Original Articles
Health and disease of hunter-gatherer groups from the eastern Pampa–Patagonia transition (Argentina) during the Late Holocene
2016 Volume 124 Issue 1 Pages 29-44

Details
2016 Volume 124 Issue 1 Pages 29-44
In the eastern Pampa–Patagonia transition, hunter-gatherer groups experienced a series of changes in social organization (e.g. subsistence, mobility, and demography) during the Late Holocene (c. 3000–250 years BP), mainly in the last c. 1000 years. In this context, some biological aspects, including health, might have also been modified. The aim of this paper is to present a review and synthesis of a recent pathological analysis of hunter-gatherer skeletons from the lower basin of the Colorado River (Argentina) during the Late Holocene. The sample includes 94 individuals from six archaeological sites. Bone and dental pathologies (e.g. osteoarthrosis, systemic stress, bone infections, trauma, caries, periodontal disease, antemortem tooth loss, and dental calculus) were considered. The results show low frequencies in most of the bone and oral pathologies. Trauma, infectious, and systemic stress processes did not show large variation among samples from the Late Holocene. However, increasing prevalence of dento-alveolar lesion and osteoarthrosis were observed in samples from the final moments of this period (c. 1000–250 years BP). These results are discussed taking into account the archaeological model proposed for the eastern Pampa–Patagonia transition, which suggests social transformations and reorganizations during the last 1000 years.
Several researches have indicated that changes in social organization and subsistence patterns produced effects on lifestyle and health of human populations (Cohen and Armelagos, 1984; Larsen, 1997; Bartelink, 2006; García Guraieb, 2010; Gómez Otero and Novellino, 2011; Pinhasi and Stock, 2011; Scabuzzo, 2011). Archaeological data for the Pampas and northeastern Patagonia regions (Argentina) indicate major changes and reorganization of cultural systems in hunter-gatherer groups during the Final Late Holocene (c. 1000–250 14C years BP). These changes included diverse socioeconomic and demographic phenomena such as population growth, population expansions, inter-ethnic contacts, social interaction networks at different spatial scales (local, regional, and extra-regional), territoriality, spatial circumscription, intensification and diversification of the subsistence activity, demographic packing, and possible regionalization (Barrientos and Pérez, 2004; Berón, 2007; Martínez, 2008–2009; Politis, 2008; Prates, 2008; Favier Dubois et al., 2009; Martínez et al., 2014).
Hunter-gatherers from the lower basin of the Colorado River (Figure 1) experienced these changes. The probabilistic distribution of radiocarbon dates suggests an increase in the archaeological signal during the last 1000 years, probably linked to a demographic growth (Martínez et al., 2013). Also, more stable settlements, inter-site variability, and changes in mobility were observed (Martínez, 2008–2009). Regarding funerary practices, multiple secondary burials and formal disposal areas were adopted during this period (Martínez et al., 2012). Pottery was more systematically used and produced than in the earlier period (c. 3000–1000 14C years BP). Zooarchaeological studies indicate a process of diversification and intensification in the exploitation of fauna (Alcaráz, 2015; Stoessel, 2014), while the analysis of lithic technology indicates more complex and extensive ‘chaînes opératoires’ and increased frequency of formal artifacts (Armentano, 2012).
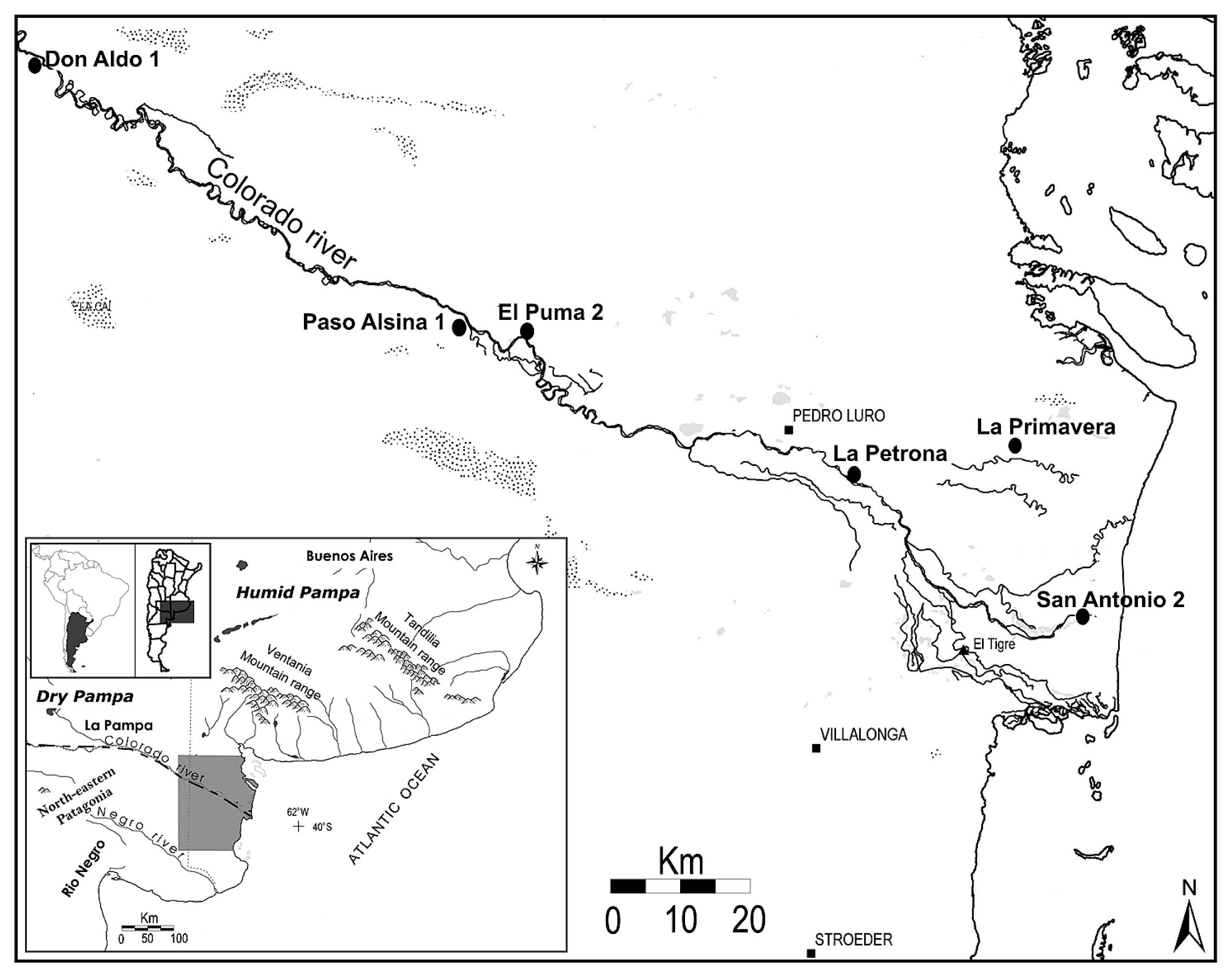
Location of the six sites analyzed in the lower basin of the Colorado River.
Changes in different spheres of social organization, including subsistence, mobility, and demography, suggest that some biological aspects, including health, might have also been modified (Flensborg, 2012). In order to study possible changes in the health of human populations as a result of the social transformations that occurred during the last 1000 years BP, this paper presents a review and synthesis of the recent pathological results obtained from the analyses of hunter-gatherer skeletal remains from the lower basin of the Colorado River during the Late Holocene (c. 3000–250 years BP).
The lower basin of the Colorado River (province of Buenos Aires, Argentina) is part of the ‘Arid Diagonal’ (Abraham de Vázquez et al., 2000), which is characterized by a steppe of arid and warm climate. During the Late Holocene, the climate was continually arid and semi-arid with alternating processes of morphogenesis and pedogenesis (Abraham de Vázquez et al., 2000; Schäbitz, 2003; Martínez and Martínez, 2011). The paleoclimatic information, although scarce, indicates no significant climatic changes (Stoessel et al., 2008; Martínez and Martínez, 2011; Fernández, 2012). At present, the area is covered by a shrub steppe, an open vegetal formation composed by short xeric trees which are mixed with hard and scarce herbaceous grasses. The dominant vegetation belongs to the ‘Distrito del Caldén’ in the phytogeographic province of the ‘Espinal,’ although vegetal communities from the phytogeographic province of the ‘Monte’ are also recorded (Cabrera, 1994; Paez et al., 2001). The area is dominated by C3 plant species (e.g. Geoffroea decorticans, Prosopis sp., and Condalia microphylla), although C4 species are also present in a subordinated proportion (Villamil and Scoffield, 2003; Martínez et al., 2009; Sánchez et al., 2014). From a zoogeographical perspective, the area is located in the Patagonia subregion, Patagonia district (Cabrera and Yepes, 1960). Ichthyogeographic studies from the Colorado and Negro Rivers indicate that there is an overlap in the distribution of fish species from the Austral and Brazilian subregions (Almirón et al., 1997). In sum, according to the association of different vegetation, animal, and fish communities, the study area is considered an ecotone, characterized by a high productivity and biodiversity of resources (Paez et al., 2001; Villamil and Scoffield, 2003; Sánchez et al., 2014).
