Material Reports
Relationship between the calcaneal size and body mass in primates and land mammals
2019 Volume 127 Issue 1 Pages 73-80

Details
2019 Volume 127 Issue 1 Pages 73-80
The relationship between calcaneal size and body mass in extant primates and other land mammals is examined using regression analyses to provide simple equations for estimating the body mass of extinct primate and land mammal species based on the calcaneus. The results imply that among the linear calcaneal dimensions, the calcaneal width at the talar articular surfaces (CA2) is likely the best body mass estimator for land mammals (including primates), and the width of the posterior talar articular surface (CA3) appears to be relatively good body mass estimator for primates. The equation with a 95% prediction interval for estimating the body mass (BM, in g) using CA2 (in mm) for land mammals is: BM = exp(2.928 × ln CA2 + 0.981 ± 0.772) × 1.076; the corresponding equation using CA3 (in mm) for primates is: BM = exp(2.555 × ln CA3 + 3.536 ± 0.641) × 1.067.
The heelbone (calcaneus or calcaneum) and anklebone (talus or astragalus) of primates and other mammals have been well studied in primatology, anthropology, archaeozoology, and vertebrate paleontology as indicators of functional adaptation, phylogeny, and taxonomy (e.g. Szalay, 1977; Gebo et al., 1991; Dagosto and Terranova, 1992; Ciochon et al., 2001; Ciochon and Gunnell, 2004; Gebo and Dagosto, 2004; Gunnell and Ciochon, 2008; Polly, 2008; Marivaux et al., 2010; Jogahara and Natori, 2013; Tsubamoto, 2014; Tsubamoto et al., 2016 and references therein). The bony structure and shape of these two bones are relatively compact and robust (e.g. Gray, 1858), so that these bones are frequently preserved and are found undamaged more often than long bones, vertebrae, or fragile skulls in the fossil and zooarchaeological assemblages (Tsubamoto, 2014; Tsubamoto et al., 2016). The body mass of primates and other mammals, on the other hand, is a useful predictor of species adaptations and diversities because it is strongly correlated with many aspects of life history, ecology, and behavior, etc. (e.g. Peters, 1983; Calder, 1984; LaBarbera, 1989). Therefore, estimates of the body mass of extinct mammalian species play an important role in paleoecological, paleoprimatological, and physical anthropological analyses (e.g. Legendre, 1986, 1989; Conroy, 1987; Anyonge, 1993; Fleagle, 1999; Egi, 2001; Smith et al., 2010; Grabowski et al., 2015; Jungers et al., 2016; Tsubamoto et al., 2016; Ruff and Niskanen, 2018 and references therein). Several studies have investigated the relationship between talar size and body mass in primates and other mammals (Dagosto and Terranova, 1992; Martinez and Sudre, 1995; Rafferty et al., 1995; Polly, 2008; Parr et al., 2011; Tsubamoto, 2014; Yapuncich et al., 2015; Tsubamoto et al., 2016; Dagosto et al., 2018). However, only a few studies have investigated the relationship between calcaneal size and body mass: for example, Dagosto and Terranova (1992) did it for ‘prosimian’ primates (strepsirrhines and Tarsius); Yapuncich et al. (2015) did it for euarchontans (primates, scandentians, and dermopterans); and Dagosto et al. (2018) did it for euarchontans and small mammals.
In this material report, the relationships between the calcaneal size and body mass in living primates and land mammals are examined using regression analyses. The purpose of this report is to provide simple equations that will allow paleoprimatologists and vertebrate paleontologists to estimate the body masses of extinct primate and land mammal species from the calcaneal size. The original data used here are limited, and hence this report should be treated as a pilot study simply for the body mass estimation from the calcaneus. Any other functional signals for, for example, behavior and ecology are not discussed here.
The original data used in this study consist of the body mass and 12 linear measurements of the calcaneus of superficially 69 individuals, representing 44 species belonging to 10 orders of extant land mammals, and ranging in body mass from 18 g to 1.4 tonnes (Table 1). Most of the samples are adult individuals with three subadult specimens. These three subadult specimens (Table 1) are used to increase the data points. Most of the individuals used here are the same individuals used by Tsubamoto (2014) and Tsubamoto et al. (2016). The body masses represent the actual body weight of the individual of each specimen and were recorded either while the animals were still alive or just after their death (Tsubamoto, 2014). The data (body mass and 12 linear measurements) for the adult males and females of Japanese monkey (Macaca fuscata fuscata: Primates, Cercopithecidae) and Japanese raccoon dog (Nyctereutes procyonoides viverrinus: Carnivora, Canidae) were derived from the mean values of more than 25 specimens for each sex of each species (Table 1). Owing to the limited availability of specimens for which such data could be obtained, the dataset is somewhat biased towards primates and carnivores (Table 1; Tsubamoto, 2014). The 12 linear measurements (CA1–CA12) are indicated in Figure 1 and Table 2. The units of the linear measurements and body mass are in millimeters (mm) and grams (g), respectively (Table 1).
