Brief Communication
A female human skeleton from the Initial Jomon period found in the Iyai rock shelter in mountainous Kanto, Japan
2018 Volume 126 Issue 3 Pages 151-164

Details
2018 Volume 126 Issue 3 Pages 151-164
A new excavation of the Iyai rock-shelter site has uncovered more than a dozen human skeletal remains from the Initial Jomon period. We describe here an almost complete female skeleton (Iyai 1), and examine this in the context of morphological variation in Jomon females, especially those of the Initial Jomon period. Two radiocarbon dates based on the Iyai 1 skeletal samples show a calibrated date c. 8300–8200 calBP, belonging to the later part of the Initial Jomon period. The Iyai 1 skeleton was found in a burial pit with an unusual body placement. Although it was a flexed burial with the both arms and limbs flexed, the upper and lower body was disconnected at the waist and the two portions placed on top of each other: the upper body was laid on its abdomen with the head faced into the pelvis, and the lower body was found under the upper trunk in a supine position. The individual is a young female, 146 cm in height using Sasou–Fujii method, and the age at death is estimated as c. 20–40 years. The neurocranium is long (cranial index = 79.9) and high (length–height index = 77.1), the face is wide and low (upper-facial index = 43.4). The dentition shows nearly horizontal heavy wear but no dental caries with an edge-to-edge bite occlusion. The postcranial long bones are gracile, although they exhibit clear musculature impressions in some attachment areas. No paleopathological bone modification was found. The talus exhibits a ‘squatting’ facet at the anterior contact of the talocrural joint. The stable isotopic signature of Iyai 1 falls among those of the inland Initial Jomon people, indicating that her proteins mainly came from C3 plants and terrestrial animals.
The formation and establishment of the Jomon people and their culture has been one of the most intriguing issues in the study of the population history of Japan. Human skeletal remains from the earliest stage of the Jomon period, as well as those from the Late Paleolithic (Pleistocene) period, have thus acquired great importance. While we lack human skeletons from the Incipient Jomon period (c. 15700–11500 cal. BP), more than a dozen human skeletal individuals have been recovered from the Initial Jomon period (c. 11500–7000 cal. BP) (dates cited from Kobayashi, 2008). Recent re-evaluation of these Initial Jomon specimens (Nakahashi and Okazaki, 2009; Kohara et al., 2011; Kaifu et al., 2017) has aroused interest in the earliest stage of Jomon population history, although these specimens were all excavated in the 1960s. In terms of quantity of skeletal remains and quality of archaeological context, we are still awaiting new findings to improve our understanding of the Initial Jomon culture and its cultivators.
Morphologically, the Initial Jomon skeleton has been reported to be characterized by gracile postcranial bones with developed muscle attachment and unusual saddle-shaped dental wear (Suzuki, 1950), and smaller mandibles and limb bones than those of the later Jomon, with males having especially small upper limbs (Ogata, 1981). Yamaguchi (1982) suggested that the observed contrast between ‘gracile’ Initial–Early Jomon and ‘robust’ Middle–Final Jomon skeletons could be due to differences in regional or local environments at each site location. Ikeda (1985) considered both temporal and regional differences from mountainous cave sites and onshore shell-mounds, and developed a hypothesis that a ‘stable diet and associated labor intensification and community expansion’ in the later Jomon period led to robust upper limbs, especially in males. Recent assessment of the specimens found in the 1960s has confirmed the above observations, and added information of an extremely low and wide face (Nakahashi and Okazaki, 2009), a smaller mandible with anteriorly developed coronoid process (Kohara et al., 2011), and a presumably faster dental wear indicative of possible short lives (Kaifu et al., 2017).
In this situation, we undertook a new excavation of an Initial Jomon period site, the Iyai rock shelter, which uncovered a nearly complete female skeleton buried in a small pit. Here we report the burial, radiocarbon dating, and stable isotope analysis, and describe the female skeleton and compare its morphology with that of females from the Initial and other Jomon periods.
The Iyai rock-shelter site is located at Naganohara-cho (latitude 36°33′28″N, longitude 138°38′50″E) in Gunma Prefecture, at the periphery of the Joshinetsu mountains between the two main river systems, Shinano and Tone, in central Japan (Figure 1). The altitude is approximately 649 m, about 60 m above the present level of the nearby Shirosuna River. The rock shelter is carved on the surface of a welded tuff rock wall and it opens southeast to a tributary branch of the Agatsuma River.

Location of the Iyai rock-shelter site.
The Department of Archaeology, Kokugakuin University, started an archaeological campaign in 2014 (Taniguchi and Asakura, 2017) with the aim of elucidating the biology and ecology of the inhabitants of the beginning of the Jomon period. It revealed that the site itself expands from the inside of the shelter to the open sloping terrace in front and contains archeological remains from the Incipient Jomon to Late Yayoi periods. The project has conducted four seasons of excavations as of 2017 and plans to continue in the future.
The human skeleton Iyai 1 (IY1) was found in the summer of 2015. The occipital surface of the cranium was first exposed, then a series of vertebral spines and ribs were identified. After sampling a vertebral spine for radiocarbon dating, we stopped the excavation and the whole body was excavated in 2016. The burial of IY1 is described later.
Morphological diagnosis and comparisonHuman bones identified as IY1 were collected and packed with 66 separate recording labels. In the laboratory, each of them was cleaned, hardened, and then restored with acetone-dissolved BUTVAR-B76. Morphological observations are mainly based on human osteology textbooks (Buikstra and Ubelaker, 1994; White and Folkens, 2005). Sex diagnosis was based on the morphology of the pelvis (e.g. Phenice, 1969) and cranium. Age at death was estimated using age metamorphosis on the pubic symphysis (Brooks and Suchey, 1990) and on the auricular surface of the ilium (Buckberry and Chamberlain, 2002), and age changes in the cranial suture closure (Meindl and Lovejoy, 1985).
