.06 |
9.30 |
1.55 |
2.47 |
4.87 |
SIFD: s-i diameter of the femoral head. %SIFD: relative head size (= SIFD/(neck s-i diam. × a-p diam.)0.5 × 100). GTPR: greater trochanter projection relative to the head superior surface. %APFN: relative a-p diam. of the neck (= a-p diam. of the neck/s-i diam. × 100). %LCW: relative width of the lateral condyle (= lateral condyle w./medial condyle w. × 100).
The neck-shaft angle is high (140°). This angle is comparable to that in gibbons and spider monkeys (Rose et al., 1992; MacLatchy et al., 2000). The trochanteric crest is collapsed and closes the trochanteric fossa (Figure 2d). Although this damage precludes an accurate measurement of neck length, the neck is apparently short (Figure 2b). The cross-section of the neck is moderately compressed in the a-p direction (16.2 mm s-i and 11.8 mm a-p). It is more expanded in the a-p direction in extant apes (Table 2). A large crista trochanterica is observed on the posterior aspect of the neck (arrows in Figure 2e) similar to Proconsul and Morotopithecus (MacLatchy et al., 2000).
The lateral side of the greater trochanter bears a protuberant insertion of m. gluteus minimus. The gluteal tuberosity is positioned at a relatively proximal level so that the gap between it and the insertion of m. gluteus minimus is narrow. The lesser trochanter of KNM-BG 35250D is wide s-i (16.7 mm at its base). It projects medially with weak retroflexion.
In KNM-BG 35250A, the anteromedial cortex of the shaft is cracked and widely collapsed into the medullary cavity. However, plastic deformation of the cortex seems weak. Approximate m-l diameter of the mid-shaft is 17.4 mm, which is taken at the most inferior level of the preserved cortex.
In the distal femur (KNM-BG 35250J), the patellar surface is square-shaped rather than trapezoidal in anterior view (Figure 3a arrows). The surface is wide (22.7 mm) compared with similar-sized cercopithecids (see Madar et al., 2002), which suggests an emphasis on knee rotation (Ward et al., 1995). The femoral condyles are symmetrical in width on the posterior aspect (Figure 3b). The medial and lateral condylar widths are 16.1 and 15.9 mm, respectively. If the post-mortem a-p compression occurred evenly on the femoral condyles, then this symmetry should be the original condition. On the lateral epicondyle of KNM-BG 35250B (Figure 3c), there is a large, deep, and round fovea for the lateral collateral ligament (lc). Distoposteriorly, there is a deep oblong depression for the tendon of m. popliteus (po). On the medial epicondyle (Figure 3d), there is a deep fovea for the medial collateral ligament (mc). The origin of m. popliteus is fairly developed but not groove-like as in Morotopithecus bishopi (MacLatchy et al., 2000).
Comparative remarks
The previously described proximal femora (Rose et al., 1996) exhibit a close morphological affinity to these new specimens but are more fragmentary. Newly recognized characters are as follows: head projection beyond the greater trochanter, wide articular cover of the head, moderately enlarged head relative to neck diameters, and high neck-shaft angle. The characteristics of the distal femur are all new reports.
While the femur of N. kerioi generally resembles that of Proconsul, it differs from the latter in a higher neck angle, shorter neck, somewhat better developed posterior bar of the trochanteric fossa, more medially directed lesser trochanter, more proximally located gluteal tuberosity, and square-shaped patellar surface (rather than trapezoid), although intra-specific variability in these features should also be considered (Rose et al., 1996; this study; see Ward et al., 1993 for the Proconsul femoral features). These features, except the orientation of the lesser trochanter, distinguish N. kerioi from Ugandapithecus major (Gommery et al., 1998, 2002). The femur of N. kerioi remarkably differs from that of M. bishopi. In fact, M. bishopi is distinct among the African Miocene hominoids whose femoral morphology is characterized by its small femoral head relative to the neck (Table 2) and the absence of the anterior displacement of the head (MacLatchy et al., 2000). N. kerioi further differs from M. bishopi in the developed posterior trochanteric bar, better-developed gluteal ridge, stronger projection of the m. gluteus minimus insertion, less rounded neck cross-section and more symmetric femoral condyles (see MacLatchy et al., 2000). Despite these differences, they are similar in showing the short and highly angled neck and medially projecting, proximally positioned lesser trochanter and (possibly) rectangular patellar surface. N. kerioi resembles E. africanus in the morphology of the proximal femur (Rose et al., 1996) and a square-shaped patellar surface (see McCrossin, 1994). The femur of E. africanus is long and slender, and lacks marked m-l expansion (Le Gros Clark and Leakey, 1951; Ruff et al., 1989; McCrossin, 1994). Although the femur of N. kerioi seems to lack marked shaft m-l expansion, it is uncertain if the shaft is slender like that of E. africanus. N. kerioi differs from Sivapithecus in a more emphasized platymery and m-l asymmetry of the distal femur in the latter (Madar et al., 2002).