| Higher taxa | Specimen no. | Species | BM (g) | Sex | CA1 | CA2 | CA3 | CA4 | CA5 | CA6 | CA7 | CA8 | CA9 | CA10 | CA11 | CA12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Primates | NSM-M 31595 | Gorilla gorilla | 216000 | M | 95.89 | 50.57 | 28.25 | 25.05 | 39.99 | 45.92 | 33.36 | 31.76 | 23.89 | 41.66 | 36.10 | 49.42 |
| Primates | KUPRI 8135 | Hylobates agilis | 8000 | F | 27.71 | 15.31 | 8.26 | 7.12 | 8.15 | 8.44 | 11.17 | 9.91 | 7.53 | 13.94 | 13.94 | 17.63 |
| Primates | NSM-M 32559 | Pan troglodytes | 50000 | F | 53.83 | 32.41 | 18.46 | 18.22 | 19.37 | 20.13 | 25.58 | 22.99 | 14.16 | 30.60 | 25.69 | 30.46 |
| Primates | NSM-M 33042 | Pan troglodytes | 43600 | F | 54.39 | 32.72 | 18.17 | 17.78 | 19.74 | 21.48 | 22.47 | 22.71 | 15.42 | 29.94 | 24.81 | 29.86 |
| Primates | NSM-M 31996 | Pongo pygmaeus | 61000 | F | 58.35 | 31.81 | 16.36 | 11.48 | 14.88 | 20.76 | 20.74 | 19.52 | 14.87 | 25.14 | 22.17 | 30.61 |
| Primates | KUPRI 4237 | Erythrocebus patas | 5000 | F | 33.38 | 14.31 | 7.40 | 6.44 | 9.59 | 12.18 | 8.87 | 10.05 | 7.43 | 13.27 | 10.10 | 11.52 |
| Primates | * | Macaca fuscata | 10245 | M | 38.05 | 17.35 | 9.28 | 8.78 | 11.66 | 13.63 | 11.75 | 12.52 | 8.87 | 16.25 | 14.05 | 16.89 |
| Primates | ** | Macaca fuscata | 7119 | F | 34.46 | 15.96 | 8.45 | 7.66 | 10.47 | 12.03 | 10.86 | 11.39 | 8.15 | 14.73 | 12.88 | 15.36 |
| Primates | KUPRI 1626 | Papio anubis | 31400 | M | 48.44 | 23.97 | 13.91 | 13.99 | 17.10 | 17.93 | 16.30 | 17.22 | 13.19 | 23.34 | 21.10 | 22.55 |
| Primates | KUPRI 1625 | Papio anubis | 29400 | M | 51.94 | 26.93 | 14.47 | 13.51 | 17.74 | 18.36 | 17.57 | 16.92 | 13.29 | 24.88 | 22.20 | 24.19 |
| Primates | KUPRI 307 | Papio anubis | 42000 | M | 50.28 | 25.68 | 14.29 | 13.70 | 16.94 | 19.48 | 14.56 | 16.76 | 12.10 | 23.67 | 20.42 | 23.74 |
| Primates | KUPRI 2779 | Papio hamadryas | 10200 | F | 37.31 | 18.75 | 9.68 | 8.41 | 12.63 | 12.54 | 13.09 | 12.82 | 9.53 | 16.78 | 13.97 | 16.66 |
| Primates | KUPRI 6077 (subadult) | Papio hamadryas | 18100 | M | 48.48 | 22.93 | 12.67 | 11.77 | 16.83 | 16.70 | 12.77 | 15.95 | 11.83 | 20.19 | 16.96 | 18.39 |
| Primates | KUPRI 6449 | Aotus trivirgatus | 940 | M | 20.52 | 8.64 | 4.13 | 3.95 | 5.20 | 6.31 | 6.69 | 5.82 | 4.85 | 8.39 | 7.53 | 8.49 |
| Primates | KUPRI 7125 | Aotus trivirgatus | 1064 | F | 19.39 | 8.29 | 4.20 | 3.84 | 5.12 | 5.16 | 6.12 | 5.65 | 4.36 | 8.35 | 7.51 | 8.48 |
| Primates | KUPRI 7130 | Callithrix jacchus | 300 | F | 11.45 | 5.49 | 2.56 | 2.94 | 3.27 | 3.16 | 3.71 | 3.74 | 2.56 | 4.90 | 4.12 | 4.46 |
| Primates | KUPRI 6424 | Callithrix jacchus | 382 | M | 11.23 | 4.46 | 2.29 | 2.35 | 3.06 | 3.48 | 3.36 | 3.12 | 2.53 | 4.73 | 4.01 | 4.09 |
| Primates | KUPRI 4487 | Callithrix jacchus | 460 | M | 10.66 | 4.17 | 2.44 | 2.27 | 2.56 | 2.49 | 3.76 | 3.14 | 2.32 | 4.43 | 3.75 | 4.22 |
| Primates | KUPRI 6091 | Cebus apella | 2600 | F | 24.55 | 10.37 | 5.18 | 4.54 | 6.90 | 7.32 | 7.31 | 7.09 | 5.64 | 8.80 | 7.85 | 8.55 |
| Primates | KUPRI 4245 | Cebus apella | 2200 | M | 26.64 | 12.77 | 5.98 | 6.19 | 7.82 | 8.24 | 8.94 | 8.89 | 5.92 | 10.56 | 8.81 | 10.41 |