Cranial and postcranial measurements were taken according to Martin’s method (Baba, 1991). Facial flatness and nasal root protrusion were also measured according to Yamaguchi (1973) and Suzuki (1969). These characteristics were compared with those of contemporary and later Jomon individuals in addition to recent–modern Japanese individuals. Cranial and facial proportions or indices were compared in the context of an Initial–Early vs. Middle–Final Jomon comparison (Ogata, 1981). In order to visualize individual variations, a cranial length–breadth index and upper facial index were plotted by each measurement in Initial Jomon females among the whole range of Jomon females from the entire Japanese archipelago (Kondo et al., 2017). For postcranial bones, limb-length proportions were compared with those of East Jomon females (Takigawa, 2005), and limb robustness indices were calculated and compared with those of Initial–Early and Middle–Final Jomon series (Ogata, 1981).
Radiocarbon dating and stable isotopic analysisWe took small samples (0.1–0.2 g) from a vertebra of IY1, seven deer bones, and a raccoon dog bone to check background variation in stable isotope values. The soil matrix adhering to the bone surface was removed by sand blasting with aluminum oxide powder. Alkali and acid treatment was performed before gelatinization (Longin, 1971; Yoneda et al., 2002). Each bone piece was washed with 0.2 M NaOH in an ultrasonication bath to remove possible contamination by humic and fulvic acids. After ultrasonication in pure water and lyophilization, the pieces were individually crushed into a fine powder with a metal mortar and pestle. Bone powder was sealed in a cellulose tube to react with 1.2 M hydrochloric acid (at 4°C for 12 hours) to remove bioapatite and small molecular contaminants. Then, high molecular weight organic matter remaining in the tube was washed with pure water to neutral pH. The organic matter was recovered by centrifugation and heated in 10 mL of pure water to change collagen into gelatin at a temperature of 90°C for 12 hours. Gelatin solution was filtered with a glass filter to remove dissolved materials, and freeze-dried gelatin was used for radiocarbon dating and stable isotope measurements.
For radiocarbon dating, a target for accelerator mass spectrometry was reduced from CO2 produced from 2.5 mg of gelatin in a vacuum system. A mass spectrometer at The University Museum, The University of Tokyo was used to measure 14C/12C in 1 mg of carbon with international standard materials for correction (Omori et al., in press). Conventional radiocarbon age was converted into calibrated age in comparison with IntCal13 (Reimer et al., 2013) by using OxCal4.2 (Bronk Ramsey, 2009). Radiocarbon dating on the same individual was also conducted independently by PaleoLabo. Corp. Ltd, Japan, who applied their own protocols for collagen extraction and graphitization (Kobayashi et al., 2007).
Stable isotope ratios in carbon and nitrogen were measured by isotope ratio mass spectrometer (IRMS) combined with elemental analysis (EA) to produce N2 and CO2 sequentially. The 13C/12C and 15N/14N ratios were expressed in delta value units (δ13C and δ15N) in comparison with international standards such as Vienna Pee Dee Belemnite (VPDB) and ambient air (AIR). Uncertainties in the measurements were estimated with running standards as 0.1‰ for δ13C and 0.2‰ for δ15N. About 400 μg of gelatin was enfolded in a tin cup and used for EA-IRMS.
IY1 was buried in an oval pit belonging to the lower strata (unit II), a gray ash-colored deposit in the middle of the rock shelter. In addition to IY1, at least six other contemporary skeletons have been found so far, which suggests that the inner space of the rock shelter was used as a burial place by the inhabitants of the Initial Jomon period.
Unit II is a deposit about 50 cm thick formed in the Initial Jomon period. Artifacts belonging to the kaigara jokonmon (incised patterns made with shell) pottery phase in the later part of the Initial Jomon period were most abundant in this stratum. Chinsenmon (incised lines and punctures) potteries from the middle part of the Initial Jomon period were also found in this stratum.
Two radiocarbon dates reported below show a calibrated date of c. 8300–8200 calBP, belonging to the later part of the Initial Jomon period. Two small fragments of kaigara jokonmon potsherds were excavated in the grave of IY1. The radiocarbon dates on the skeleton clearly match the archaeological dates based on the pottery typology.
The skeleton IY1 was buried in a bent posture (Figure 2, Figure 3). At first sight, the body posture looked like a case of kussou (a burial posture with the arms and legs flexed), which is the most common burial posture in the Jomon period (Yamada, 2008). As more body parts appeared, however, we noticed that the burial was strange: the upper half and the lower half of the body were dismembered between the 3rd and 4th lumbar vertebrae, and the two parts were arranged separately. The bones of the upper body occupied the upper level of the burial pit and consisted of almost all items from the cranium to the lumbar vertebrae in the correct anatomical positions (Figure 3). Below them, the lower body appeared but the pelvis and two lumbar segments were found just under the cranium and placed at the bottom of corner of the pit. The position of the pelvis was thus distant from the lowest vertebrae of the upper body by approximately 40 cm (Figure 2).

Plan and cross-section of the IY1 burial.

IY1 skeleton in situ.
Although the reason for such an unusual burial body position is not clear at present, it is unlikely that the body was simply buried in the grave immediately after death. It can be also discounted that this could have happened by chance. Further, it cannot be categorized as a typical secondary burial because it shows a high level of correct anatomical connection and thus almost no cut marks in articulation. We should note that the size of the grave is about 70 cm long and 50 cm wide, which seems to be too tight for an adult body size. The normal size range for kussou graves is reported as 98–116 cm in length (Yamada, 2008). Taking this into consideration, it is hypothesized that the corpse was dismembered at the level of the waist and then put into this narrow grave in two parts. Although we cannot make a definitive image, the in situ body position of this burial matches findings of unusual burial customs in the Initial Jomon period.
A similar burial situation has been found in other examples. Reports of Initial Jomon skeletons from the Kamikuroiwa rock shelter (Nakahashi and Okazaki, 2009) have described the burial of two females, no. 4 Yanase-A and no. 5 Yanase-B, where the upper body of the former and the lower body of the latter (without duplication) were discovered piled on top of each other. Although the two body parts were reported as different individuals, they looked similar as they were different body parts of the same individual and each kept the correct anatomical positions. Another example is given in descriptions of burials found in the Hegi cave, where two individuals (no. 54 and 57) from the Initial Jomon period were reported to be buried with the bodies severed at multiple points (Kagawa, 1987). Interestingly, these two individuals were both cut at the waist.
