Abstract
Two complete dog skeletons were recovered during archeological excavations from 1961 to 1970 at the Kamikuroiwa rock shelter, a site that yielded a series of cultural entities from the Late Pleistocene, Incipient Jomon, and Early Jomon periods. Since two dogs were buried close to human skeletons, it was thought that these dogs had been buried by Jomon people, and hence provided the oldest direct evidence of Canis domestication in Japan. However, the stratigraphic information and archeological contexts of these dog skeletons are incomplete due to the lack of detailed excavation reports and technical limitations of excavations at this site. Because the date of the dog burials has not been fully discussed in the context of modern chronology or recent discussions on Canis domestication, we directly measured radiocarbon ages and stable isotope analysis on two dog burials and one set of human remains from the Kamikuroiwa rock shelter. These data are important for reconstructing the relationship between humans and dogs in the Jomon period. Our results show that the human thought to have been buried with the dogs was assigned to the middle Initial Jomon period (8977–8725 calBP), whereas, on the other hand, dates for the dog burials are very close to each other and were assigned to the latest Initial Jomon or the initial Early Jomon periods (7414–7273 calBP). Although these results are not consistent with previous archeological interpretations for this site, they remain important because these two dog burials are among the oldest evidence of Canis domestication in East Asia.
Introduction
Among animals, the dogs have the longest history of living with humans. Their role as companion animal and in hunting is different from other livestock (e.g. goat, sheep, cattle, and horse), which are mainly exploited for food or labor. Research on the origin and process of dog domestication has mainly followed three approaches: genetic analysis of modern dogs and wolves, morphological analysis of excavated dog skeletons, and the archeological evidence indicating artificial burials (Larson et al., 2012).
Genetic findings suggest that dog domestication occurred in East Asia between 14 and 16 kya (thousand years ago), based on the phylogenetic analysis of mitochondrial and nuclear DNA among dogs in East Asia (Vila et al., 1999; Savolainen et al., 2002; Lindblad-Toh et al., 2005; von Holdt et al., 2010). On the other hand, recent studies based on different phylogenetic analyses revealed that the domestication event occurred independently in East Asia and West Asia at around 30 kya or earlier (Ding et al., 2012; Wang et al., 2013).
Dog domestication is also evident in fossil remains worldwide. According to osteological analysis on dog remains in West Asia and Europe, it was shown that the size of dog mandibles and lower teeth had became significantly smaller than those of wolves. The dates of these specimens were reported as Late Pleistocene from Northern Europe and the Levant, c. 14000 calBP (Benecke, 1987; Dayan, 1994). Other researchers have claimed that some incipient domesticated dogs existed in East and West Asia in the final part of the Pleistocene, based on morphological characteristics of Pleistocene dog remains (Payne, 1983; Nikolai et al., 2011). However, it is a challenging task to distinguish the incipient dog from extinct canids in the Late Pleistocene, because the morphological characteristics of the incipient dog might show similarities with those of wild canids which had wider variability in time and space (Macdonald and Sillero-Zubiri, 2004).
In contrast to identification of canid domestication by bone morphology, significant evidence of domestication has been provided by archeological contexts suggestive of intentional burial and special treatment. For example, canids buried in human tombs have been found at several sites in West Asia and Europe between c. 14000 and 11500 calBP. These individuals were considered domestic dogs, although they were incomplete skeletons isolated from a burial context (Davis and Valla, 1978; Benecke, 1987; Pionnier-Capitan et al., 2011). In fact, the number of complete dog skeletons is quite limited even in the global discussion of Canis domestication. Exceptionally, at the two oldest sites in China, Jiahu (9000–7800 calBP) and Dadiwan (7560–7160 calBP), one and four complete dog skeletons were reported, respectively (Li et al., 2003; Barton et al., 2009).
In Japan, a number of canid remains have been excavated from prehistoric sites, suggesting that the oldest domestic dog probably dates to the Initial Jomon period (Komiya, 2003). In the Initial Jomon sites, three important dog remains are well known, including those from the Natsushima shell midden located at Yokosuka City in Kanagawa Prefecture; the Mizuko shell midden located at Fujimi City in Saitama Prefecture; and as reported herein, the Kamikuroiwa rock shelter located at Kuma-kogen Town in Ehime Prefecture (Shigehara and Hongo, 2000; Sugawara, 2003). Among these sites, the dog remains from the Natsushima shell midden were excavated from the first midden dated to 12117–9281 BP (uncalibrated), which is regarded as the oldest ‘Jomon dog.’ This dog specimen was a fragmentary mandible not preserved in anatomical position (Sugihara and Serizawa, 1957; Komiya, 2003). In contrast, the two dog remains found at Layer 4 of the Kamikuroiwa rock shelter were completely preserved in skeletal articulation. Associated artifacts were also found, including Oshigatamon pottery (Kijima type) assigned to the middle Initial Jomon period. In addition, considering that several human remains were excavated from the grid near the two dog skeletons, it was believed that these two dogs were intentionally buried by Jomon people (Esaka et al., 1967; Anezaki et al., 2009). Hence these two dog individuals from the Kamikuroiwa rock shelter represent the oldest domesticated dogs in Japan among prehistoric canid remains with clear contexts for intentional burials.
However, the chronological determinations of the Kamikuroiwa dogs are very problematic. The date of the dog burials could not be precisely determined because their archeological context was not clearly described in the excavation report published in 1967. According to Esaka et al. (1967), these dog burials were clearly associated with human remains and Oshigatamon pottery assigned to the Initial Jomon period (Layer 4) because “these dogs were excavated in the position protecting human remains,” although Todoroki-type pottery (the latest Initial to the earliest Early Jomon) was recovered directly above the dog remains.