| Primates | KUPRI 6429 | Saguinus midas | 400 | M | 13.78 | 5.67 | 3.15 | 2.56 | 3.18 | 3.32 | 4.52 | 3.94 | 2.82 | 5.45 | 4.35 | 4.71 |
| Primates | KUPRI 4314 | Saguinus midas | 550 | F | 13.60 | 5.96 | 3.05 | 2.39 | 3.41 | 3.95 | 4.46 | 4.09 | 2.71 | 5.22 | 4.12 | 4.68 |
| Primates | KUPRI 7174 | Saguinus oedipus | 450 | M | 14.56 | 6.04 | 2.60 | 2.63 | 3.34 | 4.16 | 4.39 | 4.32 | 2.88 | 5.42 | 4.42 | 4.74 |
| Primates | KUPRI 4282 | Saimiri sciureus | 700 | M | 16.53 | 7.10 | 3.16 | 3.40 | 4.56 | 4.27 | 5.09 | 5.09 | 3.66 | 6.66 | 5.66 | 5.79 |
| Primates | KUPRI 3908 | Saimiri sciureus | 540 | F | 17.14 | 7.07 | 3.20 | 3.20 | 3.99 | 4.84 | 5.78 | 5.22 | 3.92 | 6.83 | 5.92 | 6.64 |
| Primates | KUPRI 4691 | Galago crassicaudatus | 910 | M | 33.26 | 6.89 | 2.78 | 3.21 | 4.77 | 8.13 | 4.61 | 5.27 | 4.05 | 7.26 | 5.75 | 6.54 |
| Primates | KUPRI 4315 | Galago senegalensis | 100 | F | 25.99 | 4.30 | 1.77 | 2.05 | 2.87 | 4.44 | 2.90 | 2.44 | 2.15 | 4.37 | 3.78 | 4.18 |
| Primates | KUPRI 6699 | Lemur catta | 2330 | F | 24.09 | 10.22 | 3.86 | 5.25 | 6.56 | 6.97 | 6.41 | 7.40 | 5.02 | 9.79 | 8.79 | 9.28 |
| Scandentia | KUPRI 2789 | Tupaia glis | 125 | F | 9.20 | 4.71 | 1.73 | 1.78 | 2.39 | 3.18 | 2.69 | 2.62 | 1.94 | 3.52 | 2.71 | 2.99 |
| Scandentia | KUPRI 2914 | Tupaia glis | 90 | F | 8.73 | 4.20 | 1.49 | 1.57 | 2.20 | 2.96 | 2.53 | 2.61 | 1.91 | 3.27 | 2.56 | 2.90 |
| Carnivora | NSM-M 31458 | Ailuropoda melanoleuca | 108300 | F | 61.64 | 28.87 | 10.97 | 12.27 | 22.64 | 28.55 | 24.81 | 17.87 | 13.75 | 26.62 | 19.85 | 25.24 |
| Carnivora | KUPRI-Z 441 | Canis familiaris | 5690 | F | 33.29 | 13.20 | 6.24 | 5.73 | 9.89 | 14.34 | 11.03 | 10.52 | 7.69 | 14.16 | 9.42 | 10.71 |
| Carnivora | KUPRI-Z 438 | Canis familiaris | 14500 | M | 39.30 | 15.23 | 7.26 | 6.91 | 11.25 | 15.75 | 12.94 | 11.40 | 8.70 | 17.22 | 11.14 | 11.82 |
| Carnivora | KUPRI-Z 986 | Felis catus | 3300 | ? | 28.72 | 11.08 | 4.81 | 5.10 | 7.35 | 10.75 | 9.21 | 5.01 | 6.94 | 10.07 | 7.58 | 7.74 |
| Carnivora | KUPRI-Z 753 | Felis catus | 6160 | M | 30.50 | 11.82 | 4.69 | 5.79 | 7.89 | 13.08 | 9.34 | 5.29 | 7.14 | 10.82 | 8.80 | 8.03 |
| Carnivora | KUPRI-Z 462 | Martes melampus | 900 | ? | 18.55 | 9.29 | 3.94 | 3.02 | 5.23 | 6.07 | 8.10 | 4.38 | 4.93 | 7.09 | 5.76 | 6.18 |
| Carnivora | KUPRI-Z 460 | Martes melampus | 900 | F | 16.97 | 8.65 | 3.68 | 2.85 | 4.91 | 6.54 | 6.97 | 4.25 | 5.01 | 6.47 | 5.43 | 5.62 |
| Carnivora | KUPRI-Z 619 | Mustela sibirica | 300 | M | 9.90 | 4.52 | 1.99 | 1.64 | 2.74 | 3.31 | 3.66 | 2.68 | 2.60 | 3.98 | 3.39 | 4.03 |
| Carnivora | KUPRI-Z 620 | Mustela sibirica | 150 | F | 7.20 | 3.44 | 1.46 | 1.26 | 2.23 | 3.79 | 1.83 | 1.77 | 1.99 | 2.81 | 2.57 | 3.10 |
| Carnivora | KUPRI-Z 621 | Mustela sibirica | 420 | M | 12.48 | 6.19 | 3.18 | 2.59 | 4.24 | 3.96 | 5.14 | 3.58 | 2.88 | 5.47 | 5.02 | 5.83 |
| Carnivora | *** | Nyctereutes procyonoides | 3651 | M | 23.94 | 10.38 | 4.28 | 4.23 | 6.45 | 9.93 | 8.31 | 6.96 | 5.70 | 9.90 | 7.36 | 9.04 |
| Carnivora | **** | Nyctereutes procyonoides | 3805 | F | 23.77 | 10.39 | 4.20 | 4.21 | 6.35 | 9.80 | 8.38 | 7.02 | 5.81 | 9.80 | 7.35 | 8.77 |