The skeletal series recovered from six Late Holocene archaeological sites of the lower basin of the Colorado River were analyzed from a pathological perspective by the author (Figure 1, Table 1; Flensborg, 2012). In general, the earliest human burials (c. 3000–1000 years BP) are characterized by a low number of individuals buried in primary modality, which are located in base camps and isolated contexts (e.g. La Primavera, San Antonio 2, and El Puma 2 sites; Table 1). In contrast, the later burials (c. 1000–250 years BP) are characterized by a greater number of individuals, represented mostly by multiple secondary burials. These are located at base camps which were reoccupied through time and at formal disposal areas or cemeteries (e.g. Paso Alsina 1, Don Aldo 1, and La Petrona sites; Table 1) (Martínez et al., 2012).
| Period | Site | MNI | MNE | MNT | MNA |
|---|---|---|---|---|---|
| Initial Late Holocene | PRI | 6 | 115 | 58 | 83 |
| EP2 | 1 | 38 | 16 | 5 | |
| SA2 | 1 | 4 | 13 | 23 | |
| Total ILH | — | 8 | 157 | 87 | 111 |
| Final Late Holocene | DA1 | 1 | 112 | 27 | 23 |
| LP | 8 | 354 | 79 | 101 | |
| PA1 | 77 | 3526 | 780 | 1025 | |
| Total FLH | — | 86 | 3992 | 886 | 1149 |
| Total sample | — | 94 | 4149 | 973 | 1260 |
Abbreviations: MNI, minimal number of individual; MNE, minimal number of elements; MNT, minimal number of teeth; MNA, minimal number of alveoli; PRI, La Primavera; EP2, El Puma 2; SA2, San Antonio 2; DA1, Don Aldo 1; LP, La Petrona; PA1, Paso Alsina 1.
The skeletal sample analyzed consists of 94 individuals including 4149 skeletal elements, 973 teeth and 1260 alveoli (Table 1). However, as a consequence of differential preservation of skeletal parts (see below), the size of the study sample varies according to the bone or oral pathology under evaluation. For comparison, the skeletal remains were grouped into two major time lapses: Initial Late Holocene (ILH; 3000–1000 years BP) and Final Late Holocene (FLH; 1000–250 years BP).
Sex determination was only made for adults (individuals older than 15 years of age) using macroscopic observations of different qualitative and/or morphological bone features. The more diagnostic bone elements were analyzed (e.g. skull, os coxae, and sacrum) using conventional methods (Buikstra and Ubelaker, 1994). To estimate the probable skeletal age at death of adults, several methodological criteria that take into account changes in the morphology of the skull, pelvis, and sacrum were considered (Buikstra and Ubelaker, 1994). For non-adults, both macroscopic and osteometrical methods were used (Scheuer and Black, 2000; see Flensborg et al., 2013a). The following age categories were taken into account: fetal (before birth), perinate (36–44 weeks), infant (1month–3 years), child (3–12 years), adolescent (13–20 years), young adult (20–35 years), middle adult (35–50 years), and old adult (older than 50 years) (Buikstra and Ubelaker, 1994).
The analysis and regional comparison is based upon the most usual markers for assessing the health of the hunter-gatherer populations from Pampas and Patagonia (e.g. Luna, 2008; García Guraieb, 2010; Menéndez, 2010; Scabuzzo, 2011; Gordón, 2013), which generally have reasonably high rates of occurrence in skeletal series. These include oral pathologies (e.g. caries, dental calculus, antemortem tooth loss, and periodontal disease), osteoarthrosis, systemic stress, bone infections, and trauma. All human remains were analyzed macroscopically and using binocular magnifier glass (40×). The frequency of each lesion was calculated by tooth, alveolus, bone element, and individual. Observed differences between the frequencies of lesions by age and sex were tested using chi-squared tests (differences at or below P = 0.05 were considered significant), with the software package PAST (version 2.08; Hammer et al., 2001).
Caries, periapical lesion, antemortem tooth loss, dental calculus, and dental wear in 68 individuals (ILH = 8; FLH = 60), 974 teeth (ILH = 87; FLH = 887) and 1260 alveoli (ILH = 111; FLH = 1149) were studied (Flensborg, 2011a, 2012, 2013). Maxilla and mandible with a completeness of over 75% were analyzed. Dental wear was scored for all permanent teeth corresponding to adolescent and adult individuals, using the scoring system proposed by Smith (1984) and Scott (1979) for anterior and posterior dentition, respectively. For the caries analysis, the criteria of Buikstra and Ubelaker (1994) were used. The locations of the lesions were standardized as occlusal, mesial, distal, lingual, and buccal (Hillson, 2001). In addition, according to the depth of carious lesions, they were classified on the basis of affected tissue into enamel, dentin, and pulp cavity caries (Lanfranco and Eggers, 2010). For periapical lesion, the methodology proposed by Ogden (2008) was used. The diameter and the texture of the alveoli and the appearance of the edge features were observed. Antemortem tooth loss was recorded by the presence of alveolar obliteration (Lukacs, 1989). Dental calculus was recorded according to the criteria proposed by Brothwell (1981) as a low amount (affects less than one-third of the tooth), a moderate amount (affects between one-and two-thirds), and a high quantity (up to two-thirds).
Osteoarthrosic lesions were recorded macroscopically in 2983 bone elements corresponding to 54 adult individuals (Flensborg, 2012). Only anatomical units with joints in good preservation were included in the analysis (ILH = 3 individuals; 110 bone elements; FLH = 51 individuals; 2873 bone elements). The guidelines proposed by Waldron (2009) were considered. The author indicates the presence of osteoarthritis when eburnation or at least two of the following bone alterations are observed: marginal osteophytosis, presence of new bone on the joint surface, alteration in joint contour, and pitting on the joint surface. The ordinal scale proposed by Jurmain (1990) was used to assess the severity of injuries.
The cribra orbitalia (CO), porotic hyperostosis (PH), and linear enamel hypoplasias (LEH) were analyzed (Flensborg, 2012). The PH was analyzed in 53 completed skulls (ILH = 6; FLH = 47 individuals), and CO in 35 skulls with both orbits preserved (ILH = 6; FLH = 29 individuals). The LEH was recorded in 518 permanent teeth (ILH = 54; FLH = 464), corresponding to 51 adults and adolescent individuals (ILH = 4; FLH = 47). The presence/absence of porotic hyperostosis and cribra orbitalia was macroscopically examined in all skulls. The severity of both conditions was categorized as stages 1–4: 1, scattered fine foramina; 2, large and small isolated foramina; 3, foramina linked into a trabecular structure; 4, outgrowths in trabecular form from the outer table surface (Stuart-Macadam, 1985). The major stage observed in each individual was considered as the final value. Finally the degree of healing was classified as active, remodeling, or reshaping (Stuart-Macadam, 1985). The linear enamel hypoplasias were identified with a binocular magnifier glass (40×) and with oblique lighting. Only symmetrical enamel defects in opposing teeth were considered in the analysis (Barrientos, 1999). To estimate the approximate age of enamel defect formation, the histological method was considered (Reid and Dean, 2006). The distance between the cemento-enamel junction and linear enamel hypoplasias was measured with a digital caliper (accurate to 0.01 mm).
The presence/absence of infectious processes was analyzed in a sample of 94 individuals and 4149 bones (ILH = 8 individuals and 157 bone elements; FLH = 86 individuals and 3992 bone elements; Flensborg, 2012). A detailed description of the lesion according to their location, general characteristics, and distribution in the skeleton (e.g. bilateral/unilateral, symmetrical/asymmetrical) was considered (Aufderheide and Rodríguez-Martín, 1998; Ortner, 2003). For subperiosteal reactions, non-invasive methods such as radiography and computed tomography were applied (see Flensborg, 2012; Flensborg et al., 2013b).
The traumatic events in 4149 bones from 67 adults and adolescent individuals (ILH = 8 individuals and 157 bone elements; FLH = 59 individuals and 3992 bone elements) were analyzed (Flensborg, 2011b, 2012). In the postcranial sector, fractures, traumas, and lithic projectile injures were considered. Along the cranial sector, lesions were classified into: (a) fractures (linear and star-shape); (b) perforations with embedded projectile points and/or perforations without projectiles but with morphologies suggesting presence of the latter, (c) depressions (circular and ovoid); and (d) bone bumps (circular and ovoid) (Lovell, 1997; Aufderheide and Rodríguez-Martín, 1998; Gordón, 2013, etc.). Trauma was discriminated as perimortem or antemortem based on the presence or absence of healing and fracture morphology (Lovell, 1997). Postmortem fractures were also analyzed in order to avoid confusing changes generated by taphonomic processes (Lovell, 1997). In addition, forensic techniques were applied to analyze the patterns of fractures and blunt trauma (Berryman and Jones Haun, 1996).
The results indicate that in both periods, the degree of tooth wear was similar. In this sense, increasing tooth wear was observed with age (Figure 2) and no sex differences were recorded (Figure 3). Therefore, the differences that can be recorded between oral pathologies by periods cannot be explained by the action of tooth wear (Flensborg, 2012, 2013).