In the case of IY1, identification as the same individual is essentially confirmed both from the burial situation (Figure 2, Figure 3) and from the morphological observations (see below). This new example of an unusual burial type from the Initial Jomon period, therefore, should prompt us to further investigate the data related to burial practices of the early Jomon people.
Skeletal morphology of IY1A detailed description of IY1 and linear measurements and indices are shown in the Appendix. The results of sex identification, determination of age at death, and the morphological comparisons are presented here.
IY1 was identified as a female based on the pubic morphology (Phenice, 1969) and diagnostic criteria of the os coxae and cranial superstructures. Smaller sizes of the postcranial bones are also indicative of a female. The age at death was estimated as young to middle age, roughly 20–40 years, based on combined results from the pubic symphysis (Brooks and Suchey, 1990), auricular surface of the ilium (Buckberry and Chamberlain, 2002), and cranial suture closure (Meindl and Lovejoy, 1985). The dental wear is heavy relative to the estimated age at death (Figure 4), which is in accordance with a previous report on Initial Jomon specimens (Kaifu et al., 2017). In contrast, the occlusal plane is mostly flat, not rounded or saddle-shaped, the latter of which is often observed in Initial Jomon dentition (Suzuki, 1950; Kohara et al., 2011). A possible antemortem dental chipping was diagnosable in the left upper first molar. The other molars and premolars exhibit a variable degree of cupped wear, where only the dentin was abraded while the enamel rim was intact. No dental caries was identified.

IY1 dentition. Inset: isolated right upper M3. Scale = 5 cm.
The cranium is almost complete, and moderate in size (Figure 5). The face is fairly wide and low, exhibiting common characteristics of members of the Initial Jomon period. Six cranial indices were compared with those of Initial–Early and Middle–Final Jomon females (Ogata, 1981) after being standardized by means and standard deviations of Kanto Japanese females of recent–modern times (Morita, 1950) (Figure 6). Compared with the standards of the Kanto Japanese females, the cranial length–breadth index of IY1 (8/1) is quite similar to that of Kanto Japanese and that of the later (Middle–Final) Jomon. The negative deviation of the Initial–Early Jomon denotes an anteroposteriorly longer cranium. Among females of the whole Jomon period, IY1 has a cranial index at 0.8 (borderline between meso- and brachycrany), and exhibits a broader cranium than other Initial Jomon females (Figure 7). The length–height (17/1) and breadth–height (17/8) indices of IY1 are both smaller than those of the Kanto Japanese but a little larger than the average of Initial–Early Jomon females (Figure 6). Large negative deviations of IY1 in the upper facial Kollmann index (48/45) and orbital index (52/51) indicate a wide and broad face and orbits. In the individual plot of breadth (M45 ZYB) vs. height (M48 NPH), IY1 falls into the lowest position among the variations (Figure 8). The mandible also exhibits a shared Jomon characteristic of a broader ramus compared with the height (Figure 6), where the ramus index (71/70) of IY1 deviates positively as found in Initial–Early and Middle–Final Jomon females.

IY1 skull. Scale = 10 cm.
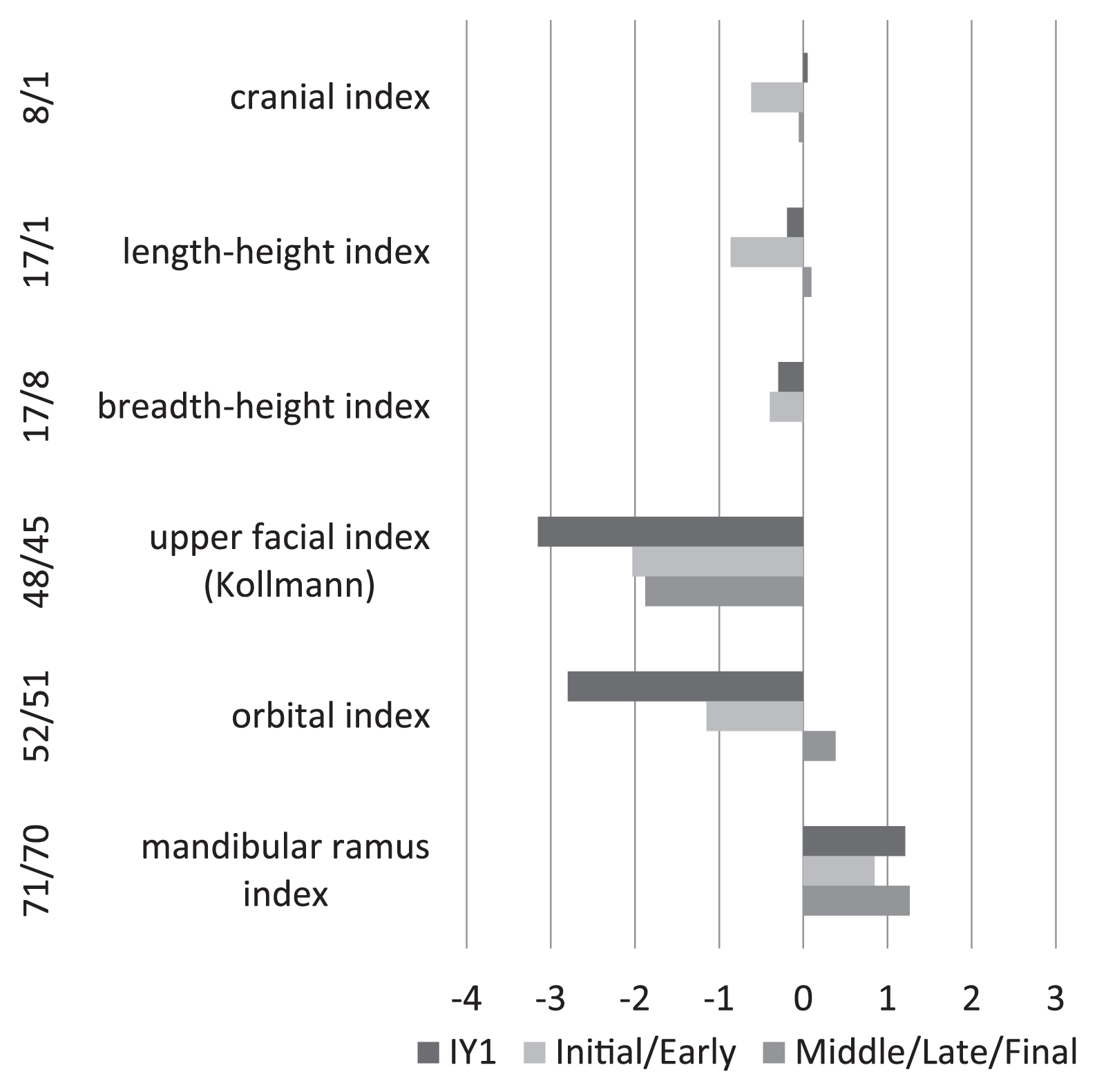
Comparison of the cranial indices of Iyai 1 (IY1), and those of Initial–Early (n = 4–8) and Middle–Final Jomon (n = 19–36) females (Ogata, 1981), standardized by means and standard deviations of Kanto Japanese females (Morita, 1950).