Rose et al. (1996) interpreted that features of the proximal femur in N. kerioi are related to well-developed lesser and greater gluteal muscles and that the lesser gluteal muscles functioned as rotators as well as extensors during arboreal quadrupedalism and climbing. While engaging in these activities, the thigh must experience partial external rotation with the knee being partially flexed and pointing laterally. Newly recovered characters such as the wide articular coverage of the head, moderate expansion of the head and the high projection of the head concur with their interpretation. However, it is not readily explained why a relatively short neck is observed, which would sacrifice hip joint mobility. This is a conundrum that is also known in M. bishopi. To ensure a wide range of hip joint motion, a high neck angle should be essential for a short neck (MacLatchy et al., 2000).
The distal joint morphology complicates such a straightforward functional interpretation. An asymmetrically wider medial condyle is suggestive of emphasized loading of the medial side of the knee (Rafferty and Ruff, 1994; Mader et al., 2002). Thus, condylar width asymmetry in primates has been interpreted as an emphasis of thigh abduction in relation to arboreal quadrupedalism and/or climbing (Tardieu, 1981; Jungers and Susman, 1984; Fleagle, 1977; Fleagle and Meldrum, 1988; MacLatchy et al., 2000; Mader et al., 2002). If the knee of N. kerioi experienced habitual abduction, then condylar width asymmetry would have been accentuated similarly to Proconsul and M. bishopi (Figure 3e, Table 2). However, this is not the case.
The patellar surface morphology poses a similar question. A trapezoidal patellar surface with a proximally extended lateral side suggests that the contraction of the vastus lateralis was proportionally greater (Fleagle and Meldrum, 1988) and accords with a flexed and abducted knee, a posture suitable for a medium-sized mammal to keep balance while perched on a branch. While this is the case for in P. nyanzae (Figure 3e), the patellar surface in N. kerioi is square-shaped. To our knowledge, there is no living large-sized primate that shows N. kerioi-like knee morphology. Though admittedly speculative, these differences may reflect that the knee joint took more varied positions compared to Proconsul.
Patella
Description
The proximal half of the right patella (KNM-BG 35250Z) is preserved (Figure 4). The maximum m-l width is 22.6 mm, probably being 1–2 mm less than the true maximum width. The maximum a-p thickness is 9.8 mm. The articular surface is broad and weakly convex m-l. The proximolateral border of the superior surface forms an osteophyte projecting proximally.
Comparative remarks
Due to the fragmentary nature, KNM-BG 35250Z adds little to knowledge already obtained from a previously collected specimen (KNM-BG 15535) (Rose et al., 1996). The m-l width of KNM-BG 35250Z is approximately 1.2 times that of KNM-BG 15535, suggesting that the latter is a female specimen. Important features of N. kerioi patella are m-l wideness, a-p shallowness, and a shallow saddle-type articular surface, which permits conjunct/adjunct rotations around the a-p axis (Rose et al., 1996). This form is common with P. heseloni and E. africanus (McCrossin, 1994; Ward et al., 1995; Rose et al., 1996) and likely represents the primitive state for hominoids (Ward et al., 1995) or catarrhines (Rose et al., 1996).