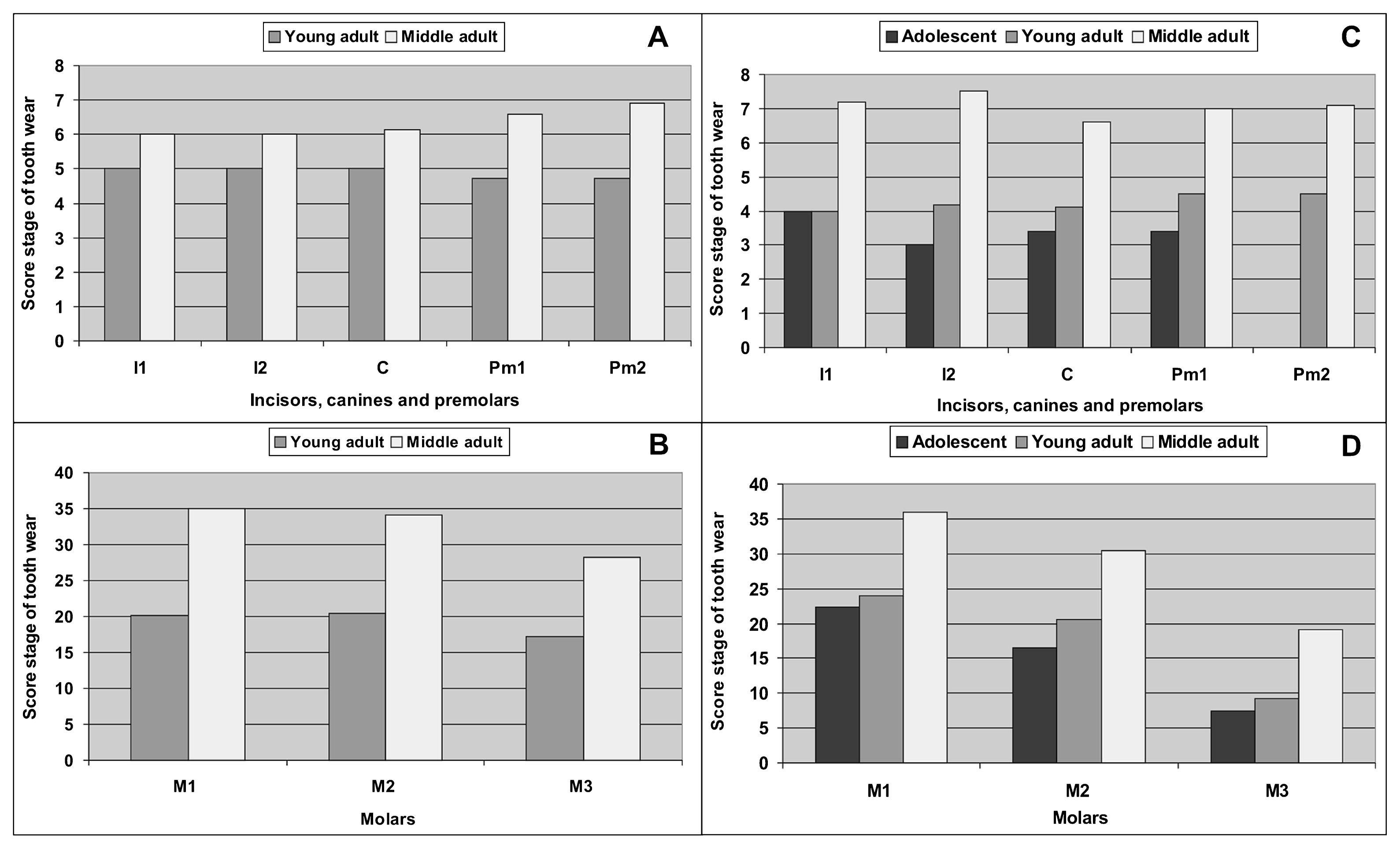
Score stage of tooth wear by category of age during the Initial Late Holocene (A, B) and during the Final Late Holocene (C, D).
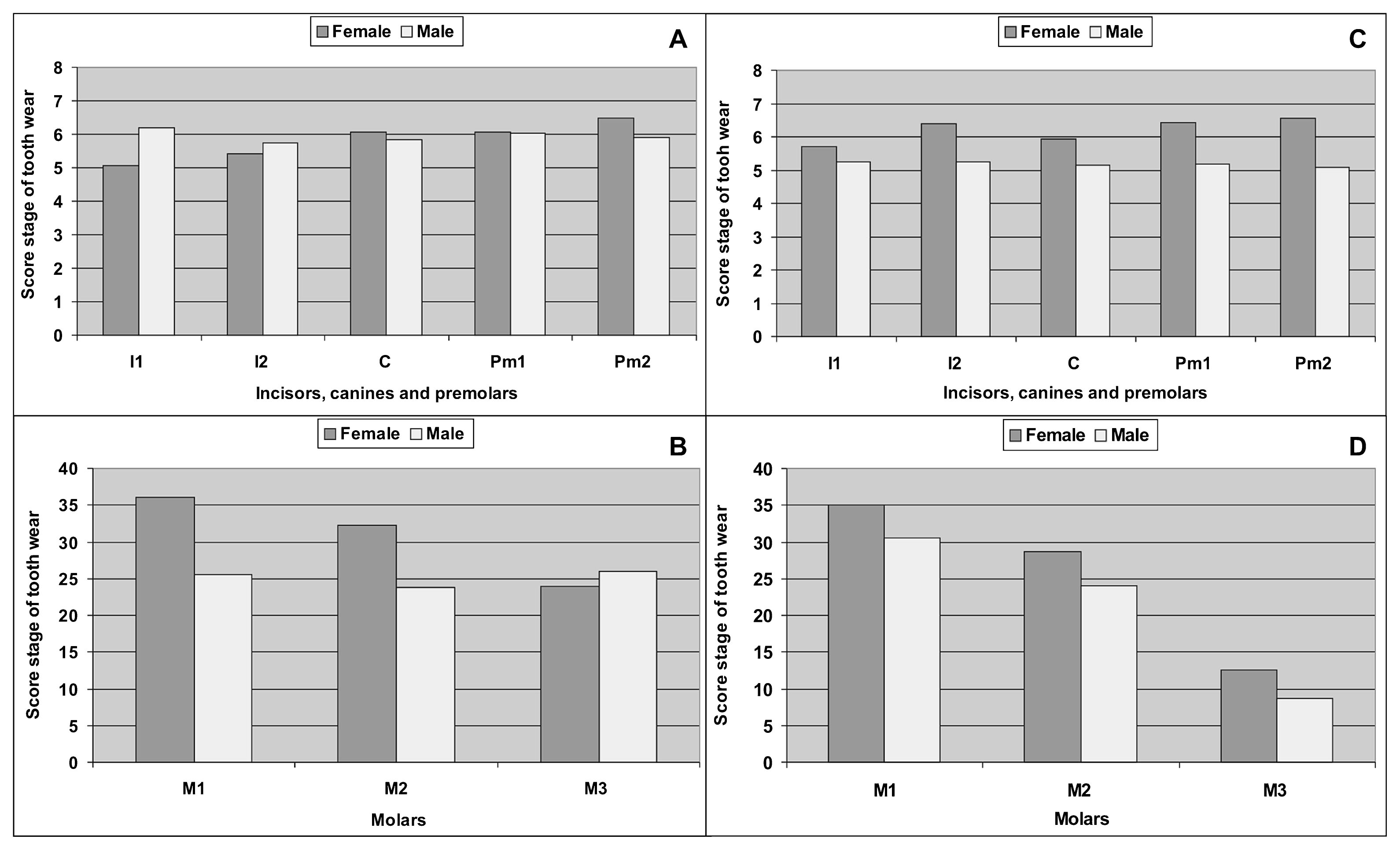
Score stage of tooth wear by sex during the Initial Late Holocene (A, B), and during the Final Late Holocene (C, D).
Caries were not observed in any of the individuals during the ILH (Table 2). In contrast, low prevalence was recorded during the FLH (5.2% of teeth; Table 2). The differences between periods are statistically significant (Table 2, Table 3). Caries were recorded in 36.6% of the individuals (Table 3), and in similar frequencies for both sexes (36% male and 39.1% female). All age categories had at least one individual with caries, although young adults (51.6%) out-number adolescents (33.3%) and middle adults (20%). These differences are only significant when the frequencies among adult categories were compared (Table 4). A higher frequency of caries in molars (9.7%; 37/375), in relation to the anterior teeth (1.7%; 9/512) was observed. Caries were located mainly in the occlusal surface (69.6%), followed by the labial (13%), lingual (8.7%), mesial, and distal surface (4.3% each). The majority of lesions were observed in the dentin (60.8%), followed by the pulp cavity (21.7%) and only a few in the enamel (17.4%).
| Variable | ILH | FLH | χ2 | P | ||
|---|---|---|---|---|---|---|
| A/O | % | A/O | % | |||
| Caries | 0/87 | 0 | 46/887 | 5.2 | 4.735 | 0.029* |
| Periapical lesion | 5/111 | 4.5 | 28/1149 | 2.4 | 1.696 | 0.192 |
| ATL | 0/111 | 0 | 83/1149 | 7.2 | 8.583 | 0.003* |
| Dental calculus | 5/87 | 5.7 | 231/887 | 26 | 17.776 | <0.001* |
Abbreviations: A/O, affected/observed; ATL, antemortem tooth loss.
| Variable | ILH | FLH | χ2 | P | ||
|---|---|---|---|---|---|---|
| A/O | % | A/O | % | |||
| Caries | 0/8 | 0 | 22/60 | 36.6 | 4.336 | 0.037* |
| Periapical lesion | 3/8 | 37.5 | 14/60 | 23.3 | 0.806 | 0.369 |
| ATL | 0/8 | 0 | 15/60 | 25 | 2.566 | 0.109 |
| Dental calculus | 2/8 | 25 | 35/60 | 58.3 | 3.161 | 0.07 |
Abbreviations: A/O, affected/observed; ATL, antemortem tooth loss.
| ILH | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Caries | Periapical lesion | ATL | Dental calculus | ||||||
| χ2 | P | χ2 | P | χ2 | P | χ2 | P | ||
| Sex | M/F | — | — | 0.15 | 0.220 | — | — | 0.37 | 0.541 |
| Age | YA/MA | — | — | 2.88 | 0.089 | — | — | 0.17 | 0.673 |
| FLH | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| χ2 | P | χ2 | P | χ2 | P | χ2 | P | ||
| Sex | M/F | 0.05 | 0.822 | 0.25 | 0.61 | 1.27 | 0.258 | 0.42 | 0.514 |
| Age | Adol/YA | 0.36 | 0.545 | 0.43 | 0.507 | 0.31 | 0.572 | 2.19 | 0.138 |
| Adol/MA | 0.28 | 0.594 | 1.59 | 0.207 | 2.52 | 0.112 | 0.12 | 0.724 | |
| YA/MA | 5.90 | 0.015* | 4.14 | 0.041* | 10.3 | 0.001* | 5.28 | 0.021* | |
Abbreviations: M, male; F, female; YA, young adult; MA, middle adult; Adol, adolescent.