| Carnivora | KUPRI-Z 985 | Paguma larvata | 3600 | F | 23.24 | 9.71 | 4.41 | 3.17 | 7.31 | 8.42 | 8.07 | 6.79 | 5.23 | 8.02 | 7.44 | 7.69 |
| Carnivora | KUPRI-Z 1376 (subadult) | Paguma larvata | 3400 | F | 24.92 | 10.08 | 4.05 | 3.44 | 6.42 | 9.08 | 8.67 | 7.14 | 5.60 | 8.29 | 7.82 | 8.33 |
| Carnivora | NMS-M 31999 (subadult) | Pantela leo | 131000 | M | 107.26 | 39.85 | 16.60 | 18.50 | 30.07 | 53.73 | 32.15 | 25.25 | 20.75 | 43.20 | 31.98 | 31.60 |
| Carnivora | NSM-M 33055 | Pantela leo | 97000 | F | 90.16 | 36.24 | 13.69 | 15.58 | 24.81 | 42.18 | 27.80 | 21.47 | 18.15 | 38.30 | 31.59 | 24.94 |
| Carnivora | KUPRI-Z 747 | Procyon lotor | 6100 | F | 28.93 | 12.94 | 4.89 | 5.07 | 8.26 | 9.21 | 10.43 | 7.70 | 7.64 | 11.08 | 8.98 | 10.39 |
| Carnivora | KUPRI-Z 767 | Suricata suricatta | 450 | ? | 15.05 | 6.01 | 2.64 | 2.53 | 4.43 | 4.97 | 4.67 | 3.13 | 3.88 | 5.59 | 4.78 | 5.21 |
| Carnivora | NSM-M 33061 | Ursus actos yesoensis | 163200 | F | 81.94 | 45.51 | 18.80 | 17.15 | 29.69 | 41.67 | 32.69 | 26.42 | 20.14 | 42.25 | 33.45 | 35.37 |
| Carnivora | KUPRI-Z 403 | Vulpes vulpes | 4700 | M | 31.90 | 12.77 | 5.11 | 5.31 | 8.09 | 14.00 | 10.16 | 9.29 | 7.83 | 12.78 | 9.27 | 9.86 |
| Carnivora | KUPRI-Z 414 | Vulpes vulpes | 3900 | F | 30.33 | 11.53 | 4.90 | 5.10 | 7.92 | 12.14 | 10.33 | 5.65 | 7.17 | 11.96 | 8.73 | 8.91 |
| Eulipotyphla | NSM-M 20690 | Urotrichus talpoides | 18 | F | 3.76 | 1.54 | 0.67 | 0.57 | 0.64 | 1.99 | 0.77 | 0.74 | 0.64 | 1.10 | 1.03 | 1.10 |
| Rodentia | NSM-M 35524 | Cavia porcellus | 550 | F | 13.09 | 5.86 | 2.54 | 2.54 | 2.75 | 3.17 | 3.60 | 2.88 | 2.30 | 4.25 | 2.68 | 3.54 |
| Lagomorpha | KUPRI-Z 785 | Lepus brachyurus | 3000 | M | 27.14 | 11.26 | 4.25 | 5.58 | 7.13 | 11.65 | 6.61 | 7.93 | 5.19 | 9.40 | 5.14 | 4.81 |
| Lagomorpha | KUPRI-Z 787 | Lepus brachyurus | 2150 | F | 27.42 | 11.54 | 4.41 | 5.12 | 6.62 | 12.30 | 7.17 | 7.92 | 3.67 | 10.07 | 5.39 | 5.58 |
| Tubulidentata | NSM-M 34334 | Orycteropus afer | 49300 | M | 75.68 | 35.62 | 14.81 | 14.28 | 17.15 | 42.19 | 19.36 | 32.68 | 19.40 | 34.93 | 25.97 | 26.89 |
| Artiodactyla | KUPRI-Z 1331 | Cervus nippon | 35000 | F | 66.33 | 22.43 | 6.34 | 6.92 | 14.28 | 43.11 | 14.88 | 16.77 | 8.59 | 23.49 | 16.04 | 18.60 |
| Artiodactyla | NSM-M 33056 | Bubalus babalis | 513000 | M | 156.59 | 54.88 | 21.18 | 25.78 | 42.72 | 101.09 | 32.02 | 15.40 | 43.02 | 67.44 | 39.14 | 49.37 |
| Artiodactyla | NSM-M 31301 | Bubalus babalis | 374400 | F | 143.39 | 50.07 | 19.05 | 22.27 | 39.17 | 89.82 | 30.50 | 16.31 | 42.88 | 57.71 | 34.69 | 43.03 |
| Artiodactyla | NSM-M 31304 | Giraffa camelopardalis | 800000 | M | 192.06 | 77.57 | 25.99 | 31.18 | 51.72 | 123.99 | 43.58 | 21.02 | 54.05 | 91.03 | 51.03 | 53.35 |
| Artiodactyla | NSM-M 33057 | Giraffa camelopardalis | 620000 | F | 184.80 | 73.78 | 22.86 | 32.17 | 53.33 | 114.17 | 45.16 | 23.41 | 56.42 | 87.53 | 49.61 | 59.00 |
| Artiodactyla | NSM-M 31318 | Oryx dammah | 99000 | M | 94.47 | 33.22 | 9.34 | 12.62 | 25.10 | 63.23 | 20.78 | 13.16 | 24.91 | 37.08 | 25.17 | 26.67 |