Tibia
Description
The left tibia consists of three distinct pieces: 94 mm long proximal part with epiphysis (KNM-BG 35250E), 107 mm long shaft (KNM-BG 35250AF), and 30 mm long distal epiphysis (KNM-BG 35250H) (Figure 5a). The right tibia consists of 57 mm long proximal (KNM-BG 35250AE) and 27 mm long distal epiphyses (KNM-BG 35250L) (Figure 5h, e). All of these specimens were subjected to plastic deformation and cracking. Due to better preservation, the description is mainly provided for the left tibia for the proximal to middle section and for the right for the distal section.

KNM-BG 35250E is subjected to m-l compression. The medial plateau of the proximal surface is displaced posteriorly and the lateral plateau anteriorly (Figure 5c). The medial plateau is hemicircular and shallowly concave. The a-p length is 26.6 mm. The lateral plateau is ovoid and 23.3 mm along its long axis. It is concave in the m-l direction and slightly convex in the a-p direction. The tubercles of the intercondylar eminence are abraded. The lateral condyle is thick and the s-i height measures 11.7 mm at the center of the fibular facet. The fibular facet is flat and faces distally (Figure 5a). It is 12.2 mm in length and 4.8 mm in width. A moderately wide tibial tuberosity (10.4 mm) is located 10 mm below the anterior border of the proximal surface.
Although the shaft piece (KNM-BG 35250AF) does not join with the proximal and distal piece, the shape of its proximal break suggests that only a short part is missing between it and KNM-BG 35250E (Figure 5a). The lateral cortex is broadly collapsed into the medullary cavity and no useful shaft measurements can be taken. The composite length of KNM-BG 35250E, AF and H (measured excluding the medial malleolus) is 217 mm. Given that the BM of KNM-BG 35250 was 22 kg (Ishida et al., 2004), a great ape prediction model estimates 225 mm and a cercopithecoid model 231 mm (Jungers, 1984). Thus, it remains unclear which model fits better. A subadult female P. heseloni (KNM-KPS 3), with an estimated BM of 9.3 kg (Rafferty et al., 1995), has a 181 mm long tibia, which accords with an estimate based on the cercopithecoid model (179 mm) rather than the great ape model (198 mm).
The right distal tibia (KNM-BG 35250L) is subjected to post-mortem m-l compression (Figure 5d–f). The posterior cortex is partially collapsed and the groove for the tendon of m. tibialis posterior is broken. The fibular notch is wide and deep, forming an angle of 120° in distal view. Due to surface erosion and a remaining matrix layer, the articular surface of the synovial distal tibiofibular joint cannot be defined, unlike previous specimens (Rose et al., 1996). The trochlear articular surface is 19.8 mm in the a-p length. The articular surface is widest (13.8 mm) anteriorly and narrowest (10.9 mm) at the bottom of the fibular notch (these measurements are affected by deformation). A well-defined median keel divides the articular surface into the lateral and medial portions. The keel is pronounced anteriorly and ends at the median beak on the anterior border of the distal tibia. The lateral portion is wider than the medial one, especially in the anterior portion. The most anterior part of the medial portion faces distoanteriorly (Figure 5d). This part would contact the trochlear articular extension on the neck of the talus (see below) in full dorsiflexion. The lateral and medial portions are highly concave a-p and to a lesser extent m-l. Although the medial malleolus is not long, it is thick (Table 3). A thick malleolus is common in extant apes. The malleolar articular surface extends onto the anterior aspect of the malleolus, occupying the lateral half of the surface (Figure 5d), to form a large malleolar–talar cup complex (see below). In distal aspect, the attachment area of the deltoid ligament is widely excavated.
Table 3
Size and shape of the tibial malleolus. For extant taxa, average (upper) and SD (lower).
| taxon |
Height (mm) |
Width (mm) |
width/height (%) |
| Nacholapithecus kerioi KNM-BG 35250L |
9.7 |
9.3 |
95.88 |
| Proconsul heseloni |
7.83 |
6.33 |
81.14 |
| n = 4 |
0.51 |
0.13 |
6.38 |
| Proconsul nyanzae KNM-RU 1939 |
12 |
9.5 |
79.17 |
| Ugandapithecus major NAP I 1958 |
13.1 |
13.8 |
105.34 |
| Pan troglodytes |
13.06 |
12.61 |
97.75 |
| 8 females |
1.62 |
0.56 |
11.72 |
| Pongo pygmaues |
8.5 |
11.53 |
138.96 |
| 4 females |
1.8 |
0.38 |
21.39 |
| Hylobates lar |
6.38 |
6.01 |
94.56 |
| 10 males |
0.54 |
0.38 |
6.90 |
| Papio cynocephalus |
11.27 |
8.73 |
77.46 |
| 10 males |
0.63 |
0.62 |
3.67 |
Proconsul heseloni: KNM-RU 2036BA, 3589, 18390, KPS 3 (TB6).