The low prevalence of caries in the FLH (5.2%) is within the expected ranges for the hunter-gatherer way of life as proposed by Turner (1979, between 0.0 and 5.3%) and Larsen (1987, between 0.0 and 7.8%). The differences observed with respect to the ILH may be due to an assortment of factors, which include the under-representation of the sample. However, some interpretations can be made. Caries are linked to the consumption of a diet rich in carbohydrates, sugars, and sticky foods; therefore, the proportion of caries provide information about carbohydrate and protein intake ratios (Hillson, 2001; Bonfiglioli et al., 2003; Eshed et al., 2006; Bernal et al., 2007; Hujoel, 2009). The frequencies reported for the Late Holocene suggest an increase and recurrent consumption of cariogenic foods and/or changes in food preparation (e.g. use of pottery) during the FLH (Flensborg, 2013). Besides, the location of the caries, mainly affecting the occlusal surface and in most cases the dentin and the pulp cavity, could indicate that enamel demineralization was a process that occurred in the long term, and continued to operate beyond different abrasives that damaged the surface of the crown (e.g. tooth wear). Furthermore, the presence of caries in different age categories indicates that they were recurrent throughout the individual’s lives. The highest frequency of lesions in young adults could be linked to the lower tooth wear recorded (Flensborg, 2012).
Periapical lesions were reported in low frequency at the alveolar level for both ILH (4.5%) and FLH (2.4%), with no statistically significant differences (Table 2, Table 3; Flensborg, 2013). During the ILH, periapical lesions were recorded in 37% of individuals (Table 3) and affected mostly female and middle adults (50%), although no statistical differences were observed when compared with other categories (Table 4). The most frequently affected alveoli were observed in the first molar (12.5%). During the FLH, the periapical lesions were recorded in 23.3% of the individuals (Table 3). The distribution of lesions was similar between sexes (24% male and 30.4% female) and affected mostly middle adults (36%) compared to young adults (12.9%). Here, the difference is statistically significant (Table 4). The highest frequency of periapical lesion occurred in the first molars (9.2%) and to a lesser extent in the anterior teeth.
Numerous factors can cause periapical lesions, such as severe wear, a crown fracture, trauma, and/or the formation of caries (Lukacs, 1992; Dias and Tayles, 1997). All of these cases can provide direct access of microorganisms to the dental pulp and cause infection. In both periods, the higher prevalence of periapical lesions in middle adult individuals may be related to the progressive process of tooth wear. During the FLH, caries may have played an important role in the development of this pathology (Flensborg, 2012, 2013).
Antemortem tooth loss was recorded with low frequency (7.2%), only in individuals from the FLH (Flensborg, 2013). The difference between periods is statistically significant at the alveolar level (Table 2), but not among individuals (Table 3). The lesions were recorded in 25% of the individuals (24% male and 39.1% female) and mainly in middle adults (48%), compared with young adults (9.6%). This difference is statistically significant (Table 4). The highest frequency of antemortem tooth loss was recorded in the first molar (17%), second molar (12%), and second premolar (10.1%). This oral pathology can occur through various events or processes, such as infection of the pulp cavity, tooth fracture, intensive attrition, or cultural avulsion (Lukacs, 1992; Littleton and Frohlich, 1993). It is possible that the tooth wear and periapical lesions were the main causes because middle adults suffered higher rates of antemortem tooth loss. However, this pathology was only recorded in individuals from the FLH, a similar situation to that of caries. In addition, the most affected alveoli (e.g. first molars) are the teeth that have the highest frequency of caries (Flensborg, 2011a, 2013). Accordingly, it is possible that caries has been a major cause of tooth loss during the life of individuals.
Dental calculus was recorded at a lower frequency during the ILH (5.7%) and moderate prevalence during the FLH (26%). These differences are statistically significant (Table 2). Dental calculus was recorded in 25% of the individuals during the ILH. These affected both sexes (50% male and 25% female), and both young (33.3%) and middle adults (20%). The plaque was deposited in low amounts, especially in molars. During the FLH, dental calculus was registered in 58.3% of the individuals, and in a similar frequency for both sexes (56% male and 65.2% female). Dental calculus was recorded more frequently in young adults (74.2%), followed by middle adults (44%) and adolescents (33.3%, Table 4). The molars and premolars were the most affected and to a lesser extent, the incisors and canines. A low amount of dental calculus was found in 87.4% of the teeth and 12.6% in the moderate category.
The presence of dental calculus in both periods suggests poor oral hygiene and indicates that individuals had an alkaline oral environment (Flensborg, 2013). This is facilitated by the precipitation of minerals from the oral fluids (e.g. saliva and gingival fluid; Lukacs, 1989; Hillson, 2000), which accumulate on tooth surfaces as plaque and then undergo a process of mineralization after the intervention of microorganisms such as bacteria (Mandel, 1987; Hillson, 2000). While there are many factors involved in the generation of dental calculus (plaque pH, the concentration of calcium phosphate in the oral fluid, etc.; Nancollas and Johnsson, 1994), diet has been a part of major interest in research (see references in Lieverse, 1999). An integrative approach suggests that the mineralization process is related to the amount of plaque and the factors that increase their accumulation, such as poor oral hygiene and consumption of carbohydrates (Hillson, 2000; Bonfiglioli et al., 2003). The results indicate a significant increase of dental calculus during the FLH. It is possible that an increase in carbohydrate intake may have had a role in its development.
Higher prevalences of caries, antemortem tooth loss, and dental calculus during the FLH indicate changes in diet associated with a higher intake of carbohydrates. In the study area there are several species of plants with sweet and sticky fruits, and a high caloric content of carbohydrates, particularly sugars and starch, including the ‘chañar’ (Geoffroea decorticans), ‘algarrobo’ (Prosopis sp.), and ‘piquillín’ (Condalia microphylla). Analyses of Prosopis alba and Prosopis flexuosa show that they contain high percentages of carbohydrates (more than 50%) with a variable proportion of fibers and proteins (Roig, 1993). This same situation is observed in G. decorticans (Orrabalis et al., 2014). According to the ethnohistorical and archaeological evidence from northern Patagonia, these sweet fruits were commonly used for direct consumption and for the preparation of flour and fermented beverages (Prates, 2009; Caparelli and Prates, 2010, 2015). While plant resources were exploited by human groups throughout the Late Holocene, is possible that most dento-alveolar lesions during the FLH were the result of a change in food processing. Although pottery technology was present 1500 years BP, increased production for culinary use occurred during the FLH (Martínez et al., 2011). This technology could provide other substances such as liquids and fermentable beverages that perhaps were not consumed regularly in the ILH (Flensborg, 2013). Furthermore, the increase in the frequency of grinding stones in the archaeological record, as well as more standardized and formal tool designs, suggests that plant resources played an important role in the hunter-gatherer diet during the FLH (Martínez, 2008–2009; Armentano, 2012).
From a comparative perspective, the frequencies shown in the study area are similar to those reported for neighboring regions (e.g. lower valley of the Negro and Chubut Rivers, north-central coast of Patagonia, Dry Pampa sub-region; Menéndez, 2010; Gómez Otero and Novellino, 2011; Luna and Aranda, 2014). In addition, an increase in the intake of plant resources was proposed for the FLH (c. 1000–250 years BP) of the lower valley of the Negro River and southeast of the Pampas region (Figure 1; L’Heureux, 2000; Menéndez, 2010). This suggests that changes in the intake of plant resources during the Late Holocene may be observed not only in the lower basin of the Colorado River but also at a wider spatial scale.
OsteoarthrosisOsteoarthrosic lesions were recorded in similar frequencies in individuals from both periods (ILH = 66.6%; FLH = 60%), affecting both sexes and different ages in the adult category (Flensborg, 2012). However, considering the percentage of anatomical units affected by osteoarthrosis and the total number of anatomical units analyzed in each period, a higher frequency was observed in the FLH (30.3%; 870/2873) than in the ILH (4.5%; 5/110). The difference is statistically significant (χ2 = 21.558; P < 0.001). Besides, individuals present lesions that affected particular joints (e.g. hip, ankle and jaw) during the ILH. In contrast, a great variety of lesions was recorded in several joints of the skeleton (e.g. hand, elbow, knee, and spine) in moderate to high percentages, during the FLH (Figure 4). Also, differences were recorded in the type of alterations. In all cases, individuals present two of the four parameters required for the definition of osteoarthrosis (e.g. marginal osteophytosis, changes in joint contour, and small pitting on the joint surface) during the ILH sample. Conversely, cases of eburnation, lysis in the vertebral bodies, and development of porosity, and a greater length of marginal osteophytosis were recorded during the FLH. Regarding the degree of development of lesions, during the ILH all cases developed in grade 1. In contrast, during the FLH, although most injuries occur in grade 1 (92%), grade 2 (7%) and grade 3 (1%) were also recorded.