| Perissodactyla | NSM-M 33530 | Ceratotherium simum | 1400000 | F | 129.50 | 81.45 | 44.32 | 45.66 | 56.34 | 73.11 | 38.60 | 35.68 | 37.24 | 66.00 | 57.22 | 78.25 |
| Perissodactyla | NSM-M 31302 | Equus ferus przewalskii | 357400 | F | 101.59 | 53.38 | 23.64 | 18.33 | 30.72 | 57.16 | 22.75 | 13.23 | 21.54 | 46.36 | 40.65 | 49.98 |
| Perissodactyla | NSM-M 31303 | Equus ferus przewalskii | 345100 | F | 105.95 | 52.46 | 22.72 | 19.75 | 32.84 | 57.37 | 23.75 | 13.72 | 19.04 | 47.35 | 39.36 | 50.33 |
| Perissodactyla | NSM-M 33398 | Equus grevyi | 295400 | F | 107.14 | 53.66 | 22.66 | 19.06 | 30.83 | 56.87 | 18.68 | 12.24 | 19.02 | 47.51 | 43.01 | 50.98 |
| Perissodactyla | NSM-M 31634 | Tapirus indicus | 297900 | M | 111.49 | 51.99 | 22.95 | 15.99 | 29.07 | 59.96 | 24.36 | 17.59 | 30.53 | 41.50 | 32.91 | 40.44 |
| Perissodactyla | NSM-M 33067 | Tapirus terrestris | 237700 | F | 101.08 | 45.66 | 26.13 | 18.19 | 24.97 | 52.62 | 23.93 | 14.69 | 27.34 | 36.21 | 30.88 | 35.92 |
| Marsupialia | NSM-M 35838 | Macropus giganteus | 56200 | M | 80.03 | 25.64 | 11.72 | 17.44 | 23.08 | 57.15 | 14.91 | 25.07 | 22.93 | 28.96 | 23.92 | 26.31 |
Institutional abbreviations: KUPRI, Primate Research Institute, Kyoto University, Inuyama, Japan; KUPRI-Z, zoological collection stored in KUPRI; NSM-M, mammalian collection stored in National Museum of Nature and Science, Tsukuba, Japan (formerly National Science Museum, Tokyo, Japan). Other abbreviations: BM, body mass; M, male; F, female; CA1–CA12, linear measurements of the calcaneus used in this study (in mm; Figure 1; Table 2).

Twelve linear measurements (CA1–CA12) made on the calcaneus used in this study. The definitions of CA1–CA12 are shown in Table 2. The illustrations are based on a left calcaneus of Macaca fuscata (Primates, Catarrhini, Cercopithecidae): (A) dorsal (anterior) view; (B) lateral view; (C) distal view.
| Measurement | Definition |
|---|---|
| CA1 | calcaneal length (= C1 of Dagosto and Terranova, 1992) |
| CA2 | calcaneal width at the talar articular surfaces (~C2 of Dagosto and Terranova, 1992) |
| CA3 | width of the posterior talar articular surface (= C4 of Dagosto and Terranova, 1992) |
| CA4 | width of the posterior calcaneal body |
| CA5 | width of the tuberosity |
| CA6 | length of the posterior calcaneal body (= C7 of Dagosto and Terranova, 1992) |
| CA7 | length of the posterior talar articular surface (= C3 of Dagosto and Terranova, 1992) |
| CA8 | width of the articular surface for cuboid (= C6 of Dagosto and Terranova, 1992) |
| CA9 | height of the articular surface for cuboid (= C5 of Dagosto and Terranova, 1992) |
| CA10 | height at the posterior talar articular surface |
| CA11 | height at the posterior calcaneal body |
| CA12 | height at the tuberosity |
The data were transformed to natural logarithms for the analyses. The regression analyses were performed using the same procedure used by Tsubamoto (2014) and Tsubamoto et al. (2016). They were performed on two data sets: the land mammal model (superficially 69 individuals; including primates) and primate model (superficially 28 individuals) (Table 1, Table 3).