Two other tibial specimens add little useful information. The right proximal piece (KNM-BG 35250AE) is smashed m-l (Figure 5h). The intercondylar eminences are prominent, probably due to post-mortem deformation. There is a deep pit on the medial side of the medial condyle, which is likely the insertion of m. semimembranosus (Figure 5h). The left distal epiphysis (KNM-BG 35250H) is strongly compressed a-p (Figure 5g).
Comparative remarks
The distal tibia of N. kerioi generally resembles that of Proconsul (e.g. KNM-RU 1939 P. nyanzae; Figure 5i, j). Although few proximal tibial specimens exist, a thick lateral condyle is also observed in P. heseloni (e.g. KNM-KPS 3). Features of the talocrural articular surface (salient median keel, distoanteriorly facing facet) provide joint stability in the close-packed position (Conroy and Rose, 1983). The talocrural joint of these fossil apes lacks the specialized mobility that living apes attain, and is more like that of cercopithecoids in this regard (Conroy and Rose, 1983). N. kerioi differs from Proconsul in the greater thickness of the medial malleolus. This trait is probably related to emphasized medial loading during inversion (DeSilva, 2009), and suggests a greater proportion of climbing in its total locomotor behavior. The very high value in U. major (Table 3) is probably size-related.
Fibula
Description
The left fibula is represented by two separate pieces: 92 mm long shaft fragment (KNM-BG 35250AH) and 76 mm long distal fragment (KNM-BG 35250F) (Figure 6a–c). The right fibula is represented by a 111 mm long shaft (KNM-BG 35250AG) and a 19 mm long distal fragment (KNM-BG 35250K) (Figure 6d–f).
Both of the shaft pieces are flat m-l throughout and have a sharp anterior border and a rounded posterior border. Medially, a 3–4 cm long interosseous border runs distally from the proximal break in both specimens. Thus, these specimens preserve almost identical parts, with the right one preserving distally a greater amount of the shaft. The right shaft is free from plastic deformation. It is widest a-p 4 cm above the distal break (11.0 mm). Comparing it with a well-preserved fibula of P. heseloni (KPS-FB 5), this appears to correspond to the original maximum a-p diameter. At this level, the m-l diameter is 7.3 mm.
In the (left) distal fibula (KNM-BG 35250F) the anterior border of the shaft changes course laterally as it descends distally, and approaches the apex of the fibular malleolus (Figure 6c). This leaves a triangular subcutaneous area on the anterior aspect of the bone. Medially, the attachment site of the anterior inferior tibiofibular ligament is prominent. The maleollus is long and projects distally and slightly laterally. The malleolar fossa is deep. The talar facet morphology is affected by post-mortem a-p compression. However, it otherwise appears similar to that of previously reported specimens (Rose et al., 1996).
Comparative remarks
The fibular shaft and distal joint features in N. kerioi are largely comparable with Proconsul and A. turkanensis (Walker and Pickford, 1983; Rose et al., 1996; Leakey et al., 1988; Walker, 1997; Ward, 1998; Ward et al., 1993). Notable features are a flat but very robust shaft and large fibular maleollus. These characters are related to the development of m. flexor hallucis longus and ligamentous reinforcement of the talocrural joint, in relation to a strong hallucal grasp (Walker and Pickford, 1983; Rose et al., 1996).
Talus
Description
The left talus (KNM-BG 35250I) is well preserved (Figure 7), although some damages are noted. The lateral tubercle of the posterior talar body is broken off. The medial tubercle is dorsally displaced. The talar head is dorsoplantarly compressed, resulting in m-l expansion and flattening of the distoplantar surface. Cortical cracks on the middle and posterior articular surfaces of the talocalcanean joint suggest that these regions also suffered from post-mortem compression.