Comparison of cranial shape: plots of length (GOL) vs. breadth (XCB) between Initial Jomon (red) and Early–Final Jomon (dots) females. Lines are XCB/GOL indices of 0.75 and 0.8, respectively. IY1, Iyai 1 (present study); Kam, Kamikuroiwa (Nakahashi and Okazaki, 2009); Yug, Yugura (Morimoto and Takahashi, 1986); Nak, Nakatsugawa (Dodo, 1976); Toc, Tochibara (Kohara et al., 2011); Shi, Shironodai-Minami (Shigehara, 1994).
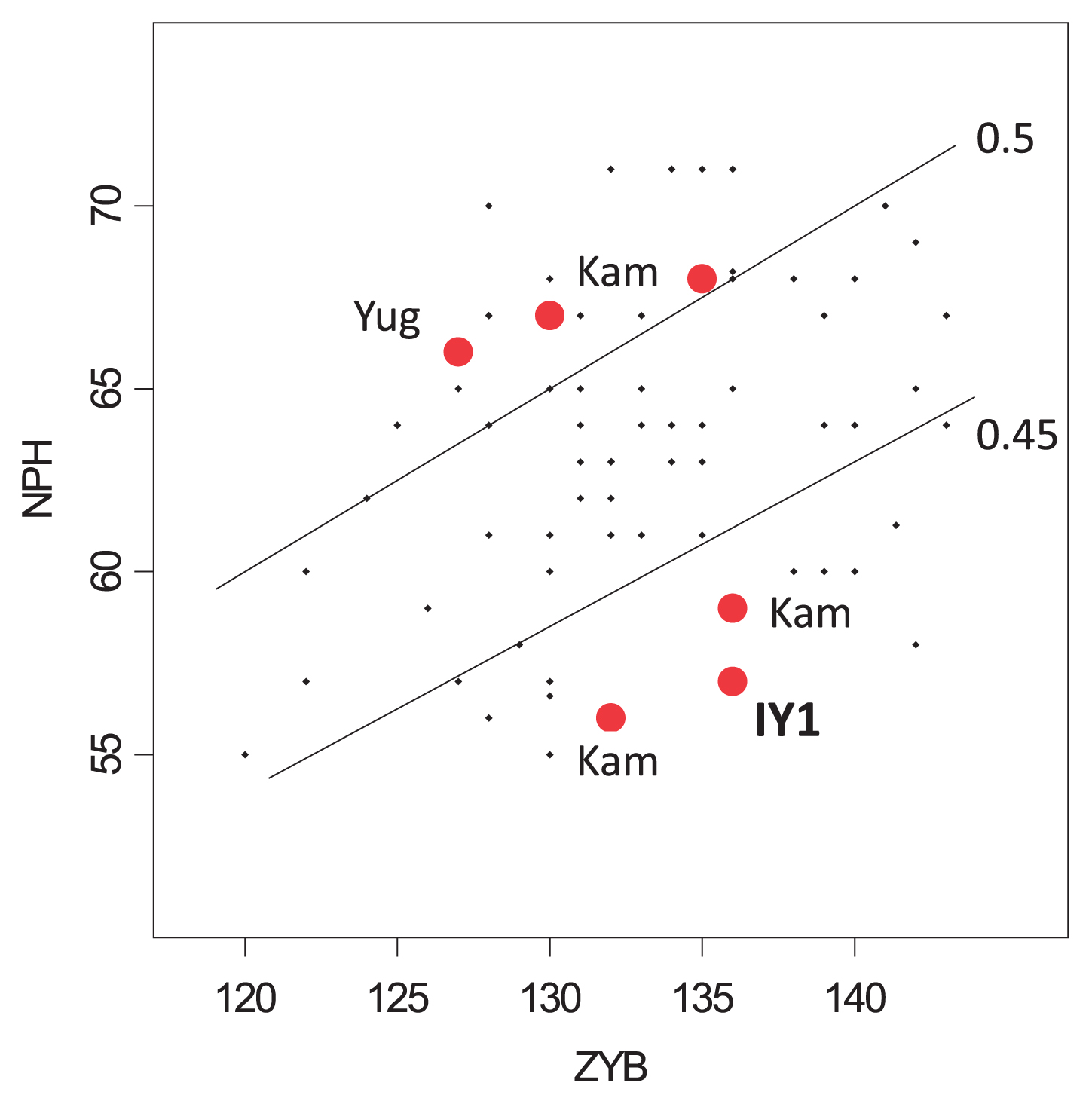
Comparison of upper face proportions: plots of breadth (ZYB) vs. height (NPH) between Initial Jomon (red) and Early–Final Jomon (dots) females. Lines are NPH/ZYB indices of 0.45 and 0.5, respectively. IY1, Iyai 1; Kam, Kamikuroiwa; Yug, Yugura (references in Figure 7).