| ln CA1 | ln CA2 | ln CA3 | ln CA4 | ln CA5 | ln CA6 | ln CA7 | ln CA8 | ln CA9 | ln CA10 | ln CA11 | ln CA12 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Land mammal model | Slope | 2.969 | 2.928 | 2.857 | 2.815 | 2.747 | 2.359 | 3.074 | 3.027 | 2.757 | 2.779 | 2.861 | 2.777 |
| N = 69 | Intercept | −1.611 | 0.981 | 3.486 | 3.629 | 2.718 | 2.773 | 1.788 | 2.390 | 3.227 | 1.600 | 2.041 | 1.917 |
| t-value = 1.9960 | SEE | 0.649 | 0.387 | 0.639 | 0.609 | 0.441 | 0.674 | 0.660 | 1.064 | 0.571 | 0.441 | 0.522 | 0.551 |
| (df = 69 – 2 = 67) | adjusted R2 | 0.941 | 0.979 | 0.943 | 0.948 | 0.973 | 0.936 | 0.939 | 0.841 | 0.954 | 0.973 | 0.962 | 0.957 |
| (95% CL) | adjusted CF | 1.156 | 1.076 | 1.203 | 1.161 | 1.116 | 1.092 | 1.342 | 1.876 | 1.037 | 1.061 | 1.205 | 1.165 |
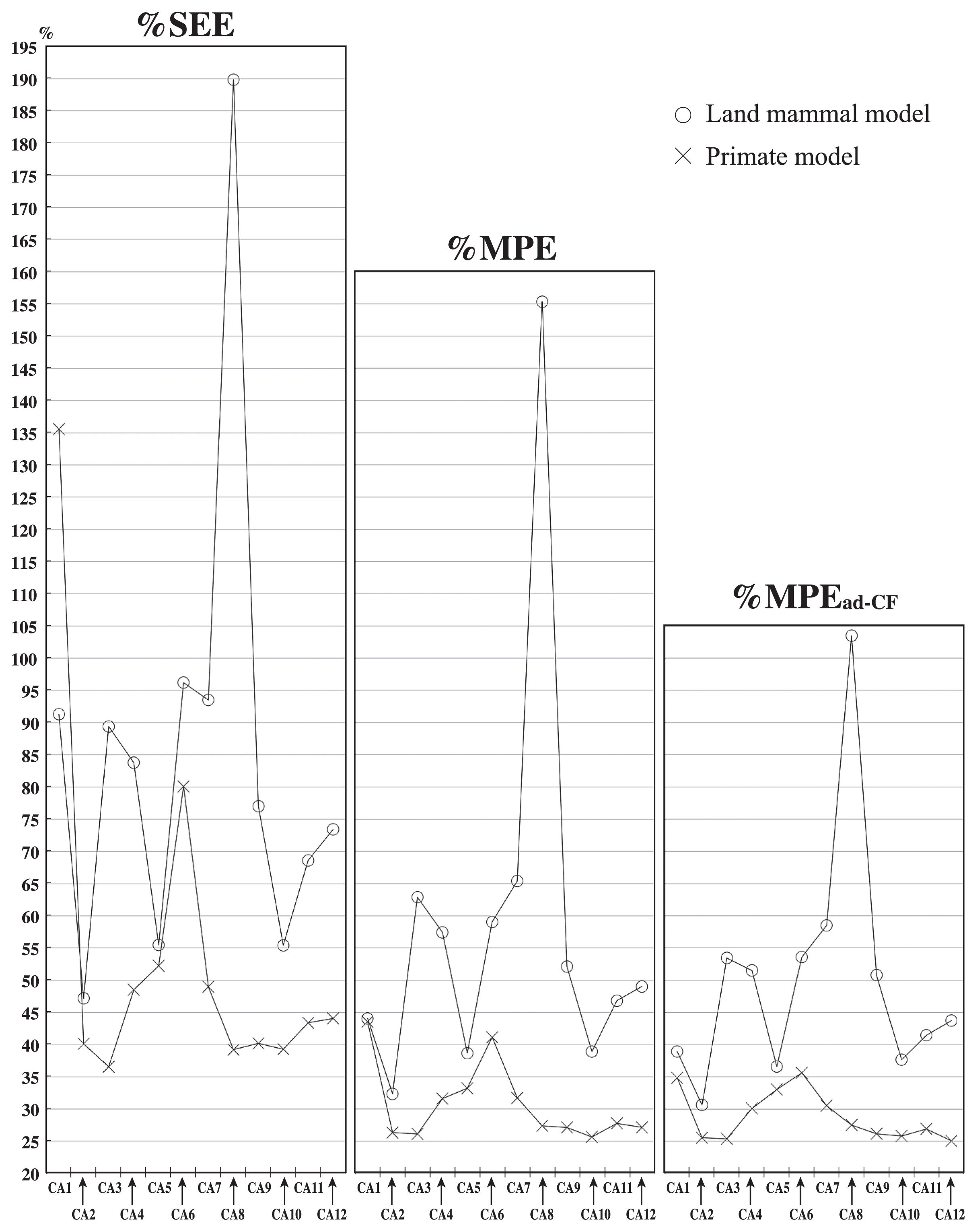
| %SEE | 91.293 | 47.210 | 89.396 | 83.779 | 55.458 | 96.225 | 93.515 | 189.815 | 77.019 | 55.415 | 68.611 | 73.418 | |
| %MPE | 44.075 | 32.360 | 62.890 | 57.423 | 38.673 | 59.048 | 65.407 | 155.350 | 52.109 | 38.924 | 46.841 | 49.036 | |
| %MPEad-CF | 38.925 | 30.649 | 53.452 | 51.509 | 36.592 | 53.591 | 58.515 | 103.496 | 50.793 | 37.654 | 41.462 | 43.755 | |
| Primate model | Slope | 3.153 | 2.782 | 2.555 | 2.678 | 2.656 | 2.617 | 2.957 | 2.836 | 2.891 | 2.938 | 2.871 | 2.690 |
| N = 28 | Intercept | −2.325 | 1.213 | 3.536 | 3.424 | 2.756 | 2.527 | 1.834 | 2.139 | 2.914 | 1.038 | 1.649 | 1.704 |
| t-value = 2.0555 | SEE | 0.857 | 0.337 | 0.312 | 0.396 | 0.420 | 0.588 | 0.399 | 0.331 | 0.338 | 0.331 | 0.361 | 0.365 |
| (df = 28 – 2 = 26) | adjusted R2 | 0.823 | 0.973 | 0.977 | 0.962 | 0.957 | 0.917 | 0.962 | 0.974 | 0.972 | 0.974 | 0.969 | 0.968 |
| (95% CL) | adjusted CF | 1.331 | 1.071 | 1.067 | 1.074 | 1.029 | 1.098 | 1.045 | 1.103 | 1.092 | 1.077 | 1.101 | 1.064 |
| %SEE | 135.574 | 40.135 | 36.565 | 48.535 | 52.244 | 80.102 | 48.991 | 39.223 | 40.210 | 39.299 | 43.408 | 44.079 | |
| %MPE | 43.558 | 26.349 | 26.139 | 31.626 | 33.218 | 41.161 | 31.717 | 27.412 | 27.144 | 25.677 | 27.780 | 27.157 | |
| %MPEad-CF | 34.847 | 25.552 | 25.390 | 30.094 | 33.046 | 35.604 | 30.527 | 27.520 | 26.167 | 25.828 | 26.928 | 25.059 |
Abbreviations: N, sample size; SEE, standard error of estimate; adjusted R2, coefficient of determination adjusted to the number of variables; adjusted CF, correction factor adjusted using the three correction factors proposed by Tsubamoto (2014); df, degrees of freedom; CL, confidence level; %SEE, percent standard error of estimate; %MPE, mean percentage prediction error; %MPEad-CF, %MPE for the corrected values using the adjusted CF (Tsubamoto, 2014). Bold value with underline indicates the lowest value in each row of %SEE, %MPE, and %MPEad-CF; bold value indicates the second lowest value in it; and value with underline indicates the third lowest value in it.