In previous studies of prehistoric dogs in Western countries, direct radiocarbon dating of dog remains has been a powerful tool (Protsch and Berger, 1973). In contrast, few direct dates for prehistoric dog remains of the Jomon period have been reported, because direct dating is usually applied to associated charcoal in Japanese archeology. However, the radiocarbon dates obtained from charcoal can be quite different from those from bone. In particular, in a complicated deposit of caves or rock shelters, such as Kamikuroiwa, direct dating of dog remains is needed for a more concrete discussion. Therefore, in this study, we evaluated the archeological context of the two dog burials and one human skeleton by radiocarbon dates directly obtained from bone samples. Furthermore, we discuss the relationship between domestic dogs and human populations in the Initial and Early Jomon periods with comparisons of dietary signatures based on stable carbon and nitrogen isotope analysis.
Materials and Methods
The Kamikuroiwa rock shelter was excavated from 1961 to 1970 using a grid system consisting of units named A–F and ‘expanded’ grids. The rock shelter deposits were stratigraphically divided into nine layers, which included a series of artificial relics assigned to the Initial Jomon period to the present (Esaka et al., 1967). Esaka et al. (1967) reported that two complete dog skeletons were recovered from the top of Layer 4 in Grid A, and assigned them to the Initial Jomon period because this lower layer yielded Oshigatamon pottery. However, an excavation note points out that the dog burials were located at the top of Layer 4 in Grid A, and pot shards of the Todoroki type, which were mainly recovered from Layer 3, were found just above the dog remains. Considering these archeological contexts, it is possible that the dog burials belonged to Layer 3. In contrast, ten human remains were found in Layer 4 in the expanded Grid A neighboring Grid A, and were assigned to the Initial Jomon period despite being associated with Oshigatamon pottery (Kijima type) of the middle Initial Jomon period (Kobayashi and Onbe, 2009; Onbe, 2009).
The dog remains (axis, cortical parts without spongious parts) analyzed in this study comprise Dog No. 1 and Dog No. 2 (Figure 1). Because these remains constituted complete skeletons that were clearly smaller than other large canines (e.g. wolf and extinct wild canines), the two individuals were identified as domesticated dogs (Komiya et al., 2015). Additionally, the buried human remains (No. 11, adult woman) stored at the Kamikuroiwa Rock Shelter Museum in Kuma-kogen Town, Ehime Prefecture, were analyzed for this study. These human remains were excavated from Layer 4 in the expanded Grid A. In addition, the carbon and nitrogen isotope ratios of ten zooarchaeological remains (one giant flying squirrel, one marten, two badgers, two wild boars, and four sika deers) were measured to evaluate the feeding habitats of the dogs and the human.

Based on previous studies on how to extract bone collagen, c. 0.5 g of cortical bones was sampled from faunal and human remains using a diamond cutting tool (Longin, 1971; Yoneda et al., 2002). To remove contamination of organic matter from soil, the bone surfaces were cleaned using a sand blaster and an ultrasonic cleaning device. The washed samples were reacted in 0.2 M NaOH at 4°C for 12 h. After neutralization and drying the reacted samples, they were pulverized using a mortar and pestle. The sample powders were decalcified in 1.2 M HCl at 4°C for 12 h in a cellulose tube, and were subsequently gelatinized in distilled water acidified to pH 3.0 at 90°C for 12 h. The gelatinized samples were cleaned using a glass filter (GF/F, Wattman) and the dissolved gelatin was recovered by freeze drying.
To measure the atomic ratios of carbon and nitrogen (C : N) and δ13C and δ15N values, extracted collagen (c. 500 μg) was analyzed using an elemental analyzer (Flash 2000, Thermo Fisher Scientific) coupled to an isotope ratio mass spectrometer (EA-IRMS; DELTA V and ConFloIII, Thermo Fisher Scientific). The isotope ratios were calculated based on the difference from reference materials of carbon and nitrogen (PDB and AIR, respectively) in per mille (‰) units. Commercial standard alanine (No. AZ1Z0, Lot. M8A 0384, SI Science) that can be traced back to PDB and AIR were used to correct isotope ratios of unknown samples and evaluate measurement error. In this study, a measurement precision of 0.1‰ for the carbon and nitrogen isotope ratios was taken as one standard deviation.
To synthesize graphite for radiocarbon dating, CO2 gas from the gelatin was purified, reduced to graphite with hydrogen over iron powder in a vacuum line (Minagawa et al., 1984; Kitagawa et al., 1993). The 14C content in the synthesized graphite was measured by accelerator mass spectrometry (AMS) performed at the Paleo Labo Co. Ltd. (PLD) and the University of Tokyo (MALT) (Kobayashi et al., 2007; Matsuzaki et al., 2007). The 14C count was corrected using international standard materials IAEA C1, SRM4990C, and ANU C6.
In addition, the preparation and radiocarbon measurement were independently replicated at the Research Laboratory for Archaeology and the History of Art (RLAHA), University of Oxford. The second samples, taken from next to the first sampling positions, were pretreated by ultrafiltration, and the radiocarbon dating was carried out at RLAHA on a commercial basis (Ramsey et al., 2004).
The 14C content was corrected by δ13C measured by AMS. Conventional 14C age (BP) was calibrated to the calendar year (calBP) using the calibration program OxCal 4.2 with the IntCal 13 dataset (Ramsey, 2009; Reimer et al., 2013).
Results
The human and animal remains from Kamikuroiwa produced enough collagen for analysis (Table 1). To evaluate the effect of contamination and diagenetic alteration, the carbon and nitrogen content (%C and %N) in collagen was measured to evaluate the atomic C : N ratio. All samples have C : N ratios within the range of modern ones (2.9–3.6) (Deniro, 1985; Ambrose, 1990). Furthermore, the correlation between C : N ratios and δ13C or δ15N values (Spearman’s test, P > 0.05) suggests that the effect of contamination and diagenetic alteration on radiocarbon dating and stable isotope analysis is quite limited (van Klinken, 1999).