The trochlea is 15.5 mm in anterior width and 15.2 mm at mid-width (measurements following Rose et al., 1996). The a-p length is 23.2 mm along the lateral trochlear rim. The trochlea is moderately long (or narrow): width/length ratio (index 1 in Rose et al., 1996) is 70.7%. It is only faintly wedged posteriorly (wedging index = 96.2%; index 3 in Rose et al., 1996) with the medial and lateral trochlear rims virtually being parallel (Figure 7a). The trochlear groove is deep, and the lateral rim is much higher than the medial one. Thus, the lateral slope of the trochlear groove is far wider than the medial slope (Figure 7e). Although the medial rim might be lowered by post-mortem compression, the asymmetry of the distal tibial joint (Figure 5) indicates that the medial rim was originally lower than the lateral rim. In dorsal view, the lateral rim projects more distally than the medial rim. The medial rim runs distomedially and continues to the anterior border of a cup-shaped depression (malleolar cup) on the neck (Figure 7a). The malleolar cup is large. Lateral to the malleolar cup and beyond the medial trochlear rim is a wide depression, which is reciprocal for the median beak on the anterior border of the distal tibia.
The lateral surface of the trochlea is moderately tall. This surface is vertical except for the triangular-shaped inferior part (Figure 7c). Posterior to the lateral articular surface is a deep pit for the posterior talofibular ligament. The trochlear height measured at the most distal level of the lateral rim is 12.8 mm. The trochlear height/length ratio (56.9%) is common in anthropoids (index 2 in Rose et al., 1996). The medial tubercle of the posterior talar body is very large (Figure 7b) as seen in some atelines (Alouatta, Lagothrix). This tubercle guides the tendon of m. flexor hallucis longus medially and bears the attachment of the posterior tibiotalar ligament.
On the plantar aspect, the posterior surface of the talocalcanean joint is 17.5 mm long along the long axis and concave to a degree common in extant non-hominoid catarrhines. Its anterior margin is beveled distoanteriorly, to contact with the calcaneal body in the close packed position. The sulcus tali is deep, and it was certainly deeper before the fractures of the posterior and middle calcanean articular surfaces (Figure 7b: st). Medial to the distal articular surface of the talocalcanean surface is a deep pit for the talonavicular ligament (tn).
The neck deviates medially, forming an angle of 144° with respect to the medial trochlear rim. The neck is moderately wide (13.8 mm at the narrowest part) (index 4 (Harrison, 1982) = 59.5%). The relative neck length (index 3 (Harrison, 1982)) is also intermediate (65.9%). The head is very wide (18.3 mm) with the relative width (index 5 (Harrison, 1982)) of 78.9%. However, this is exaggerated by post-mortem deformation. The original size of the head is unknown.
Comparative remarks
Previously, the talar morphology of N. kerioi was reported based on KNM-BG 15529, a weathered talar trochlea (Rose et al., 1996). The trochlea of KNM-BG 35250I generally resembles this specimen (Figure 8), except its d-p taller form. This difference resulted from post-mortem deformation in KNM-BG 15529.