The results of the multiple regression analysis with a stepwise option performed using the JMP package (SAS Institute Inc.) vary according to the criteria used in the software. Therefore, here, simple linear bivariate regression analysis was applied to perform the body mass estimation. Although multiple regression analysis or areal/volumetric regression analysis may provide more ‘accurate’ equations, I chose simple linear regression analysis to provide simpler equations and to extend the availability and applicability of the equations for vertebrate paleontologists. On the other hand, although Model II regression techniques such as major axis and reduced major axis regressions are sometimes used for body mass estimation (e.g. Egi, 2001; Niskanen et al., 2018; Ruff and Niskanen, 2018; Ruff et al., 2018), I chose the least-squares regression because it can provide prediction errors (Warton et al., 2006).
When regression is performed using log-transformed data, a systematic detransformation bias is introduced (Smith, 1993a, b). To correct for this bias, correction factors are sometimes used (Sprugel, 1983; Snowdon, 1991; Smith, 1993a, b; Egi et al., 2002, 2004; Tsubamoto, 2014; Tsubamoto et al., 2016). Here, the adjusted correction factor (adjusted CF) proposed by Tsubamoto (2014) was calculated (Table 3) and applied. When estimating body mass, the estimated log value of the body mass is first de-transformed to the actual value in grams, and then is multiplied by the adjusted CF (Tsubamoto, 2014).
For the body mass estimation process, the 95% prediction intervals were calculated. The approximations of the 95% prediction intervals can be calculated as follows using the standard error of estimate (SEE) (Ruff, 2003; Tsubamoto, 2014; Tsubamoto et al., 2016): ± t-value × SEE. This approximation was used here to calculate the estimated body masses easily. The estimated body mass (BM in the equation) with 95% prediction interval for the measurements using the adjusted CF is calculated as follows: BM in g = {exp[slope × ln(measurement in mm) + intercept (± t -value × SEE )> × adjusted CF.
The degree of correlation (accuracy) between body mass and calcaneal size was evaluated using the percent standard error of estimate (%SEE) and the mean percentage prediction errors (%MPE and %MPEad-CF) (Tsubamoto, 2014; Tsubamoto et al., 2016). %SEE for natural log-transformed data was calculated as %SEE = (eSEE – 1) × 100 (Smith, 1984a; Egi et al., 2002; Ruff, 2003). %PE is the percentage of prediction error of the de-transformed value (not using the adjusted CF) and is calculated as %PE = (original value – estimated value)/estimated value × 100 (Smith, 1981, 1984a, b). %MPE is the arithmetic mean of the absolute values of %PE for each variable calculated for each individual (Smith, 1981, 1984a, b; Dagosto and Terranova, 1992). %MPEad-CF is %MPE for the values corrected using the adjusted CF (Tsubamoto, 2014).