Table 1
Result of collagen extraction and elemental analysis from Kamikuroiwa rock shelter samples
| Sample ID |
species |
Bone (mg) |
Gelatin (mg) |
Gelatin yield |
Carbon content (%) |
Nitrogen content (%) |
Atomic C/N |
δ13CPDB |
δ15NAIR |
Analysis ID (EA-IRMS) |
| Dog (No. 1) |
Canis familiaris |
210.4 |
7.6 |
3.6 |
44.3 |
15.7 |
3.3 |
−20.8 |
8.1 |
YL0047 |
| Dog (No. 2) |
Canis familiaris |
242.7 |
18.3 |
7.5 |
44.5 |
16.0 |
3.2 |
−20.6 |
8.5 |
YL0048 |
| UT092301 |
Homo sapiens |
228.8 |
6.4 |
3.1 |
42.2 |
15.0 |
3.3 |
−20.8 |
7.6 |
— |
| KR9801 |
Meles meles |
215.9 |
3.2 |
1.5 |
43.1 |
14.9 |
3.4 |
−19.8 |
4.9 |
YL04815 |
| KR9802 |
Sus scrofa |
373.1 |
12.8 |
3.4 |
44.4 |
15.3 |
3.4 |
−19.7 |
3.0 |
YL04816 |
| KR9803 |
Sus scrofa |
361.1 |
18.5 |
5.1 |
43.2 |
14.8 |
3.4 |
−21.2 |
4.3 |
YL04817 |
| KR9809 |
Cervus nippon |
296.0 |
25.9 |
8.8 |
45.0 |
15.5 |
3.4 |
−21.2 |
3.4 |
YL04818 |
| KR9810 |
Cervus nippon |
466.2 |
38.8 |
8.3 |
44.9 |
15.3 |
3.4 |
−21.2 |
3.4 |
YL04819 |
| KR9811 |
Cervus nippon |
380.6 |
32.7 |
8.6 |
44.3 |
15.7 |
3.3 |
−23.8 |
4.1 |
YL04820 |
| Deer (No.1) |
Cervus nippon |
527.0 |
42.3 |
8.0 |
45.0 |
16.1 |
3.3 |
−21.0 |
6.7 |
YL0049 |
| KR9820 |
Lepus brachyurus |
211.9 |
3.4 |
1.6 |
45.3 |
15.4 |
3.4 |
−25.0 |
3.1 |
YL04821 |
| KR9821 |
Petaurista leucogenys |
196.5 |
4.1 |
2.1 |
43.2 |
14.3 |
3.5 |
−19.4 |
2.3 |
YL04822 |
| KR9904 |
Martes melampus |
232.2 |
6.3 |
2.7 |
42.1 |
13.9 |
3.5 |
−18.9 |
5.5 |
YL04824 |
| KR9905 |
Martes melampus |
241.4 |
7.1 |
2.9 |
42.9 |
14.0 |
3.6 |
−18.8 |
6.0 |
YL04825 |
Regarding the uncalibrated radiocarbon ages of Dog No. 1 and Dog No. 2, results from the PLD were 6326 ± 42 BP (PLD-21149) and 6291 ± 38 BP (PLD-21150), respectively. The 14C date of the No. 11 human associated with the dog burials was 7958 ± 48 BP (MTC-13800), which is significantly different from the dates of the dogs (Table 2). The 14C dates replicated by RLAHA were 6557 ± 36 BP for Dog No. 1 (OXA-26798) and 6427 ± 34 BP for Dog No. 2 (OXA-26799), which are slightly older than results from the PLD (Table 2). However, both results indicated that the dog burials were clearly younger than the human remains, although the dogs were previously assumed to have been buried with the human.
Table 2
Result of radio carbon dating for the human and buried dogs in Kamikuroiwa rock shelter
| Sample Name |
Analysis ID (AMS) |
14C Age (yrBP ± 1σ) |
Calibrated Age 1σ (calBP) |
Calibrated Age 2σ (calBP) |
Note |
| Dog (No. 1) |
PLD-21149 |
6326 ± 42 |
7308–7241 (48.6%)
7213–7178 (19.6%) |
7412–7395 (2.1%)
7369–7360 (0.9%)
7329–7165 (92.4%) |
Burried Dog |
| Dog (No. 2) |
PLD-21150 |
6291 ± 38 |
7259–7231 (25.2%)
7225–7175 (43.0%) |
7306–7160 (95.4%) |
Burried Dog |
| Dog (No. 1) |
OxA-26798 |
6557 ± 36 |
7481–7430 (68.2%) |
7560–7540 (5.9%)
7514–7422 (89.5%) |
Ultrafileration |
| Dog (No. 2) |
OxA-26799 |
6427 ± 34 |
7416–7349 (54.4%)
7340–7323 (13.8%) |
7424–7280 (95.4%) |
Ultrafileration |
| Dog (No. 1) |
— |
6442 ± 56 |
7422–7322 (68.2%) |
7440–7261 (95.4%) |
Mean of two laboratories |
| Dog (No. 2) |
— |
6359 ± 51 |
7413–7392 (8.7%)
7372–7357 (5.4%)
7331–7249 (54.1%) |
7419–7235 (84.9%)
7222–7175 (10.5%) |
Mean of two laboratories |
| Dog (No. 1 + No. 2) |
— |
6397 ± 38 |
7414–7390 (15.7%)
7373–7356 (10.5%)
7332–7273 (42.0%) |
7420–7266 (95.4%) |
Mean of two dogs |
| Human (No. 11) |
MTC-13800 |
7958 ± 48 |
8977–8879 (30.2%)
8871–8826 (14.0%)
8807–8725 (24.0%) |
8990–8647 (95.4%) |
Human remain (No.11) |
Subsequently, stable isotope analysis was done on the same bone collagen as the radiocarbon dating. The carbon isotope ratios (δ13C) were −20.8‰ and −20.6‰, and the nitrogen isotope ratios (δ15N) were 8.1‰ and 8.5‰ for Dog No. 1 and Dog No. 2, respectively (Table 1). Considering that the δ13C value was −20.8‰ and the δ15N value was 7.6‰ for the human remains, the isotope signatures of both dogs were similar to that of the human (Figure 2). In addition, the δ13C and δ15N values of the dogs and human were higher than those of herbivores (deer, wild boar, and giant flying squirrel; −21.1 ± 1.5‰ for δ13C, 3.4 ± 0.7‰ for δ15N) and omnivores (badger and marten; −19.2 ± 0.6‰ for δ13C, 5.5 ± 0.7‰ for δ15N) from this site.