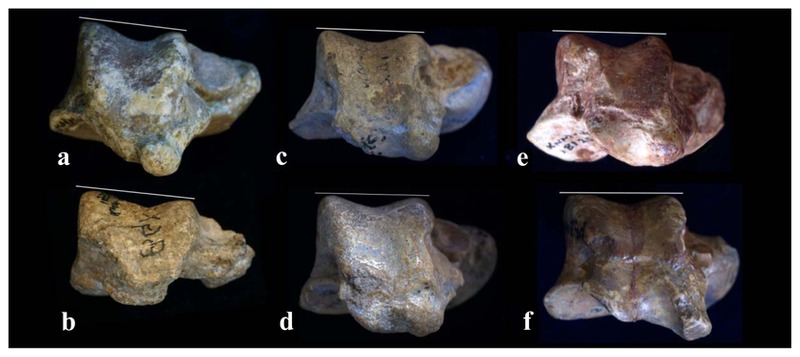
The talar morphology is generally homogenous in African Miocene apes (Harrison, 1982; Conroy and Rose, 1983; Langdon, 1986) and N. kerioi is not exceptional. The trochlea is a subtly asymmetric trapezoid with weak posterior wedging, rather than the asymmetric quadrangle with strong posterior wedging observed in extant African apes (Ward, 1998). It is neither very long nor very wide (Rose et al., 1996). The trochlear groove is deep being similar to that of cercopithecoids, though differing from the shallow morphology as observed in extant African apes (Harrison, 1982; Conroy and Rose, 1983; Langdon, 1986). The lateral rim is higher than the medial rim (Harrison, 1982). Like cercopithecoids, the malleolar cup is well developed and a concave tibial stop is formed at the distal border of the trochlear groove (Le Gros Clark and Leakey, 1951; Harrison, 1982; Conroy and Rose, 1983; Ward et al., 1993). The neck orients medially, more than orangutan, platyrrhines and prosimians, but less than gorillas and humans (Harrison, 1982). Since these features are generally common among Pliopithecus and small-sized non-cercopithecoid catarrhines from the Miocene of Africa, these represent the primitive catarrhine condition (Conroy and Rose, 1983; Langdon, 1986; Rose, 1993; Ward et al., 1993). In these fossil African apes, inversion of the talocrural joint is intermediate between extant cercopithecoids and great apes, and conjunct inversion and abduction occur during dorsiflexion like the cercopithecoid ankle (Conroy and Rose, 1983). The tibial stop in the distal trochlea, and the maleollar cup, provide the joint with stability in full dorsiflexion, which might have been taken in the propulsive thrust toward the end of the stance phase in quadrupedal walking and/or climbing. This basic foot structure is representative of extant monkeys and apes, and was certainly suitable for a wide range of generalized arboreal activities (Conroy and Rose, 1983).
However, there is some (relatively minor) variation among these fossil hominoids. In N. kerioi, the asymmetry of the trochlea groove is more emphasized than in Proconsul and other African fossil hominoids due to a more prominent lateral trochlear rim (Figure 8), which may be related to effective weight transfer during inversion. The medial tubercle of the posterior talar body is remarkably large, suggesting greater development of the long hallucal flexor, which must be associated with the unusually large size of the hallux (Nakatsukasa et al., 2003). The implication is that N. kerioi engaged in climbing or other antipronograde activities more frequently compared to Proconsul.
Calcaneus
Description
The distal segment of the right calcaneus (KNM-BG 35250O) is preserved (Figure 9). The proximal break crosses the calcaneal body obliquely from the posterior base of the sustentaculum tali to the distolateral corner of the posterior talar facet (PTF). The sustentaculum tali is intact. The margin of the cuboid facet is eroded with the exception of the plantarmedial part.

The maximum breadth of the body (Harrison, 1982) is 25.4 mm. Although the distal segment (distal from the anterior border of the PTF) appears somewhat short, this is an artifact of erosion on the cuboid facet. The middle talar facet (MTF) on the sustentaculum is continuous with the anterior talar facet (ATF) via a narrow articular band (isthmus). The breadth of the MTF is 6.8 mm and the combined MTF/ATF length is 19.6 mm. The relative width of the MTF is as low (34.7%) as macaque and gibbon (index 7 in Rose, 1986). The MTF/ATF region is strongly curved in medial view (Figure 9b). Lateral to the articular isthmus is a distinctively deep (>2 mm) pit for the interosseous talocalcaneal ligament (ITL). Under the sustentaculum, there is a quite deep groove for the tendon of m. flexor hallucis longus. This condition is accentuated by the lateral expansion of the distomedial planter tubercle, the site of origin for the short plantar ligament and most fibers of the spring ligament.
Comparative remarks
KNM-BG 35250O differs from the previously described calcaneus (KNM-BG 17805; Rose et al., 1996) (Figure 9d, e) in the height of the distal body and the deep pit formation for the ITL. However, the shallowness of the distal calcaneal body in KNM-BG 17805 is attributed to post-mortem deformation (see below). In KNM-BG 17805, the attachment of the ITL is a shallow depression.