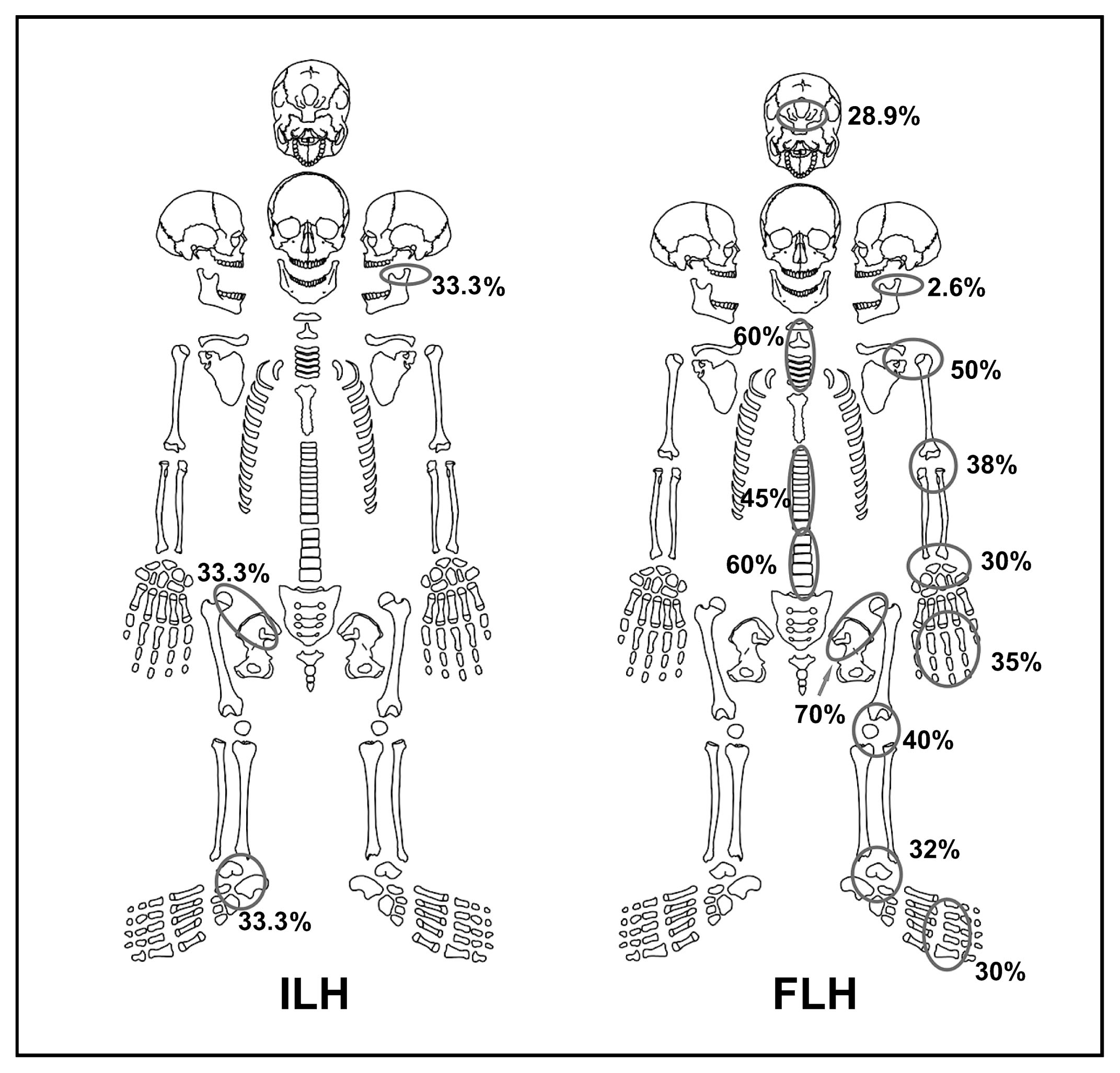
Frequency and distribution of osteoarthrosic lesions in each joint in both periods.
The differences between the two periods can be caused by problems in the sampling bias, because of the low representation of individuals during the ILH. Although interpretations should be cautiously examined, further development of degenerative joint disease may have also occurred throughout the FLH. There are many factors that can generate osteoarthrosis, such as deficiencies in metabolism, nutrition, trauma, body mass, genetic influences, age and sex (Jurmain, 1990; Solano, 2002; Weiss, 2006; Weiss and Jurmain, 2007; Waldron, 2009, 2012). The possibility of metabolic disorders, genetic influences, systemic processes (age) and trauma are not considered as the main factors in this case study (see discussion in Flensborg, 2012). The skeletal series analyzed correspond to hunter-gatherer groups whose lifestyle involves a demanding level of muscle-skeletal effort, physical activity and mechanical stress. These could be the main causes for the development of joint lesions (e.g. locomotion, weight bearing and various domestic activities; Jurmain, 1990; Weiss, 2006; Rojas-Sepúlveda et al., 2008).
From a larger spatial scale, researches in the Dry Pampa (Luna, 2008) and Humid Pampa sub-regions (Scabuzzo, 2011) reveal similar osteoarthrosic injury, affecting the same sections of the skeleton in adult individuals of both sexes. These lesions were interpreted as a result of recurrent ambulatory activity and different domestic tasks (e.g. use of the bow and arrow, moving heavy objects; Luna, 2008; Scabuzzo, 2011). Researches in northeastern Patagonia are scarce and do not allow a comparison. According to the information generated at the regional level, it is suggested that physical activities played an important role in the development of osteoarthrosic lesions in the hunter-gatherer groups. In this sense, changes in social organization in the lower basin of the Colorado River during the FLH probably impacted on behaviors and activities of human groups (Martínez, 2008–2009). The adoption of a logistical mobility strategy, with more stable settlements and longer occupations, may have caused changes in the behavior of human groups. These include the systematic adoption of certain body postures (e.g. squatting) for the development of different domestic activities and a change in the ambulatory pattern. In this regard, it is proposed that a higher intensity of physical activities (e.g. stone knapping, grinding, and movement of objects) generated stress on the musculoskeletal system. This caused further development of degenerative joint disease during the FLH (Flensborg, 2012).
Systemic stressPorotic hyperostosis (PH) and cribra orbitalia (CO) were only recorded in individuals from the FLH (Table 5), although no statistical differences were observed between both periods for PH (χ2 = 3.1; P = 0.077) and CO (χ2 = 0.93; P = 0.333). Porotic hyperostosis was recorded in 34% and cribra orbitalia was calculated at 13.8%. Both sexes and adult categories were recorded in similar frequencies (Table 5). Porotic hyperostosis was located bilaterally in the posterior portion of both parietal bones, the region adjacent to the lambdoidal suture, and the center of the occipital squamous. Orbital and vault lesions were characterized in stage 1 of severity, and the degree of healing was remodeled in all cases.
| Sex | Age at death | |||||||
|---|---|---|---|---|---|---|---|---|
| A/O | % | M | F | I | YA | MA | I | |
| PH | 16/47 | 34 | 64.7 | 60 | 30 | 60.8 | 56.2 | 0 |
| CO | 4/29 | 13.8 | 25 | 22.2 | 0 | 28.5 | 20 | 0 |
Abbreviations: A/O, affected/observed; M, male; F, female; YA, young adult; MA, middle adult; I, indeterminate.
Linear enamel hypoplasias were recorded in both periods. Enamel defects were observed during the ILH in 5.5% of the teeth (all in canines) and in 50% of the individuals (Table 6), all male and young adults. During the FLH, linear enamel hypoplasias were recorded in 5.2% of the teeth (22 canines and 2 incisors), affecting 27.6% of the individuals and in similar proportions for both sexes (23.1% male and 21.4% female) and categories of young adults (25%) and middle adults (33.3%). The differences observed between the two periods are not statistically significant (Table 6). With respect to the age estimation of linear enamel hypoplasias formation, the results indicate that the defects occurred in an age range between 2 and 5 years, although there is a more limited range for the ILH (2 and 3.5 years, Figure 5).
| ILH | % | FLH | % | χ2 | P | |
|---|---|---|---|---|---|---|
| Individual | 2/4 | 50 | 13/47 | 27.6 | 0.886 | 0.346 |
| Tooth | 3/54 | 5.5 | 24/464 | 5.2 | 0.014 | 0.904 |
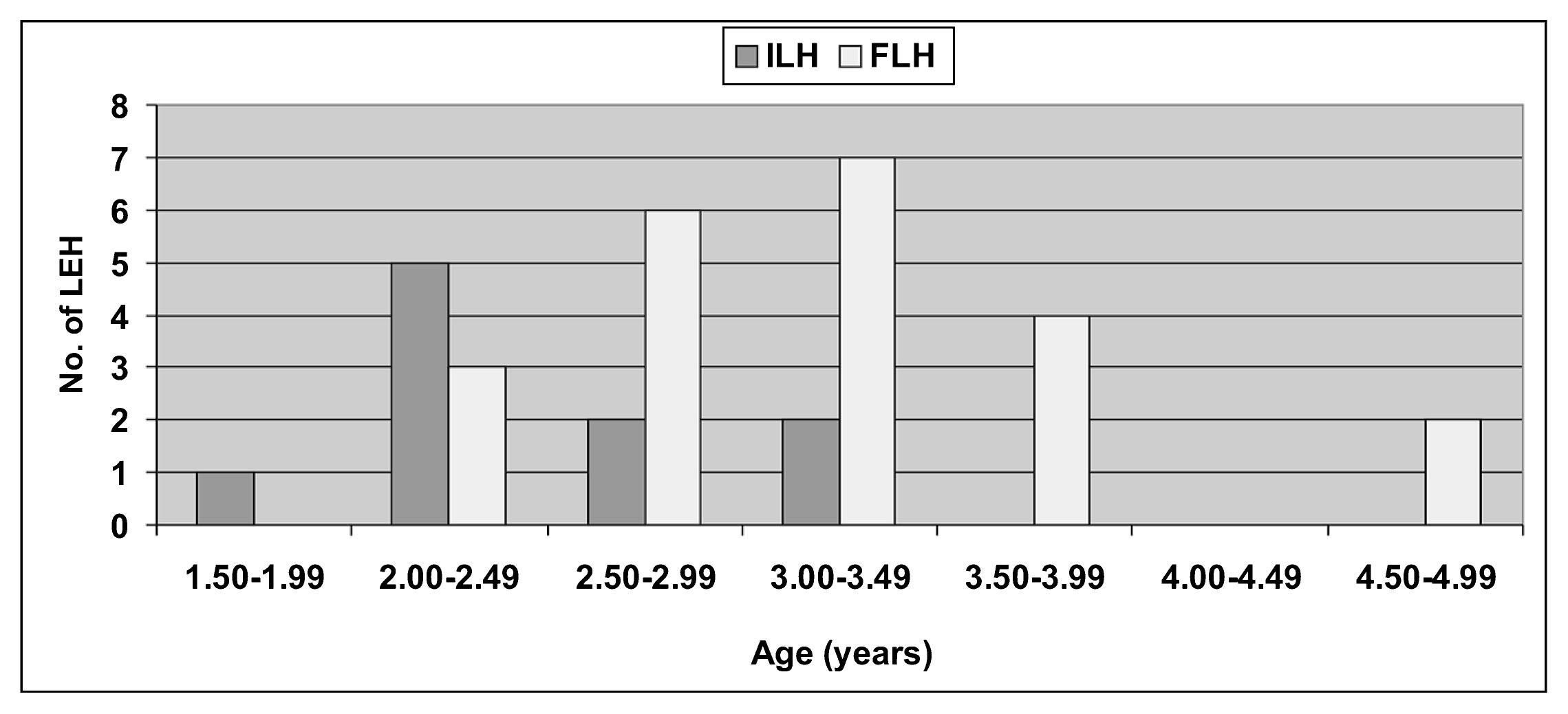
Age (in years) for linear enamel hypoplasias formation in both periods for 6-month intervals.