Discussion
The domestic dogs likely consumed the same varied dietary resources as the human. If it is supposed that the dogs ate marine resources, then the radiocarbon dates need to be corrected for the marine reservoir effect. Hence, the δ13C and δ15N values were used to evaluate this effect on the dogs’ radiocarbon ages, because the δ13C and δ15N values of marine fish and shellfish are generally higher than those of terrestrial mammals (Schoeninger and Deniro, 1984). According to our results, the δ15N values of the dogs and human were 4–5‰ higher than those of the herbivores, but these values did not deviate from the range of the enrichment effect (3–5‰) observed in a terrestrial ecosystem (Bocherens and Drucker, 2003).
In human populations that do not consume C4 plants, δ13C values can also be used as an index for marine contribution to the diet (Richards and Trinkaus, 2009). To investigate the difference between the δ13C values of the dogs and wild mammals, the δ13C values of the dogs were subjected to the Smirnov–Grubbs test, which detects outliers from normal distributions within a population. The results show that the δ13C values of the dogs were not significantly different from those of terrestrial wild mammals. While previous studies (Richards et al., 2005) suggested that marine food consumption raised the δ13C values more than −18‰, the δ13C values of the dogs were less than −18‰. These results indicate that the dogs did not consume a significant amount of marine resources. Therefore, we conclude that the marine reservoir effect on the 14C dates of the dogs is negligible.
To evaluate the difference in the 14C dates produced by the PLD and RLAHA, conventional 14C ages on the same individuals were evaluated by T values following χ2 distribution (Ward and Wilson, 1978). The result from Dog No. 1 showed T = 17.0 (df = 1, P < 0.05) and Dog No. 2 showed T = 7.1 (df = 1, P < 0.05), suggesting that the conventional 14C ages of the two dogs from PLD are statistically different from those of RLAHA. It is possible that these differences depend on collagen extractions, the graphite synthesis and/or data correction between RLAHA and PLD. On the other hand, evaluating the difference in dates between both of the dogs, which were very likely buried at the same time, it was shown that the difference between them was minimal in the PLD results (χ2-test: T = 0.38, df = 1, P > 0.05), but significant in the case of RLAHA (χ2-test: T = 6.7, df = 1, P < 0.05). It is presumed that the two Kamikuroiwa dogs were buried simultaneously by Jomon people, because it was reported that the excavated area of Dog No. 1 was very close to Dog No. 2. Accordingly, there is no strong indication that the RLAHA 14C dates for the samples pretreated by ultrafiltration are more reliable than those produced by PLD. Therefore, we finally calculated the arithmetical mean of the conventional 14C ages between the two institutions: Dog No. 1 dates to 6442 ± 56 BP and Dog No. 2 dates to 6359 ± 51 BP. The calibrated ages (1σ) assigned were 7422–7322 calBP (68.2%) for Dog No. 1; and 7413–7392 calBP (8.7%), 7372–7357 calBP (5.4%) and 7331–7249 calBP (54.1%) for Dog No. 2.
Based on these mean values, the contemporaneity between these two dogs was again subjected to a χ2 test. The T values were less than 3.8 (T = 1.2 < 3.84, df = 1), showing that the burial events of these two dogs could be regarded as simultaneous in terms of radiocarbon dating. In summary, we estimated that the age of the burial event was 6397 ± 38 BP (uncalibrated), and 7414–7390 calBP (15.7%), 7373–7356 calBP (10.5%) and 7332–7273 calBP (42.0%) (1σ).
Esaka et al. (1967) previously suggested that these dog burials originated from Layer 4 along with Oshigatamon potteries of the middle Initial Jomon period. Recently Kobayashi and Onbe (2009) reported that the radiocarbon ages of the dog burials are clearly younger than the middle Initial Jomon period (Figure 3, Table 3). Assigning the ages of dog burials to any potteries after the Oshigatamon type in Layer 4 means that they were similar or slightly older than the Todoroki type.