The postcranial remains of IY1 are almost complete (Figure 9). They show almost no signs of age-related or pathological alterations. Their morphology largely fits the characteristics already reported in Initial Jomon specimens, that is, they are gracile but exhibit strong muscle-attachment impressions. We confirmed that the limb bones of Initial–Early Jomon females were more gracile than those of Middle-to-Final ones (Figure 10). Among those gracile Jomon specimens the clavicle of IY1 is quite long and thus gracile, and the other limb bones exhibit a variable degree of gracility. The interlimb proportions of IY1 were compared with those of East Jomon and Kanto Japanese females (Takigawa, 2005) (Figure 11). The deviations of IY1 from the Kanto Japanese average are all directed to the same direction as those of East Jomon females. This means the proportional characteristics in limb-bone lengths are common between IY1 and East Jomon females, i.e., they have a longer clavicle, longer forelimb and longer lower thigh, and a shorter humerus compared with the femur.

IY1 postcranial bones. Scale = 10 cm.

Comparison of limb robustness of Iyai 1 (IY1), and those of Initial–Early (n = 3–6) and Middle–Final Jomon (n = 10–22) females (Ogata, 1981), based on Kanto Japanese females (Kohara et al., 2011). The limb robustness is measured as a ratio of midshaft diameter (circumference or sum of diameters) per maximum length.

Comparison of limb proportion indices of Iyai 1 (IY1), and those of East Jomon females (Takigawa, 2005), standardized by means and standard deviations of Kanto Japanese females (Takigawa, 2005). The index of limb segments is calculated based on the maximum lengths.
The stature was estimated based on the limb bone lengths. According to the Sasou–Fujii method (Sasou and Hanihara, 1998) using limb lengths of both sides, a mean value of 145.7 cm was calculated. A second stature estimate, calculated using the formulae detailed by Pearson (1899), which was formerly used for Jomon stature estimation, was 148.2 cm.
Radiocarbon dates and stable isotope ratiosThe quality of gelatin was estimated using carbon and nitrogen contents, and its atomic C/N ratio, as well as the gelatin yield from bone powder. The yield of 9.5% in human bones and 1–6% from animal bones, with the exception for one deer (0.1%), suggests the collagen was well preserved. The C/N ratios for IY1 had a value of 3.2 as collagen, and animals between 3.2 and 3.6, which are in the normal biological range (2.9–3.6; DeNiro, 1985). Additionally, the higher contents of carbon and nitrogen suggest that extracted gelatin samples suffered little alteration and contamination. Hence, we could conclude that the radiocarbon dates and stable isotope ratios in gelatin are reliable.
The radiocarbon age of the IY1 human skeleton was dated as 7413 ± 28 years BP (TKA-16437, conventional uncalibrated age), corresponding to 8323–8180 calBP (95.4%), as shown in Table 1. This result is very similar to the radiocarbon date of 7427 ± 28 years BP (PLD-31614) measured by an independent commercial laboratory, suggesting the robustness of the age determination of this specimen. The calibrated radiocarbon age supports the stratigraphic observation at the site that this burial originated in the late part of Initial Jomon period (11500–7000 calBP; Kobayashi, 2008).
| Sample | Lab. ID | Conventional 14C age | Calibrated age (1 sigma) | Calibrated age (2 sigma) |
|---|---|---|---|---|
| IY1 | TKA-16437 | 7413 ± 28 BP | 8304–8242 calBP (45.8%) 8217–8186 calBP (22.4%) |
8323–8180 calBP (95.4%) |
The carbon and nitrogen isotopic ratios of –20.0‰ and 7.6‰ suggest only negligible contribution from aquatic protein and C4 plants in the last 10 years of her life (Table 2). If the IY1 individual consumed aquatic animals including freshwater fish and anadromous salmon, her nitrogen isotopic values would be expected to be higher in both the carbon and nitrogen isotope ratios. As the δ15N was higher in IY1 than in the deer and raccoon dog, with little difference in δ13C, this individual obtained protein mainly from terrestrial animals (Figure 12). However, the estimated range of diet is slightly higher in δ13C than those of deer, which suggests that IY1 did not solely obtain protein from deer meat. The diet range was estimated according to the enrichment factor for δ15N (3.8 ± 1.1‰) and δ13C (1.1 ± 0.2‰) in collagen (Bocherens, 2015). The possible nutritional resources for human at the Iyai rock shelter should be discussed in conjunction with faunal and floral data.
| sample ID | species | gelatin yield | %C (wt) | %N (wt) | atomic C/N | δ13C (VPDB) | δ15N (AIR) |
|---|---|---|---|---|---|---|---|
| IY1 | human | 9.5 | 44.9 | 16.2 | 3.2 | −20 | 7.6 |
| IY-Z01 | shika deer | 5.9 | 44.3 | 15.9 | 3.2 | −22.6 | 1.9 |
| IY-Z02 | shika deer | 5.3 | 43.7 | 15.7 | 3.2 | −22.7 | 3.3 |
| IY-Z03 | shika deer | 6.0 | 44.6 | 16 | 3.3 | −22.4 | 2.2 |
| IY-Z04 | shika deer | 1.0 | 42.1 | 13.5 | 3.6 | −23.6 | 3.5 |
| IY-Z05 | shika deer | 3.0 | 45.1 | 15.6 | 3.4 | −21.9 | 2.6 |
| IY-Z06 | shika deer | 5.0 | 44.5 | 16.1 | 3.2 | −23.1 | 3.2 |
| IY-Z07 | shika deer | 4.0 | 43.6 | 15.4 | 3.3 | −22 | 3.4 |
| IY-Z08 | black bear | 0.07 | ND | ND | ND | ND | ND |
| IY-Z09 | racoon dog | 2.8 | 42.1 | 14.6 | 3.4 | −19.2 | 5.5 |
ND, not determined.