Systemic stress lesions were recorded for both periods, but mainly for the FLH. Although the sample has significant biases that may influence the interpretation of the data, some aspects can be discussed. There are numerous factors that could cause systemic stress events and iron deficiency. They consist of metabolic disorders, vitamin deficiencies, low dietary intake, megaloblastic and hemolytic anemia, infectious processes, gastrointestinal hemorrhage, among others (Lallo et al., 1977; Stuart-Macadam, 1985; Mensforth, 1991; Walker et al., 2009; Vercellotti et al., 2010). In this case study, porotic hyperostosis and cribra orbitalia are not consistent with genetic anemia. In America, this type of anemia (e.g. thalassemia, sickle cell anemia) has not been reported in prehistoric populations (Aufderheide and Rodríguez-Martín, 1998; Ortner, 2003). Therefore, the presence of PH and CO was possibly due to exposure of acquired anemia.
It is unlikely that a dietary deficiency has occurred in the lower basin of the Colorado River, because the hunter-gatherer groups often consumed iron-rich meat resources. Zooarchaeological studies indicate recurrent exploitation and consumption of terrestrial species (e.g. guanaco) during the Late Holocene. Moreover, a greater variety of resources (e.g. birds, mollusks, freshwater and marine fish) were exploited and consumed during the FLH (Alcaráz, 2015; Stoessel, 2012). In addition, a process of intensification in the exploitation of guanaco was observed for this period (e.g. meat, fat, and marrow; Stoessel, 2014). Stable isotope studies on human skeletal remains indicate a consumption of protein provided by meat of terrestrial herbivores (e.g. artiodactyls) complemented by fish, during the Late Holocene (Martínez et al., 2009). In this sense, the results support the idea of iron-rich intake of foods, cobalatina (vitamin B12) and folic acid (vitamin B9). These nutrients are essential for growth and development of individuals (Larsen, 1997; Walker et al., 2009; García Guraieb, 2010). Nevertheless, since porotic hyperostosis and cribra orbitalia are probably developed during childhood (Stuart-Macadam, 1985; Ortner, 2003), some causes may be related to processes of systemic stress during this stage of development. The process of prolonged breastfeeding in infants may generate a poor source of iron, vitamin B12, nutrients, and a low proportion of lymphocytes and immunoglobulins, because the milk tends to decrease its quality throughout time (see Katzenberg et al., 1996; Ryan, 1997; Walker et al., 2009). In addition, children suffer health risks of megaloblastic anemia when mothers begin to deplete vitamin B12 during breastfeeding (Katzenberg et al., 1996; Sellen, 2001; Casella et al., 2005). Dental enamel hypoplasias indicate that amelogenic disruptions occurred during childhood in both periods. The average estimated age of enamel defects was similar between the two periods (2.5–4.0 years), indicating that systemic stress processes were possibly similar to each other during the Late Holocene. This coincides with the expected age of weaning infants in most populations (see Katzenberg et al., 1996). Although other causes cannot be discounted (e.g. parasitic infections), it is hypothesized that prolonged breastfeeding, the weaning process and replacement of breastfeeding with solid foods would have generated physiological perturbations during growth and development of individuals (see discussion in Katzenberg et al., 1996).
The presence of the linear enamel hypoplasias and the remodeled condition of the porotic hyperostosis and cribra orbitalia in adults suggests that individuals were able to survive the different episodes of stress during childhood. No difference in the frequency of lesions was observed between males and females, which does not support the idea of sex as a risk factor predictive of anemia in human populations from the study area. However, it is also necessary to consider other causes that may explain the presence of PH and CO until adulthood. The acquired anemia often results from infections with gastrointestinal parasites and zoonotic infections (Blom et al., 2005; Walker et al., 2009; Suby, 2014). The medical literature demonstrates that iron deficiency anemia can be caused or exacerbated by one or more intestinal parasites (Crompton, 2000; Horton, 2003). Transmission between individuals is related to poor health and a low personal hygiene condition. The most common modes of transmission are oral or fecal routes. This may be related to the type of settlement, water pollution and the spatial proximity to the where the waste is disposed. However, this feature is commonly related to sedentary settlements in which human groups occupy the same place for a long time (Lallo et al., 1977; Larsen, 1997; Bathurst, 2005). So far, the cause still needs to be evaluated. Also, the intake of aquatic resources such as fish and shellfish is a major route of contamination (Blom et al., 2005; Bartelink, 2006; Walker et al., 2009). These resources have worms and parasites (e.g. nematodes), which can cause anemia (Aufderheide and Rodríguez-Martín, 1998; Walker et al., 2009). In the eastern Pampa–Patagonia transition, it was proposed that the consumption of freshwater and marine aquatic resources (e.g. fish and shellfish; Martínez et al., 2009; Stoessel, 2012), can be considered as a possible explanation for some cases of parasitic infections and systemic stress (Flensborg, 2012). With regard to other causes, none of the individuals presented bone lesions related to scurvy or rickets. Bone evidence of tuberculosis and treponematosis was not observed (see below).
In sum, despite the problems of biases that exist in the skeletal series analyzed, the results in this study indicate no differences in systemic stress between the two periods. This suggests that the proposed changes in social organization, such as a longer duration in base camps, higher population density, changes in subsistence linked to processes of intensification, and diversification in the exploitation of resources during the FLH, would not have affect the health of individuals. The low frequencies observed in the variables analyzed in hunter-gatherer populations are possibly linked to an adequate intake of animal protein, high levels of assimilation of iron in the organism, and scarce situations socioenvironmental stress. This situation has also been proposed for neighboring regions, from the analysis of CO and PH. Low prevalence’s were reported in the Dry Pampa sub-region (PH: 0.002%, CO: 5.9%; Luna, 2008), the southeastern Pampean region (absence of CO and PH; Barrientos, 1997), the northwest of southern Patagonia (PH: 4.3%, CO: 26.1%; García Guraieb, 2010) and the central-northern Patagonia coast (CO: 14%; Gómez Otero and Novellino, 2011). Conversely, the linear enamel hypoplasias were recorded in higher prevalence than in this work, such as in the southeastern Pampean region (27.7%; Barrientos, 1999) and northwestern South Patagonia (85.7%; García Guraieb, 2010). These results were interpreted as moderate to high levels of metabolic disorders of acute nature during the early postnatal period of growth and development, occasioned by systemic causes such as the synergistic action of malnutrition and disease states (e.g. infectious processes; Barrientos, 1997). In addition, the formation of linear enamel hypoplasias could be linked to critical periods of childhood, mainly weaning age and the biological risks associated with this process (García Guraieb, 2010). This scenario contrasts with the frequencies obtained in this work, which can be interpreted in part as different processes of adaptation to changing socioenvironmental contexts through time.
Infectious processesBone lesions associated with infectious processes were not recorded in the ILH sample, while a low frequency of lesions for elements (0.1%; 4/3992) and individuals (1.1%; 1/86) was registered during the FLH. These differences were not statistically significant between both periods (χ2 = 0.094; P = 0.759). In this particular case, four anatomical units (tibia, femur, fibula, and skull) of a middle adult female individual showed bilateral and asymmetrical periosteal bone reactions. Woven and lamellar bone formation within the marrow cavity, a thickening of cortical bone with a slightly lytic surface and a small pitted area associated with cloacae were observed through radiographic and CT images (see description in Flensborg et al., 2013b: figures 3 and 4). These lesions were diagnosed as secondary osteomyelitis, and as a result of a contiguous focus of infection (Flensborg et al., 2013b).
In the study area there were no differences in the expression of infectious lesions throughout the Late Holocene. However, it cannot be discounted that other individuals may have suffered from diseases that could not be diagnosed in the skeletal record due to an immune response by the host to the infection or to the early death of the individual (Wood et al., 1992). Furthermore, post-depositional processes could have led to differential preservation of anatomical units, thus biasing the study of bone lesions (Waldron, 1994). Although they could be biases in the sample, the low frequency of infectious processes during the FLH may indicate that organizational changes proposed for this period did not created conditions favorable to possible exposure of infectious diseases. Osteomyelitis was probably caused by the introduction of an infectious agent through an abscess located in the maxilla of the individual (see details in the differential diagnosis: Flensborg et al., 2013b).