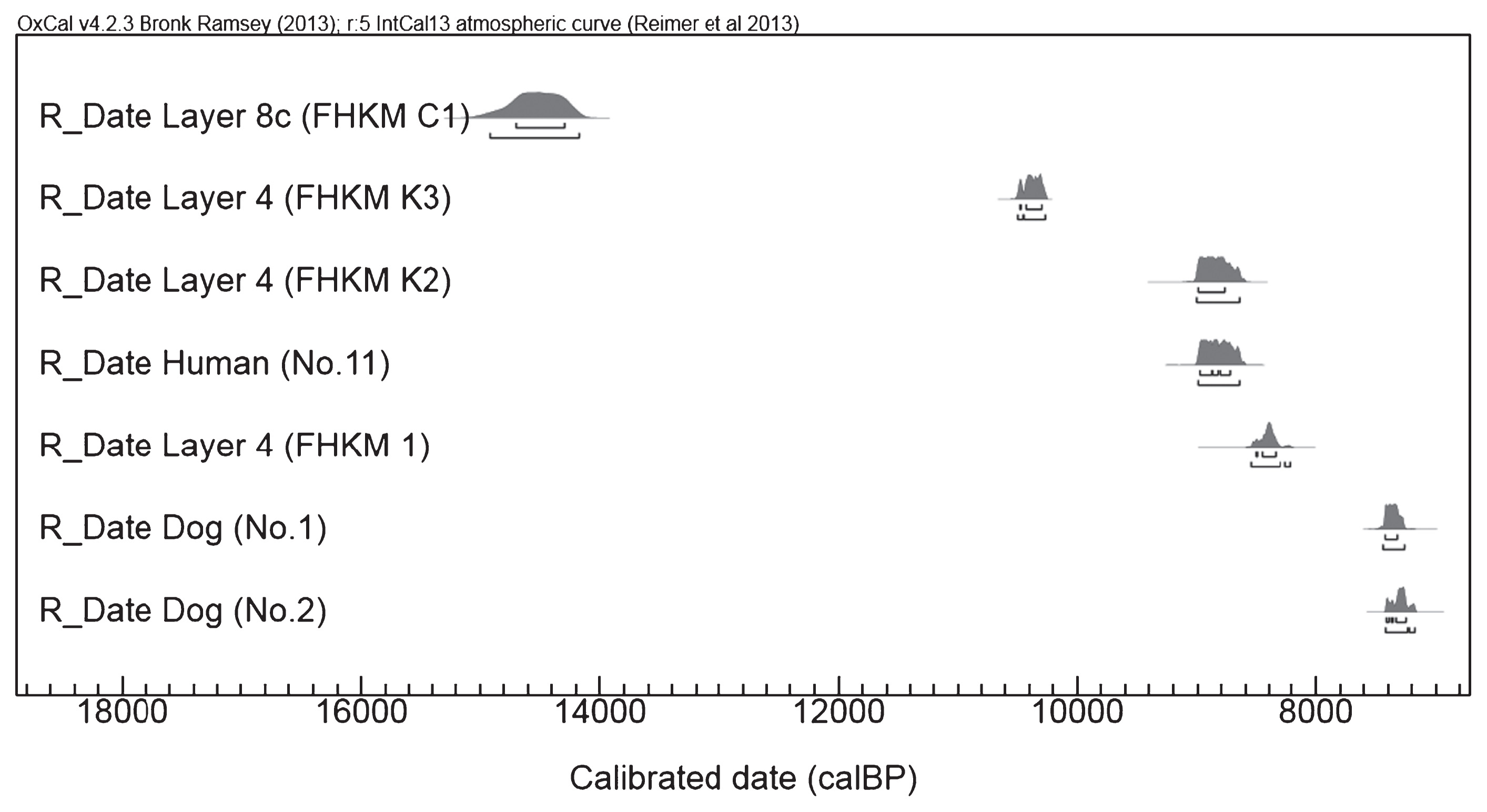
Table 3
Reported radiocarbon ages in previous studies (
Kobayashi and Onbe, 2009)
| Sample ID |
Material |
Location |
Layer |
14C Age (BP) |
Lab-Code |
| FHKM |
C1 |
Charcoal |
A |
8c |
12420 ± 60 |
MTC-04312 |
| FHKM |
K3 |
Freshwater clam |
Extended A |
4 (Lower layer) |
9220 ± 30 |
PLD-8845 |
| FHKM |
K2 |
Thiaridal snail |
Extended A |
4 (Lower layer) |
7980 ± 60 |
MTC-08961 |
| FHKM |
K1 |
Thiaridal snail |
Extended A |
4 |
7600 ± 70 |
MTC-08960 |
| FHKM |
K10 |
Thiaridal snail |
B |
9 |
11640 ± 60 |
MTC-08965 |
| FHKM |
K12 |
Thiaridal snail |
B |
9c |
9680 ± 50 |
MTC-08967 |
| FHKM |
K11 |
Thiaridal snail |
B |
9c |
9150 ± 50 |
MTC-08966 |
| FHKM |
K7 |
Thiaridal snail |
B |
6 |
11670 ± 70 |
MTC-08963 |
| FHKM |
K8 |
Thiaridal snail |
B |
6 |
8840 ± 60 |
MTC-08964 |
| FHKM |
K5 |
Thiaridal snail |
B |
5 |
9690 ± 60 |
MTC-08962 |
| FHKM |
C2 |
Charcoal |
C |
9-VI |
12530 ± 40 |
Beta-201260 |
The Todoroki-type potteries are divided into Todoroki-A of the latest Initial Jomon and Todoroki-B of the earliest Early Jomon. The Todoroki-A type was mainly recovered from layers beneath the K-Ah (Kikai-Akahoya eruption) deposit in southern Kyushu, whose age is greater than 7300 calBP (Machida and Sugiyama, 2002). On the other hand, the Todoroki-B type was excavated above the K-Ah deposit, in layers whose age was estimated at 7000–6700 calBP (Kuwahara, 2008). Given that this chronology based on pottery typology could be applied to the Kamikuroiwa rock shelter, it is possible that the dog burials (7414–7273 calBP) could be assigned to the period of Todoroki A of the latest Initial Jomon.