Carbon and nitrogen isotope ratios in human (IY1) and its expected diet range in comparison with those of deer and raccoon from the Iyai rock shelter.
The carbon and nitrogen isotope ratios of IY1 were compared with those previously reported in Initial Jomon populations (Figure 13). The position of IY1 suggests a similar isotopic signature with inland sites such as the Tochibara rock shelter in Nagano Prefecture (Yoneda et al., 2002), the Kamikuroiwa rock shelter in Ehime Prefecture (Gakuhari et al., 2015), and Iwashita Cave in Nagasaki Prefecture (Yoneda et al., 2017). In contrast, the Higashimyo shellmidden in Oita Prefecture (Yoneda et al., 2016) showed significant contributions from the aquatic ecosystem, suggesting a possible high variety of subsistence in the Initial Jomon period as it adapted to early Holocene conditions. However, it is worth noting that no Jomon individuals, including IY1, showed any isotopic signatures of C4 plants, including foxtail, barnyard and broomcorn millet, and beans with N2-fixing rhizobia in this period.

Carbon and nitrogen isotope ratios in humans from the Initial Jomon cultural layers.
A new excavation of the Iyai rock shelter has uncovered both archaeological and anthropological data from the Initial Jomon period, which is valuable for understanding the earliest stage of Jomon culture and its cultivators. A well-preserved female skeleton, IY1, aged 20–40 years, and exhibiting similar morphological characteristics to Initial Jomon people, including a low and wide face, smaller mandible with a shorter ramus, heavy dental wear but without caries, and gracile limb bones with strong muscle impressions, was found. The stable isotopic signature of IYI falls among those of the inland Initial Jomon people, indicating that her proteins mainly came from C3 plants and terrestrial animals. The burial of IY1 was strange: the lower body seemed to have been dismembered at the waist and buried separately. As mentioned, similar examples of disconnected corpses have been found in a few of the earliest Jomon sites, e.g. the Hegi cave-site in Oita Prefecture and the Kamikuroiwa rock shelter in Ehime Prefecture. More research should be carried out on such mortuary practices as well as further anthropological observation of Jomon skeletal remains. All the information can then be integrated to help to understand the prehistoric Jomon people and their behavior.
We thank Kazutaka Asakura, Momoko Saga, Kanako Inokuma, Asuka Chihara, Kousaku Matsumoto, Takuya Iino, and Zen Minamidate for their collaboration and cooperation in the fieldwork of the Iyai rock-shelter site. We also appreciate Kyomi Yamazaki’s contribution in sampling animal bone remains for stable isotope analysis. This research project is supported by JSPS (no. 17H00939, jointly nos. 15H02946 and 16K07530).
The pubic portion of the os coxae was preserved at both sides, where the sexually dimorphic traits of ventral arc, subpubic concavity, and the medial aspect of the ischiopubic ramus all appear as feminine (Phenice, 1969). In addition, the greater sciatic notch widely opens and the preauricular sulcus exists. The cranium of IY1 exhibits a weak nuchal profile and a weak glabellar protrusion with a sharp edge of the upper orbital margin although the mastoid process is relatively large. The mandibular morphology expresses a feminine pointed chin.
The age at death of IY1 was estimated as young to middle age. The surface on the pubic symphysis starts to diminish in horizontal furrows and exhibits a ventral beveling (ventral rampart), the oval outline of the symphyseal face comes to completion, although bony outgrowth or upper or lower nodules are faint, all matching phase 3 or 4 of the Suchey–Brooks method (Brooks and Suchey, 1990). The age metamorphosis on the auricular surface is grouped into Buckberry’s stage 3 (Buckberry and Chamberlain, 2002), showing faint horizontal wrinkles on less than half of the auricular surface, fine granularity evident over most of the surface, with limited micro- and macroporosity, and a sharp apex without any outgrowth. Both of these methods indicate the estimated age at death to be c. 20–40. Cranial suture scores by Meindl and Lovejoy (1985) are score 3 in the vault (the suture closure starts at the obelion, anterior-sagittal and pterion) and score 1 in the lateral-anterior portion (only the pterion), which indicate almost the same age, i.e. the 30s. These all indicate that the individual IY1 probably died at around 30 years of (range 20–40 years).
Cranium (see Figures 4–8)The cranium was almost complete (Figure 5). The cranial index (max. length/max. breadth) is 79.9, which belongs to the upper range of mesocrany (Figure 7). The neurocranium is high, the length–height index is 77.1 (hypsicrany) and the breadth–height is 96.5 (metriocrany). The frontal bone exhibits clear temporal lines with a relatively strong postorbital narrowing. The supraorbital rims exhibit notches (no foramina) for the supraorbital nerves. The face is wide and low, the upper facial Kollmann index is 43.4, indicating a very wide or broad face (hypereuryeny) (Figure 8). The orbital rims are also wide and low, the orbital index is 70.0 (chamaeconchy). The nasal aperture is wide, the nasal index is 57.8 (platyrhiny) with a blunt lower marginal rim. In contrast to the wide and low face, which is common with the other Jomon and particular the Initial Jomon people, the degrees of facial flatness and nasal root protrusion are flatter and weak in IY1. The maxilla shows a shallow canine fossa; the maxillary alveolar process does not protrude anteriorly. The dentition occludes in edge-to-edge bite. The temporal bone possesses wide and shallow mandibular fossae without traces of mandibular joint inflammation. In the basal view, a tympanic dehiscence (foramen Huschke) is observed at both sides. The mandible is small but the superstructure looks robust. The corps is continuously low from the posterior to the mid-symphyseal portion. The mental protuberance points anteriorly. The ascending ramus is wide and low (ramus index = 61.4).