There is additional calcaneal material from the KNM-BG 15532 specimen, which was collected in 1984 and never reported (Figure 9f). KNM-BG 15532 is a distal half of a small right calcaneus, missing the calcaneal tuberosity, sustentaculum tali, and medial half of the cuboid facet. The middle and distal segment length (distal from the posterior border of the PTF) is 24.5 mm. This specimen is likely a small female or an immature male. It resembles KNM-BG 17805 in the morphology of the medial plantar process (only its anterior portion is preserved) and KNM-BG 35250O in general morphology except for a shallower groove for the tendon of m. flexor hallucis longus (probably size-related) and the absence of a deep ITL pit. The PTF of KNM-BG 15532 is almost intact. It is oval in outline (12.3 mm long and 8.1 mm wide) with its long axis being modestly angulated proximomedially to distolaterally. The relative width of 65.9% is moderate (Rose, 1986). The PTF is tightly curved along the long axis. The degree of the curvature is tightest at the midpoint. The cuboid facet is broken dorsomedially. The peripheral (lateral and dorsal) part of the cuboid facet is fairly wide and flat, resembling calcanei of extant African apes (Rose, 1986).
These features are generally common in other African fossil hominoid calcanei. The subtalar articulation probably had a greater range of motion than in extant cercopithecids but received habitual loading in either inversion or eversion, without large weight-bearing in transitional position (Conroy and Rose, 1983; Harrison, 1982; Rose, 1986; Rose et al., 1996; Ward et al., 1993). A good development of the medial plantar process suggests well-developed intrinsic toe flexors, which would facilitate toe grasp even during foot dorsiflexion (Sarmiento, 1983; Rose et al., 1996).
However, a deep pit for the ITL attachment site observed in KNM-BG 35250O is rare in fossil apes and living catarrhines. In Proconsul, its morphology varies from a shallow depression to a rugose surface. U. major (KNM-SO 390) shows a somewhat similar pit, but much less deep. This site is merely a rugose surface in Sivapithecus (Rose, 1986). Although a deep pit is observed in only one out of the three N. kerioi calcanei, none of the other African Miocene ape calcanei (about 10) show a similar feature. More N. kerioi specimens are needed to evaluate the frequency and expression of this trait.
Medial cuneiform
Description
The left medial cuneiform (KNM-BG 35250AI) is almost intact (Figure 10). The a-p length is 18.8 mm. The d-p length is 21.8 mm. The relative length (Harrison, 1982) is 86.2%. The m-l width is 10.5 mm. The margin of the proximal (= navicular) articular surface is partly abraded. It is elliptical (9.4 mm wide and ~12 mm long).
The hallucal metatarsal (MT1) articular surface is 18.6 mm high. It is reniform and highly convex m-l (Figure 10b). The MT1 articular surface in N. kerioi is not a simple ellipse. Its main articular region is a bulged band which spirals on the medial and distal sides of the bone superomedially to inferodistally. However, along the plantar articular border, the peripheral region curves in dorsally (upward). Therefore, the plantar third of the whole articular surface takes seller form.
On the lateral side, there are proximal and distal facets for the intermediate cuneiform, and the MT2 facet. The latter facet is continuous with the distal facet for the intermediate cuneiform.
Comparative remarks
The medial cuneiform of N. kerioi is similar to that of Proconsul in overall shape. However, the prehallux articulation, which is present in Proconsul, is absent in N. kerioi. The prehallux participates in the medial cuneiform-MT1 joint medioplantarly. The presence of this bone is a primitive feature retained in platyrrhines and gibbons (Lewis, 1972; Conroy, 1976; Harrison, 1982). When the MT1 is fully abducted on the medial cueniform, a bare joint surface remains on the medioplantar part of the articular surface in Proconsul, which would serve for the prehallux (Figure 10f). There is no equivalent surface in N. kerioi. An absence of the prehallux is also evidenced from the MT1 morphology (see below). However, the occurrence and development of the prehallux are rather variable individually and ontogenetically, and its utility for functional and phylogenetic consideration is questioned (Wikander et al., 1986). The medial cuneiform in N. kerioi is unusually large for its body mass being approximately equal to that of P. nyanzae (Figure 10). The large size is unquestionably related to the large hallux.
Metatarsals
Five MT specimens are associated with KNM-BG 35250. Four of them preserve the proximal joint. The remaining specimen is a non-hallucal MT head with a short piece of the shaft and cannot be assigned to a particular ray.