In another example, a dog burial from the Mizuko shell midden in Saitama Prefecture was found beneath a pit house, and was associated with Kurohama-type pottery with estimated calibrated ages of 7500–7000 calBP (Kobayashi, 2008). This implies that the Jomon people had a similar custom of intentional burial for dogs in both western and eastern Japan in the latest Initial Jomon period. Furthermore, the age of the Natsushima dog, which is considered the oldest dog in Japan, was assigned to the period of Tado-Kashoushiki-type pottery (9400–9200 calBP) of the early Initial Jomon (Kudo, 2012). Thus, the archeological contexts suggest that the fragmentary dog remains from the Natsushima shell midden are older than the complete dog remains from the Kamikurowai rock shelter.
Comparing other cases of contemporaneous dog remains in East Asia, clear evidence of a domesticated dog was found at the Dadiwan Site from the pre-Yanshao culture in northern China. It was reported that the ages of these dogs were 7560–7160 calBP, which are similar to the ages of the Kamikuroiwa dogs. In the Dadiwan site, the size of the dog remains indicated the animal was significantly smaller than a wolf, and stable isotope analysis indicated that these domesticated dogs were fed C4 plants (e.g. cultivated millet) (Barton et al., 2009). In addition, the oldest complete dog skeleton in East Asia was excavated from the Jiahu Site in China, which is dated to 9000–7800 calBP. Therefore, based on the radiocarbon dating and stable isotope analysis, the dog burials from the Kamikuroiwa rock shelter represent some of the oldest complete dogs in East Asia.
Although the age of the dog burials differed from that of the human remains in the Kamikuroiwa rock shelter, it is interesting that the dogs’ diet was quite similar to that of the human. In a previous study of dog nutrition in modern times, the diet of dog is generally similar to that of the humans with whom it lives (Tankersley and Koster, 2009). In archeological studies, it is often reported that the isotopic signatures of dog remains are similar to those of human remains compared with other wild mammals (Noe-Nygaard, 1988; Clutton-Brock and Noe-Nygaard, 1990; Fischer et al., 2007; Yoneda, 2008; Rick et al., 2011; Barton et al., 2009; Koike, 1995; Yoneda and Gakuhari, 2011).
Although the δ13C values of the dogs from the Kamikuroiwa rock shelter do not differ from those of wild herbivores and omnivores in the same site, their δ15N values were clearly higher than those of wild omnivores (Figure 2). This signal probably indicates that the dogs’ dietary signature was affected by eating food left by Jomon peoples. Although the human and the two dogs lived at different times, the similarity of both isotopic signatures might indicate that the human diet in the earliest Jomon was not significantly different from the early Jomon in this area.
Conclusion
In this study, the two dog burials from the Kamikuroiwa rock shelter were assigned to the latest Initial Jomon or initial Early Jomon periods, which is the marginal period between Layer 4 and Layer 3, although they had been previously assigned to Layer 4 of the middle Initial Jomon. On the other hand, the human remains were clearly dated to Layer 4, suggesting that the ages of the dogs differed from the age of the human. The results of the direct carbon dating of the Kamikuroiwa dog burials indicate that they are one of the oldest dog burials in Japan and also one of the oldest clearly buried dog specimens in East Asia.
Acknowledgments
In regard to radiocarbon dating, we received generous support from Dr. T. Omori in The University Museum of The University of Tokyo. We are grateful to Dr. Y. Tanimura and Dr. M. Saito for cross-checking the stable isotope analysis results. We thank Dr. R. W. Schmidt for his help with improving the English grammar. This study was supported by a Grant-in-Aid for JSPS Fellows and Grant-in-Aid for Scientific Research in Japan Society for the Promotion of Science (JSPS).
References
- Ambrose S.H. (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. Journal of Archaeological Science, 17: 431–451.
- Anezaki T., Yoshinaga A., Sato T. and Nishimoto T. (2009) Animal bones. Bulletin of the National Museum of Japanese History, 154: 325–342 (in Japanese).
- Barton L., Newsome S.D., Chen F., Wang H., Guilderson T.P. and Bettinger R. (2009) Agricultural origins and the isotopic identity of domestication in northern China. Proceedings of the National Academy of Sciences, 106: 5523–5528.
- Benecke K. (1987) Studies on early dog remains from Northern Europe. Journal of Archaeological Science, 14: 31–49.
- Bocherens H. and Drucker D. (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. International Journal of Osteoarchaeology, 13: 46–53.
- Clutton-Brock J. and Noe-Nygaard N. (1990) New osteological and C-isotope evidence on mesolithic dogs: companions to hunters and fishers at Star Carr, Seamer Carr and Kongemose. Journal of Archaeological Science, 17: 643–653.
- Davis S.J.M. and Valla F.R. (1978) Evidence for domestication of the dog 12000 years ago in the Natufian of Israel. Nature, 276: 608–610.
- Dayan T. (1994) Early domesticated dogs of the Near East. Journal of Archaeological Science, 21: 633–640.
- Deniro M.J. (1985) Postmortem preservation and alteration of in vivo bone collagen in relation to palaeodietary reconstruction. Nature, 317: 806–809.
- Ding Z.L., Oskarsson M., Ardalan A., Angleby H., Dahlgren L.G., Tepeli C., Kirkness E., Savolainen P. and Zhang Y.P. (2012) Origins of domestic dog in Southern East Asia is supported by analysis of Y-chromosome DNA. Heredity, 108: 507–514.
- Esaka T., Okamoto K., Nishida S. and Kataoka G. (1967) Shikoku chiho no doukustu iseki. In: Nihon koukogaku kyokai doukutsu tyousa tokubetsu iinkai (eds.), Nihon no Doukutsu Iseki. Heibonsha, Tokyo, Chapter 7, pp. 219–250 (in Japanese).