From a comparative perspective at an extra-areal level, significant differences were observed. In the southeastern Pampean region, moderate frequencies of infectious injury during the Late Holocene (28%; Barrientos, 1997) were reported, including two adult individuals with hematogenous osteomyelitis. In northwestern South Patagonia, a prevalence of periosteal reactions was reported in 32.6% during the Late Holocene, although the skeletal series post-1000 years BP presented the highest frequency (c. 37%). These lesions were related to traumatic stress and systemic processes such as infections, possibly tuberculosis (García Guraieb, 2010). The observed differences in prevalence between regions can respond to different processes of adaptation of the individuals to changes in the socioenvironmental contexts in the last c. 1000 years BP. However, the frequency of infectious processes recorded in this work is strikingly low, given that they are usually recorded among hunter-gatherer groups (Powell, 1988). Therefore, it is proposed that the results and interpretations reached in this study should be interpreted with caution and explored in future research.
TraumaThe results indicate low frequencies of traumatic lesions in both periods. Traumas were recorded in 25% (2/8) of the individuals and 1.9% of the anatomical units (3/157) during the ILH, while traumas were identified in 37.3% of individuals (22/59) and 1.3% of the bone elements (52/3992) during the FLH. Differences between periods are not statistically significant among individuals (χ2 = 0.462; P = 0.496) or anatomical units (χ2 = 0.427; P = 0.513). However, a greater variety of lesions during the FLH were observed (Table 7). In postcranial bones, arrowhead wounds were observed (0.05%; 2/3992) in few individuals (3.4%; 2/59). In an adult of indeterminate sex, a projectile point was recorded in the acromion process of the scapula. The trajectory of the projectile was in a postero-anterior direction. In an individual aged between 16 and 19 years, and of indeterminate sex, a projectile point was recovered inside the dorsal vertebrae body. Here, the projectile was in the antero-posterior direction. In both cases, the bone tissue is not remodeled (Table 7). In addition, complete and incomplete fractures in adult individuals of indeterminate sex (20.3%; 12/59) were observed in different bones (0.4%; 17/3992) (Table 7). Also, trauma (0.5%; 19/3992), especially in the tibia and phalanges of the hand and foot in adult individuals of indeterminate sex, were recorded (11.9%; 7/59). Ankylosis, new bone formation, increased thickness of the cortical tissue, and periosteal reactions were observed (Table 7). Less diversity in the types of lesions was observed during the ILH. An incomplete oblique fracture remodeled in the distal diaphysis of a right ulna and new bone formation, and periosteal reactions linked with a trauma in proximal diaphysis of a right tibia were recorded only in a male individual aged between 20 and 25 years of age.
| Trauma | Bone element | A/O | % | Location | Type | Remodeled |
|---|---|---|---|---|---|---|
| Arrowheads | scapula | 1/96 | 1 | acromion | perforation with projectil embedded | no |
| dorsal vertebrae | 1/274 | 0.3 | body | perforation with projectil embedded | no | |
| Fractures | clavicle | 1/58 | 1.7 | proximal diaphysis | complete, transverse | yes |
| ulna | 4/97 | 4.4 | distal diaphysis | complete, transverse | yes | |
| distal diaphysis | complete, oblique | yes | ||||
| distal diaphysis | incomplete, oblique | yes | ||||
| distal diaphysis | incomplete, oblique | yes | ||||
| radio | 1/83 | 1.2 | distal diaphysis | incomplete, oblique | yes | |
| metacarpus | 2/75 | 2.6 | diaphysis | complete, oblique | yes | |
| diaphysis | incomplete, transverse | yes | ||||
| femur | 1/116 | 0.8 | midshaft and distal diaphysis | incomplete, comminuted | yes | |
| fibula | 1/97 | 1 | lateral malleolus | complete, transverse | yes | |
| metatarsus | 2/248 | 0.8 | proximal diaphysis | incomplete, oblique | yes | |
| distal diaphysis | incomplete, oblique | yes | ||||
| Trauma | tibia | 3/113 | 2.6 | diaphysis | depression | yes |
| bone bumps | yes | |||||
| bone bumps | yes | |||||
| phalange hand | 11/273 | 4.9 | distal diaphysis and diaphysis | ankylosis, new bone formation, periosteal reactions | yes | |
| phalange foot | 5/163 | 3 |
A great diversity of lesions in the skull was observed for the FLH (Table 8). In this period, 29.8% (17/57) of the individuals showed skull trauma, unlike the ILH sample which had 12.5% (1/8); although no statistical differences (χ2 = 1.051; P = 0.305) were recorded. The lesions predominated in male individuals, but once again, no statistically significant differences were observed between sexes (Table 9). According to the categories of age, trauma occurred at similar frequencies among young and middle adults (Table 9). Taking into account the distribution of lesions among individuals (Figure 6), females have lower percentages and more variety of lesions (mainly depressions); while males have the highest percentage of linear fractures (Figure 6). Linear fractures have variable dimensions (between 1 and 20 mm in length) and were recorded in different portions of the skull, although the occipital bone was the most affected, mainly by perimortem fractures (Table 10). All depressions were remodeled, have different dimensions (diameter between 3 and 18 mm), and mainly localized parietal bones (Table 10). Bumps were recorded on parietal (diameter c. 5 mm) and occipital (diameter c. 20 mm) bones. Finally, a perforation without projectile was recorded in a left parietal (9.80 × 6.74 mm), with no evidence of remodeling.
| Variable | ILH | FLH | ||
|---|---|---|---|---|
| n | % | n | % | |
| Perforation without projectile | 0 | 0 | 1 | 2.4 |
| Bone bumps | 0 | 0 | 2 | 4.8 |
| Depression: circular | 0 | 0 | 7 | 16.7 |
| Depression: ovoid | 1 | 100 | 15 | 35.7 |
| Linear fracture | 0 | 0 | 17 | 40.5 |
| Total | 1 | — | 42 | — |
| n | N | % | P | |
|---|---|---|---|---|
| Male | 11 | 24 | 45.8 | 0.192 |
| Female | 6 | 22 | 27.3 | |
| Indeterminate | 0 | 11 | 0 | |
| Younger adult | 9 | 31 | 29 | 0.732 |
| Middle adult | 8 | 24 | 33.3 | |
| Indeterminate | 0 | 2 | 0 |
Abbreviations: n, affected individuals; N, total individuals.
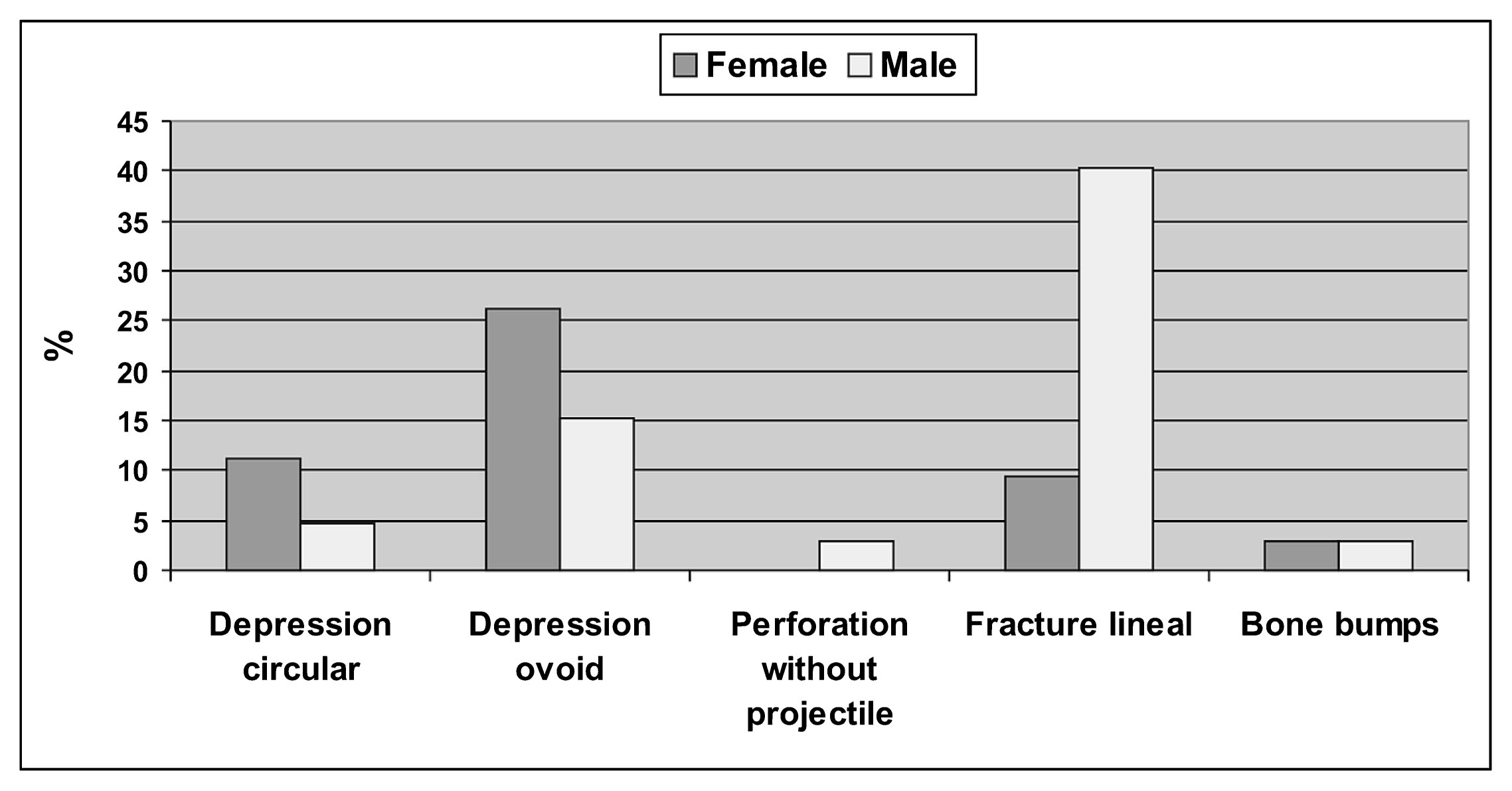
Frequency of types of skull trauma by sex during the Final Late Holocene.