The dental remains lack the right maxillary second molar (M2), but all the other permanent teeth were preserved (Figure 4). The right upper M2 seems to have been lost before death because the root socket is completely obliterated, but it might have survived up to near or at the time of death because the adjacent upper M1 and M3 show clear approximal facets to the M2, and the mandibular corresponding molars show horizontal wear. The dental wear is heavy relative to the estimated age at death and the occlusal plane is almost flat in the entire dentition. All the dental crowns from the anterior to posterior teeth expose the dentine on the occlusal surface. In the lower medial incisors, the enamel is worn off in all the crown outer rims. The degree of wear corresponds to Smith stages 6–7 (Smith, 1984) for molars. The enamel only survives around the rim in most molars and the inside dentin surface is concave, i.e. showing cupped teeth. The occlusal surface of the left upper M1 is lingually inclined, although the corresponding one in the lower teeth is horizontal, which suggests that the left upper M1 was chipped off at portions of the lingual cusp before death and the wear was advanced around the chipping.
The skull was accompanied by the ear ossicles (right malleus and left incus) and the hyoid.
Trunk of the body (see Figure 9)All the bones of the trunk were well preserved, and we could identify all the vertebrae from cervical to lumbar sections, with 12 rib pairs at both sides and the sternum. There is no senile bony outgrowth at the synchondrodial joints between the rib and the sternum.
In the vertebral column, a sequential connection is observed at the intervertebral and zygapophysial joints from the top of the atlas to the lower 3rd lumbar vertebra. The columnar segments from the 12th thoracic to the 3rd lumbar bodies continuously lack the left-anterior portion of the body, suggesting a physical or chemical erosion on the mass of the conjoint vertebrae. In contrast, the 4th lumbar vertebra exhibits a large portion of damage from the upper half of the body to the pedicle root. The columnar connection is interrupted at this point. Below this, the 4th and 5th lumbar and the 1st sacral connection are confirmable. The lower connecting bodies also show a large deletion of the left-anterior and right-anterior portions of the vertebral body. There is no lipping formation on all the observable intervertebral articular facets.
The pelvis is almost complete, preserving both the os coxae and the sacrum. These show feminine characteristics and young to middle-aged metamorphosis on the pubis symphysis and the auricular surface.
Upper and lower limbs (see Figure 9)Bones of the upper limb are well preserved with identification of the clavicle, scapula, humerus, radius, and ulna, as well as the hand bones at both sides. The clavicle is straight and long with strong impressions for muscle attachment of the deltoid and subclavius muscles. A strong concavity of the anterior wall near the sternal end of the shaft relates to the origin of the pectoralis major muscle. The scapula retains a middle-sized glenoid cavity, and possesses a relatively large distance from the glenoid to the acromion. The boundaries of the muscle attachments form strong ridges. The humerus is almost complete, but the left humeral shaft possesses a trauma running obliquely on the posterior-lateral surface distal to the radial nerve groove. From this point distally, the surface is partially worn and eroded to the lateral epicondyle. The trauma is a shallow and wide V-shape, seemingly not a typical ‘cutmark’ by a sharp lithic object but a trace of the impression of a straight object. The muscle attachment for the pectoralis major and the deltoid is developed, but the shaft itself is gracile with a smaller proximal caput and distal articular end. The cross-sectional index at the midshaft indicates a relatively flat diaphysis of the humerus (right = 71.5, left = 76.1). Both radius and ulna are gracile and small in size but exhibit developed interosseous margins, which may indicate a high level of muscle function. The distal portion of the ulna shows a well-developed ridge for the pronator quadratus muscle, and thus the distal shaft itself curves outwards. Among the well-preserved hand bones, the following are identified: 4 carpals (trapezoid, capitate, hamate, pisi-form), 1st–5th metacarpals, and 13 hand phalanges in the right hand, 6 carpals (trapezium, capitate, hamate, lunate, triquetrum, pisiform), 1st–5th metacarpals, and 12 hand phalanges in the left hand. There was no pathological alteration, such as lipping formation, on the articular surface.
Lower limb bones are also well preserved, although portions around the left knee are largely lacking and several length measurements are impossible to obtain. Identified bones include the femur (both sides), patella (left), tibia (both sides), and fibula (both sides), as well as the bones of the foot. The femur is complete except for the left distal articulation. The length almost approaches the average for Jomon females, and the head and the distal articular portion are small. The shaft looks typically ‘pilasteric’ with a modest degree of pilaster development. The anterior–posterior curvature is moderate. The cross-sectional index (113–114) at the midshaft falls at around the average for Jomon females. The proximal shaft develops a gluteal tuberosity and a lateral flange of the shaft, forming a flatter femur with a platymeric index (73.7 right, 79.2 left), which belong to a ‘platymeric’ femur. The tibia only lacks a portion of the tibial plateau at the left proximal end. The diaphysial surface of the right tibia suffers a degree of erosion near both ends. The posterior surface of the shaft holds a vertical line of the tibialis posterior muscle. The midshaft cross-section thus becomes rhomboid, but the flatness is weak in comparison with that typically found in Jomon people. The platycnemic index at the nutrient foramen (63–64) falls in the lower range (near the platycnemic) of ‘mesocnemy.’ The fibula is complete except for the left proximal end, and has a wide diaphysial shaft and a longitudinal depression (‘gutter’) on the lateral surface of the shaft. As for the foot bones, all the tarsals, metatarsals, and proximal phalanges are identified. The talar trochlea exhibits a higher lateral surface than the medial one, which is typical of that found in Jomon skeletons. At the neck of the talus, a so-called ‘squatting’ small facet or bony swelling is observed on both sides. At the counterportion of the distal surface of the tibia, a depression is formed instead of a pseudoarticular facet. Both calcanei are small and gracile, possessing a demarcated type of anterior talar facet. The long axis of the posterior talar articular facet is tilted against the long axis of the calcaneus.