- Fischer A., Olsen J., Richards M., Heinemeier J. and Sveinbjornsdottir A.E. (2007) Coast–inland mobility and diet in the Danish Mesolithic and Neolithic: evidence from stable isotope values of humans and dogs. Journal of Archaeological Science, 34: 2125–2150.
- Kitagawa H., Masuzawa T., Nakamura T. and Matsumoto E. (1993) A batch preparation method for graphite targets with low background for AMS 14C measurements. Radiocarbon, 35: 295–300.
- Kobayashi K. (2008) Jomon doki no nendai (Higashi nihon). In: Kobayashi T. (ed.), Souran Jomon Doki. Amu Promotion, Tokyo, pp. 896–903 (in Japanese).
- Kobayashi K. and Onbe S. (2009) Carbon 14 dating of charcoal and shellfish specimens excavated at Kamikuroiwa. Bulletin of the National Museum of Japanese History, 154: 325–342 (in Japanese).
- Kobayashi K., Niu E., Itoh S., Yamagata H., Lomtatidze A., Jorjoliani I., Nakamura K. and Fujine H. (2007) The compact 14C AMS facility of Paleo Labo Co., Ltd., Japan. Nuclear Instruments and Methods in Physics Research Section B, 259: 31–35
- Koike H. (1995) Mizuko kaiduka shutsudo jinkotunado no tanso chisso antei douitaihi bunseki. In: Fujimi City Board of Education (eds.), Fujimi-shi Bunkazai Houkoku, 46, Fujimi City Board of Education, Fujimi, pp. 208–209 (in Japanese).
- Komiya H. (2003) Jomon ken no kennkyu doukou. Koukogaku Journal, 501: 4–5 (in Japanese).
- Komiya H., Sawada J., Saeki F. and Sato T. (2015) Morphological characteristics of buried dog remains excavated from the Kamikuroiwa Rock Shelter site, Ehime Prefecture, Japan. Anthropological Science, 123: 73–85.
- Kudo Y. (2012) Environment and Cultural History of the Upper Palaeolithic and the Jomon Period. Shinsen, Tokyo (in Japanese).
- Kuwahara M. (2008) Todoroki shiki doki. In: Kobayashi T. (ed.), Souran Jomon Doki. Amu Promotion, Tokyo, pp. 328–335 (in Japanese).
- Larson G., Karlsson E.K., Perri A., Webster M.T., Ho S.Y.W., Peters J., Stahl P.W., Piper P.J., Lingaas F., Fredholm M., Comstockl K.E., Modiano J.F., Schelling C., Agoulnik A.I., Leegwater P.A., Dobney K., Vigne J.D., Vilà C., Andersson L. and Lindblad-Toh K. (2012) Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proceedings of the National Academy of Sciences, 109: 8878–8883.
- Li X., Harbottle G., Zhang J. and Wang C. (2003) The earliest writing? Sign use in the seventh millennium BC at Jiahu, Henan Province, China. Antiquity, 77: 31–44.
- Lindblad-Toh K., Wade C.M., Mikkelsen T.S., Karlsson E.K., Jaffe D.B., Kamal M., Clamp M., Chang J.L., Kulbokas E.J., Zody M.C., Mauceli E., Xie X., Breen M., Wayne R.K., Ostrander E.A., Ponting C.P., Galibert F., Smith D.R., deJong P.J., Kirkness E., Alvarez P., Biagi T., Brockman W., Butler J., Chin C-W., Cook A., Cuff J., Daly M.J., DeCaprio D., Gnerre S., Grabherr M., Kellis M., Kleber M., Bardeleben C., Goodstadt L., Heger A., Hitte C., Kim L., Koepfli K-P., Parker H.G., Pollinger J.P., Searle S.M.J., Sutter N.B., Thomas R., Webber C. and Lander E.S. (2005) Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature, 438: 803–819.
- Longin R. (1971) New method of collagen extraction for radiocarbon dating. Nature, 230: 241–242.
- Macdonald D.W. and Sillero-Zubiri C. (2004) The Biology and Conservation of Wild Canids. Oxford University Press, Oxford.
- Machida H. and Sugiyama S. (2002) The impact of the Kikai- Akahoya explosive eruptions on human societies. In: Grattan J. and Torrence R. (eds.), Natural Disasters and Cultural Change. Routledge, London, pp. 313–346.
- Matsuzaki H., Nakano C., Tsuchiya Y.S., Kato K., Maejima Y., Miyairi Y., Wakasa S. and Aze T. (2007) Multi-nuclides AMS performances at MALT. Nuclear Instruments and Methods in Physics Research B, 259: 36–40.
- Minagawa M., Winter D.A. and Kaplan I.R. (1984) Comparison of Kjeldahl and combustion methods for measurement of nitrogen isotope ratios in organic matter. Analytical Chemistry, 56: 1859–1861.
- Nikolai D.O., Crockford S.J., Kuzmin Y.V., Higham T.F.G., Hodgins G.W.L. and van der Plicht J. (2011) A 33000-year-old incipient dog from the Altai Mountains of Siberia: Evidence of the earliest domestication disrupted by the Last Glacial Maximum. PLoS ONE, 6: e22821 doi: 10.1371/journal.pone.0022821.
- Noe-Nygaard N. (1988) δ13C-values of dog bones reveal the nature of changes in man’s food resources at the mesolithic-neolithic transition, Denmark. Chemical Geology: Isotope Geoscience section, 73: 87–96.
- Onbe S. (2009) Carbon 14 dating of cord-patterned pottery from the Kamikuroiwa site. Bulletin of the National Museum of Japanese History, 154: 325–342 (in Japanese).
- Payne S. (1983) The animal bones from the 1974 excavations at Douara cave. The University Museum, The University of Tokyo Bulletin, 21: 1–10.