KNM-BG 35250AJ is the right MT1. The head is missing along with the distal shaft (Figure 11a, b). The preserved length is 39 mm. The shaft is squashed d-p. (Note orientations are defined by the human standard anatomy and differ from those in articulated position (= in opposition) here.) Likewise, the epiphysis is compressed d-p to a lesser degree (Figure 11c). Due to this compression, the proximal articular surface is lower d-p and more tightly curved than it would have been. The articular surface is reniform. There is no articular extension on the medial side of the bone, which would contact with the prehallux if it exists (see Figure 11d).

The MT1s of African Miocene apes are generally uniform. It is large and robust and the proximal joint ensures good abduction–adduction mobility (Harrison, 1982; Walker and Pickford, 1983; Ward et al., 1993). The sole exception is E. africanus, which is reported to have a reduced articular surface of the proximal joint and suggested to have had a limited range of abduction (McCrossin, 1994).
A 20 mm long proximal part of the left MT2 (KNM-BG 35250AK) is preserved (Figure 11e, f). The proximal articular surface measures 11.4 mm m-l and 12.9 mm d-p. On the lateral side of the shaft, there are articular surfaces for the lateral cuneiform and MT3. On the medial side, there is a large rectangular articular surface for the medial cuneiform (8.8 mm high, 5.2 mm long). The broken end of the shaft is ovoid, being 6.9 mm m-l and 9.1mm d-p. This specimen is similar to that of Proconsul, particularly KNM-RU5872P. The size of the epiphysis is also equivalent (11.2 mm, 12.5 mm). However, the shaft is more slender compared with P. nyanzae (8.2 mm m-l and 10.2 mm d-p at the comparable level).
The left MT4 (KNM-BG 35250AL) is a 30 mm proximal part (Figure 11g, h) and is squashed m-l. The cortex of the shaft is cracked and collapsed into the medullary cavity. The epiphysis is also compressed and the articular surfaces for the MT3 are flattened. The lateral surface bears an extensive, concave articular surface for the MT5 (Figure 11g).
The left MT5 (KNM-BG 35250AM) is broken proximal to the head (Figure 11k, l). The preserved length is 62.4 mm. The proximal articular surface is triangular in shape, wide (10.2 mm in width and 7.3 mm in medial height), and highly convex d-p, which suggests hyperdorsiflexion capabilities (Figure 11k). On the medial side, there is a large crescent articular surface for the MT4. The dorsal horizontal part is long and faces dorsomedially. The styloid process is obtuse, but projects proximolaterally, with the insertion of the m. peroneus brevis located far laterally. Proximomedially on the plantar side, there is a tubercle for the origin of m. flexor digiti minimi brevis (Figure 11l, arrow). Between this tubercle and the styloid process, there is a groove for m. abductor digiti minimi. The shaft is slender. The cross-section is d-p long and elliptical. At approximately mid-level, it is 5.0 mm m-l and 5.8 mm d-p. This specimen closely resembles KNM-RU 5872H P. nyanzae specimen (Figure 11m). The latter specimen is slightly larger in overall size and the shaft is more robust.
A 3.0 cm long distal end of a metapodial (KNM-BG 35250AN) of a lateral ray has also been collected (Figure 11i, j). The head is squashed m-l. Even if deformation is taken into account, the narrowness of the head is more reminiscence of a MT rather than a metacarpal (e.g. KNM-BG 17807; see Rose et al., 1996). Apart from the head proportion, its articular features are common with those of the metacarpal (which is also true for P. heseloni). The pits for the collateral ligaments are located close to the mid-line so that the articular surface is pinched dorsoproximally. On the plantar side, the articular surface is more extensive proximally near the medial and lateral sides, which accommodate the sesamoid bones. However, “fluting” to separate sesamoid areas from the central part (Lewis, 1989) is absent. This suggests that the metatarsophalangeal joint allowed higher degrees of abduction–adduction and rotations, though not as extensively as is in extant great apes (Rose et al., 1996). The shaft is circular in cross-section and thicker than the distal shaft of KNM-BG 35250AM (6.2 mm d-p and 6.1 mm m-l at the break).