| Cranial measuement | Cranial index | ||||
|---|---|---|---|---|---|
| M1 | max. cranial length (GOL) | 179 | 8/1 | cranial length–breadth index | 79.9 |
| M5 | cranial base length | 103 | 9/8 | transverse front-parietal index | 67.8 |
| M8 | max. cranial breadth (XCB) | 143 | 9/10 | transverse frontal index | 82.2 |
| M9 | min. frontal breadth | 97 | 17/1 | cranial length–height index | 77.1 |
| M10 | max. frontal breadth | 118 | 17/8 | cranial breadth–height index | 96.5 |
| M11 | biauricular breadth | 125 | 20/1 | length–(po-b height) index | 64.8 |
| M12 | max. occipital breadth | 104 | 20/8 | breadth–(po-b height) index | 81.1 |
| M17 | basion–bregma height | 138 | 48/45 | upper facial index (Kollmann) | 43.4 |
| M20 | porion–bregma height | 116 | 48/46 | upper facial index (Virchow) | 60.2 |
| M26 | frontal sagittal arc | 118 | 52/51 | orbital index | 70.0 |
| M29 | frontal sagittal chord | 107 | 54/55 | nasal index | 57.8 |
| M40 | basion–prosthion length | 98 | 71/70 | mandibular ramus index | 61.4 |
| M43 | upper facial breadth | 104 | |||
| M44 | biorbital breadth | 97 | |||
| M45 | bizygomatic breadth (ZYB) | (136) | Facial flatness measurement (Yamaguchi, 1973) | ||
| M46 | bimaxillary breadth | 98 | frCH | frontal chord | 98.0 |
| M48 | upper facial height (n-pr) (NPH) | 57 | frSUB | frontal subtense | 9.9 |
| upper facial height (n-av) | 59 | frontal flatness index | 10.1 | ||
| M51 | orbital breadth (mf) | 40 | sCH | simotic chord | 8.9 |
| M51a | orbital breadth (d) | 39 | sSUB | simotic subtense | 1.2 |
| M52 | orbital height | 28 | simotic flatness index | 13.5 | |
| M54 | nasal breadth | 26 | zmCH | zygomaxillary chord | 98.0 |
| M55 | nasal height | 45 | zmSUB | zygomaxillary subtense | 18.8 |
| M60 | maxilloalveolar length | 51 | zygomaxillary flatness index | 19.2 | |
| M61 | maxilloalveolar breadth | 60 | |||
| Mandibular measurement | Nasal root measurement (Suzuki, 1969) | ||||
| M65 | bicondylar breadth | 125.6 | M50 | nasal root breadth | 22.6 |
| M66 | bigonial breadth | 96.0 | IA | nasal root arc | 26.0 |
| M68 | mandibular length | 76.0 | 50/IA | prominent index | 86.9 |
| M69 | symphysis height | 27.5 | |||
| M70 | ramus height | 57.0 | |||
| M71 | ramus breadth | 35.0 | |||
| M69(2) | body height (M2) | 25.2 | |||
| M69b | body breadth (M2) | 16.6 | |||
Note: estimated value in parentheses.
| Right | Left | Right | Left | ||||
|---|---|---|---|---|---|---|---|
| Clavicle | M1 max. length | 142.0 | 138.0 | Femur | M1 max. length | 390.0 | — |
| M6 midshaft circumference | 31.0 | 31.0 | M2 physiological length | 388.0 | — | ||
| 6/1 robustness index | 21.8 | 22.5 | M6 sagittal shaft diameter | 26.3 | 26.4 | ||
| M7 transverse shaft diamter | 23.3 | 23.2 | |||||
| Scapula | M12 glenoid fossa length | 29.3 | 28.4 | 6/7 cross-sectional index | 112.9 | 113.8 | |
| M13 glenoid fossa breadth | 21.8 | 22.1 | M8 midshaft circumference | 78.0 | 79.0 | ||
| M13-01 acromioglenoid distance | 21.2 | — | M9 transverse subtrochanteric diameter | 28.9 | 28.4 | ||
| M10 sagittal subtrochanteric diameter | 21.3 | 22.5 | |||||
| Humerus | M1 max. length | 275.0 | 272.0 | 10/9 platymeric index | 73.7 | 79.2 | |
| M4 biepicondylar width | 46.2 | 46.3 | M18 vertical head diameter | 38.9 | 39.1 | ||
| M5 max. diameter of midshaft | 20.0 | 18.8 | M19 antero-posterior head diameter | 38.7 | 38.8 | ||
| M6 min. diameter of midshaft | 14.3 | 14.3 | M21 bicondylar width | 65.2 | — | ||
| 6/5 cross-sectional index | 71.5 | 76.1 | |||||
| M7 min. circumference | 52.0 | — | Patella | M1 max. height | — | 36.6 | |
| 7/1 robustness index | 18.9 | — | M2 max. breadth | — | 41.6 | ||
| Radius | M1 max. length | 205.0 | 203.5 | Tibia | M1 total length | 314.0 | — |
| M4 max. transverse shaft diameter | 14.7 | — | M1a max. length | 321.0 | — | ||
| M5 min. sagittal shaft diameter | 9.9 | — | M8 max. diameter at midshaft | 26.1 | 26.6 | ||
| 5/4 cross-sectional index | 67.3 | — | M9V transverse diameter (Vallois) | 19.1 | 18.9 | ||
| 9V/8 cross-sectional index | 73.2 | 71.1 | |||||
| Ulna | M1 max. length | 216.5 | 221.0 | M8a max. diameter at nutrient foramen | 29.4 | 29.8 | |
| M11 dorsoventral shaft diameter | 13.1 | 11.7 | M9aV transverse diamter (Vallois) | 18.8 | 18.9 | ||
| M12 transverse shaft diameter | 13.5 | 13.1 | 9aV/8a platycnemic index | 63.9 | 63.4 | ||
| 11/12 cross-sectional index | 97.0 | 89.3 | M10 midshaft circumference | 72.0 | 77.0 | ||
| M10b min. circumference | 64.5 | 64.0 | |||||
| Fibula | M1 max. length | 315.0 | — | ||||
| M2 max. diameter at midshaft | 15.9 | 14.7 | |||||
| M3 min. diameter at midshaft | 8.6 | 8.7 | |||||
| 3/2 cross-sectional index | 54.1 | 59.2 | |||||
| M4 midshft circumference | 40.0 | 39.0 | |||||