- Pionnier-Capitan M., Bemilli C., Bodu P., Celerier G., Ferrie J., Fosse P., Garcia M. and Vigne J. (2011) New evidence for Upper Palaeolithic small domestic dogs in South-Western Europe. Journal of Archaeological Science, 38: 2123–2140.
- Protsch R. and Berger R. (1973) Earliest radiocarbon dates for domesticated animals. Science, 179: 235–239.
- Ramsey B.C. (2009) Bayesian analysis of radiocarbon dates. Radiocarbon, 51: 337–360
- Ramsey B.C., Higham T. and Leach P. (2004) Towards high-precision AMS: progress and limitations. Radiocarbon, 46: 17–24.
- Reimer P.J., Bard E., Bayliss A., Beck J.W., Blackwell P.G., Bronk Ramsey C., Grootes P.M., Guilderson T.P., Haflidason H., Hajdas I., HattŽ C., Heaton T.J., Hoffmann D.L., Hogg A.G., Hughen K.A., Kaiser K.F., Kromer B., Manning S.W., Niu M., Reimer R.W., Richards D.A., Scott E.M., Southon J.R., Staff R.A., Turney C.S.M. and van der Plicht J. (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50000 years cal BP. Radiocarbon, 55: 1869–1887.
- Richards M.P. and Trinkaus E. (2009) Isotopic evidence for the diets of European Neanderthals and early modern humans. Proceedings of the National Academy of Sciences, 106: 16034–16039.
- Richards M.P., Jacobi R., Cook J., Pettitt P.B. and Stringer C.B. (2005) Isotope evidence for the intensive use of marine foods by Late Upper Palaeolithic humans. Journal of Human Evolution, 49: 390–394.
- Rick T.C., Culleton B.J., Smith C.B., Johnson J.R. and Kennett D.J. (2011) Stable isotope analysis of dog, fox and human diets at a Late Holocene Chumash village (CA-SRI-2) on Santa Rosa Island, California. Journal of Archaeological Science, 38: 1385–1393.
- Savolainen P., Zhang Y., Luo J., Lundeberg J. and Leitner T. (2002) Genetic evidence for an East Asian origin of domestic dogs. Science, 298: 1610–1613.
- Schoeninger M. and Deniro M.J. (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochemica et Cosmochimica Acta, 48: 625–639.
- Shigehara N. and Hongo H. (2000) Ancient remains of Jomon dogs from Neolithic sites in Japan. Dogs Through Time: An Archaeological Perspective. Archaeopress, Oxford.
- Sugawara H. (2003) Maisou sareta Jomonken to sanranshita kenkotsu. Kouko Gaku Journal, 501: 16–19 (in Japanese).
- Sugihara S. and Serizawa T. (1957) Kanagawa-ken Natsushima niokeru Jomon bunka syotou no kaiduka. Rinsen, Kyoto (in Japanese).
- Tankersley K.B. and Koster J.M. (2009) Sources of stable isotope variation in archaeological dog remains. North American Archaeologist, 30: 361–375.
- van Klinken G.J. (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. Journal of Archaeological Science, 26: 687–695.
- Vila C., Maldonado J.E. and Wayne R.K. (1999) Phylogenetic relationships, evolution and genetic diversity of the domestic dog. Journal of Heredity, 90: 71–77.
- von Holdt B.M., Pollinger J.P., Lohmueller K.E., Han E., Parker H.G., Quignon P., Degenhardt J.D., Boyko A.R., Earl D.A., Auton A., Reynolds A., Bryc K., Brisbin A., Knowles J.C., Mosher D.S., Spady T.C., Elkahloun A., Geffen E., Pilot M., Jedrzejewski W., Greco C., Randi E., Bannasch D., Wilton A., Shearman J., Musiani M., Cargill M., Jones P.G., Qian Z., Huang W., Ding Z.-L., Zhang Y.-p., Bustamante C.D., Ostrander E.A., Novembre J. and Wayne R.K. (2010) Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication. Nature, 464: 898–902.
- Wang G., Zhai W., Yang H., Fan R., Cao X., Zhong L., Wang L., Liu F., Wu H., Cheng L., Poyarkov A.D., Poyarkov N.A., Jr, Tang S., Zhao W., Gao Y., Lv X., Irwin D.M., Savolainen P., Wu C. and Zhang Y. (2013) The genomics of selection in dogs and the parallel evolution between dogs and humans. Nature Communications, 4: 1860 doi: 10.1038/ncomms2814.
- Ward G.K. and Wilson S.R. (1978) Procedures for comparing and combining radiocarbon age determinations: a critique. Archaeometry, 23: 19–31.
- Yoneda M. (2008) Jomon jinkotsu oyobi doubutsukotsu no douitaibunseki. In: Toshi saisei kikou Chiba ken kyouiku shinkou zaidan (ed.), Chiba-ken kyouiku shinkou zaidan tyousa houkokusho Chiba-shi Ariyoshi-minami kaiduka, Toshi saisei kikou Chiba ken kyouiku shinkou zaidan, Chiba, pp. 392–397 (in Japanese).
- Yoneda M. and Gakuhari T. (2011) Kushiro cho Tenneru I iseki kara shutsudo shita Jomon jidai jinkotsu doubutsukotsu no douitaibunseki. In: Kushiro City Board of Education (ed.), Kushiro Cho Tenneru I Iseki 2, Chuou Kouban, Hokkaido, pp. 404–408 (in Japanese).
- Yoneda M., Tanaka A., Shibata Y., Morita M., Uzawa K., Hirota M. and Uchida M. (2002) Radiocarbon marine reservoir effect in human remains from the Kitakogane site, Hokkaido, Japan. Journal of Archaeological Science, 29: 529–536.