| Trauma | Bone | n | % | % antemortem | % perimortem |
|---|---|---|---|---|---|
| Linear fracture | occipital | 8 | 19.1 | 25 | 75 |
| frontal | 3 | 7.1 | 100 | 0 | |
| nasal | 1 | 2.4 | 100 | 0 | |
| parietal | 2 | 4.7 | 100 | 0 | |
| Depression: ovoid | parietal | 15 | 35.7 | 100 | 0 |
| frontal | 2 | 4.7 | 100 | 0 | |
| Depression: circular | parietal | 8 | 19.1 | 100 | 0 |
| Perforation without projectile | parietal | 1 | 2.4 | 0 | 100 |
| Bone bumps | occipital | 1 | 2.4 | 100 | 0 |
| parietal | 1 | 2.4 | 100 | 0 |
The observed differences in the skeletal series for both periods could influence interpretation of the data. However, the similar traumatic injuries were recorded at low frequencies for both periods. The analysis of trauma has been used to understand social behaviors and lifestyles of human groups, linked to physical activity, violence, and conflict (Larsen, 1997; Jurmain, 2001; Torres-Rouff and Costa Junqueira, 2006; Gordón, 2013). The results obtained here indicate fractures in the distal diaphysis of the ulna (Table 7) for both periods. These fractures are often called ‘Parry’ type (Judd, 2008) and are related to acts of intentional violence, individual conflicts, and wars (Lovell, 1997; Ortner, 2003; Judd, 2008). However, cultural contexts are too complex to discriminate the cause of the fracture (e.g. accidents). Some methodological studies have been proposed for diagnosis of ‘Parry’ type fracture using a set of criteria (e.g. Judd, 2008). In this case criteria such as location, disruption of the ligamentous insertions, and absence of radial involvement were recorded. But other features are not observed, such as a transverse fracture line, since most of the fractures are oblique (Table 7). Given that there are isolated signs of violence in both periods (all in the FLH, see below), it is possible that fractures observed in the ulna correspond to accidents (e.g. falls) and/or chronic stress (Rogers, 1992; Jones, 2006). The rest of the fractures and trauma recorded in the clavicles, femur, fibula, tibia, and bones of the hand and foot, may be due to direct trauma for accidents. For example, clavicular fractures can occur by direct trauma or falls on the shoulder (Ortner, 2003). The location of the lesions (near the distal ends) in the phalanges of the hands and feet may be due to stress fractures (Bennike, 2008; Lovell, 2008; see discussion in Flensborg, 2012).
The clearest evidences of interpersonal violence are the arrowheads embedded in bones (Knüsel, 2005; Smith et al., 2007). Considering that the lesions are not remodeled, it is possible that this could have been the cause of death in two individuals. Except for these two cases, the rest of the traumatic conditions in the postcranial section are remodeled, indicating the survival of individuals of various traumatic events (Flensborg, 2012).
The skulls presented a wide variety of lesions, especially for the FLH (Table 8). Lesions classified as depressions were produced during the life of the individuals. The remodeled condition indicates that they were not lethal. The location of the lesions, mostly in the parietal bone, could be the result of accidental falls or confrontations between two or more individuals (see examples of face to face confrontations in Gat, 1999). The diversity of forms and the small size of the lesions indicate that weapons or specific objects were not used on the skulls (Flensborg, 2011b; see Walker, 1989). Linear fractures were recorded in several individuals. Most were antemortem, but in other cases they were perimortem. These lesions could be caused by the impact of different sharp and thin objects (see below).
No differences were observed in the frequency of traumatic injuries between sexes. It is possible that both sexes suffered similar traumas through life, both accidents and violence. However, male individuals have the highest frequency of linear fractures (Figure 6), which could be more exposed to the impact of objects with sharp edges (e.g. knives, scrapers, arrowheads of small sizes and different shapes; Armentano, 2012). Some of these artifacts were recorded in the bones (e.g. arrowheads) and could have caused some lesions to the skull (e.g. linear fractures, perforations without projectiles). In the case of arrowheads, bows and arrows were the weapon system used by the hunter-gatherer groups in the eastern Pampa–Patagonia transition during the FLH. The characteristics of the lesions do not suggest the use of metal weapons (Barrientos and Gordón, 2004; Gordón, 2013).
Traumatic lesions, such as arrowheads inside the bones, linear fractures on the skull, and perforations without projectiles, could be related to acts of interpersonal violence. The rest of the traumas are considered unreliable to differentiate between accidents and violence (Flensborg, 2012). In the case of violent events, fights could occur through confrontations in everyday life and, to a lesser extent, between neighboring groups. Numerous studies indicate that hunter-gatherer societies were not peaceful (see Larsen, 1997), and violence was documented both in domestic contexts (e.g. between individuals of both sexes, fights between men, ritualized fighting) and between different groups (e.g. ambushes, raids, see examples in Larsen, 1997; Gat, 1999; Lee and Daly, 1999). Flensborg (2011b, 2012) suggested that considering the high frequency of remodeled lesions, perhaps a habit of the hunter-gatherer groups was resolving disputes through duels, in which opponents attacked but did not kill each other (Walker, 1989; Gordón, 2013; see several examples Gat, 1999). The possibility of war as organized aggression between autonomous political units (Thorpe, 2003) is discarded. The archaeological context is not characterized by a pattern of defensive constructions, use of specific weapons, mutilation, scalping, or mass killings which could have affected a group. In many hunter-gatherer societies, ambushes and raids have been documented (see examples in Gat, 1999), which killed or caused damage without differentiating between age and sex. The results in the eastern Pampa–Patagonia transition are not consistent with these types of attacks.
Major changes and reorganization of cultural systems towards the FLH (c. 1000–250 14C years BP), which included population growth, population expansions, demographic packing, territoriality, and spatial circumscription (Martínez, 2008–2009), may have increased violence and conflict among different human groups. However, the results do not indicate an increase in interpersonal violence in the hunter-gatherer groups of the study area during c. 1000–250 years BP (before the period of European–native contact; Flensborg, 2012). This is consistent with research conducted in northeastern Patagonia (Barrientos and Gordón, 2004; Gordón, 2013). In contrast, in the Dry Pampa sub-region, further evidence of indicators of interpersonal violence was recorded between c. 400 and 300 years BP, due to the presence of 54 arrowheads associated and embedded in different bones (Berón, 2012). This conflict scenario was part of organizational changes of hunter-gatherers during 1050–300 years BP, which included a reduction in mobility due to environmental degradation, competition for the access to resources, and control of exchange circuits (Berón, 2007). This context contrasts with the results reported here, suggesting that different regional population dynamics occurred in the same period.
The results of this study, which were obtained in conjunction with other research, enable a discussion of whether the cultural changes occurred in the eastern Pampa–Patagonia transition during the FLH had an impact on the health of hunter-gatherer groups adapted to semi-arid environmental conditions. Higher prevalences of caries, antemortem tooth loss, and dental calculus during the FLH indicates a number of changes in diet associated with a higher intake of carbohydrates. It is possible that an increased dento-alveolar lesion is accompanied by a change in food processing through the frequent use of pottery technology and grinding stones. Osteoarthrosic lesions indicate the increase in the distribution, frequency and severity in individuals during the FLH, suggesting differences in activity patterns developed by the groups. This is consistent with the proposed changes at the behavioral level in the groups that inhabited the area during this period (e.g. mobility, intensification of domestic activities). Porotic hyperostosis, cribra orbitalia, and linear enamel hypoplasias do not indicate differences through the Late Holocene. The main causes of its development could be linked to acquired anemia during childhood by a combination of factors. Poor nutrient intake and protein as a result of a decrease in the quality of breast milk due to prolonged breastfeeding and weaning could expose individuals to higher stages of infection and access of contaminated food. Also, exposure to gastrointestinal parasites during the life of individuals may have affected the health of populations. However, it is likely that the synergistic action of other factors (e.g. malnutrition, chronic diarrhea) played an important role in the expression of these stress markers. A low frequency of infectious processes was recorded during the Late Holocene. The proposed organizational changes probably were not sufficient to generate contexts for the development of infectious processes in bone tissue. Traumas were recorded in very low frequency in both periods and no changes were observed throughout the Late Holocene. The traumas were associated mainly to a combination of factors such as stress, accidents and, to a lesser extent by violence. It is proposed that the organizational changes and socio-demographic processes that occurred during the FLH (e.g. population growth, spatial circumscription, demographic packing, territoriality) would not have generated increases in social conflict and interpersonal violence. If social tensions existed in the study area and neighboring regions during the FLH, human populations would have been able to resolve tensions through other mechanisms such as population movements, group fissions and fusions, creating alliances, the exchange of goods and individuals, among others (see a similar proposal in Gordón, 2013).
In sum, the results of the pathological studies indicate that changes of health during the FLH were mainly related to subsistence and physical activities. There is an important difference between the quality and quantity of the sample in both periods, mainly due to the presence of multiple secondary burials and formal inhumation areas, and an increased population density during the FLH (Flensborg, 2012; Martínez et al., 2013). Despite these important differences, the pathological results suggest some trends. All results must be confirmed by the analysis of a larger sample, especially for the ILH. Also, a major spatial scale analysis will improve the interpretation of changes in health in a dynamic socioeconomic context.
This research was supported by CONICET (PIP-CONICET No. 6147/05) and the ANPCyT (PICT No. 264) projects. The INCUAPA-CONICET (Facultad de Ciencias Sociales, UNICEN) provided the facilities and support to carry out this analysis. Dr. Jorge Suby and Dr. Gustavo Martínez provided valuable comments and suggestions that improved the paper. Dr. Daniel Rafuse and Dr. Natalia Carden revised the English. Two anonymous reviewers made valuable comments.