The results of the simple bivariate regression analyses are shown in Table 3 and in Figure 2 and Figure 3. The values of the degree of correlation for the measurements (%SEE, %MPE, and %MPEad-CF) vary in each model.

Examples of body mass (BM; in g) estimate regressions and data scatters on a natural log scale: (A) using CA2 (in mm) for the mammal model; and (B) using CA3 (in mm) for the primate model. Black lines indicate least-squares axis. Dashed lines indicate the upper and lower 95% prediction limits.
In the land mammal model, CA2 has the lowest %SEE, %MPE, and %MPEad-CF values (Table 3; Figure 2), i.e. CA2 is the most suitable for the body mass estimation with land mammals as a target, among the 12 measurements. These %SEE, %MPE, and %MPEad-CF values (47.21, 32.36, and 30.65, respectively) of CA2 in the land mammal model are higher than those of the best measurement for the body mass estimation based on the talus of land mammals studied by Tsubamoto (2014) (41.98, 28.83, and 28.00, respectively). This implies that the talus is likely better than the calcaneus for body mass estimation with land mammals as a target.
In the primate model, CA3 has the lowest %SEE value and the second lowest %MPE and %MPEad-CF values (Table 3; Figure 2). Based on the %MPE and %MPEad-CF values of the primate model, CA2 and CA8–CA12 are as low as CA3 (Table 3; Figure 2). Therefore, CA3 appears to be the most suitable for the body mass estimation with the primates as a target, although CA2 and CA8–CA12 are roughly as suitable as CA3. Compared to the values of the good measurements for the body mass estimation in the talus of the primates studied by Tsubamoto et al. (2016), the %SEE value of CA3 of the primate model is lower than that studied by Tsubamoto et al. (2016); and its %MPE and %MPEad-CF values are nearly as low as that studied by Tsubamoto et al. (2016). This suggests that, unlike the land mammal model, the calcaneus appears to be as good as the talus for body mass estimation with the primates as a target. However, we should note the fact that the original data of these studies are in fact limited, so that the additional data may alter the results.
The examples of the regression equation with a 95% prediction interval to estimate the body mass (BM, g) are as follows. Using CA2 (in mm) for the land mammal model, BM = exp(2.928 × ln CA2 + 0.981 ± 0.772) × 1.076. Using CA3 (in mm) for the primate model, BM = exp(2.555 × ln CA3 + 3.536 ± 0.641) × 1.067.
The results of the regression analyses of the primate model for CA1–CA3 and CA6–CA9 are briefly compared with those of the calcaneus of the all-strepsirrhine model for the linear measurements C1–C7 by Dagosto and Terranova (1992) (Table 4). The results of CA2–CA3 and CA6–CA9 are roughly similar to those by Dagosto and Terranova (1992). However, the results of CA1 (calcaneal length) are distinguished from those by Dagosto and Terranova (1992): in particular, the slope and intercept are quite different with each other (Table 4). This difference can be information in considering the ecological or phyletic characteristics of the calcaneus of the strepsirrhines among the primates.
| This study primate model | Dagosto and Terranova (1992) | This study primate model | Dagosto and Terranova (1992) | This study primate model | Dagosto and Terranova (1992) | This study primate model | Dagosto and Terranova (1992) | |
|---|---|---|---|---|---|---|---|---|
| ln CA1 | ln C1 | ln CA2 | ln C2 | ln CA3 | ln C4 | ln CA6 | ln C7 | |
| Slope | 3.15 | 1.48 | 2.78 | 2.86 | 2.55 | 3.34 | 2.62 | 2.48 |
| Intercept | –2.33 | 2.13 | 1.21 | 1.42 | 3.54 | 3.23 | 2.53 | 2.70 |
| SEE | 0.86 | 1.16 | 0.34 | 0.34 | 0.31 | 0.38 | 0.59 | 0.70 |
| r | 0.95 | 0.48 | 0.95 | 0.97 | 0.95 | 0.96 | 0.95 | 0.85 |
| %SEE | 135.57 | 218.99 | 40.14 | 40.49 | 36.56 | 46.23 | 80.10 | 101.38 |
| %MPE | 43.56 | 59.32 | 26.35 | 26.13 | 26.14 | 29.57 | 41.16 | 46.36 |
| ln CA7 | ln C3 | ln CA8 | ln C6 | ln CA9 | ln C5 | |
|---|---|---|---|---|---|---|
| Slope | 2.96 | 2.74 | 2.84 | 2.70 | 2.89 | 2.93 |
| Intercept | 1.83 | 2.35 | 2.14 | 2.46 | 2.91 | 2.81 |
| SEE | 0.40 | 0.36 | 0.33 | 0.28 | 0.34 | 0.44 |
| r | 0.95 | 0.96 | 0.95 | 0.98 | 0.95 | 0.94 |
| %SEE | 48.99 | 43.33 | 39.22 | 32.31 | 40.21 | 55.27 |
| %MPE | 31.72 | 28.44 | 27.41 | 21.05 | 27.14 | 32.84 |
Abbreviations: SEE, standard error of estimate; r, Pearson’s product moment correlation coefficient; %SEE, percent standard error of estimate; %MPE, mean percentage prediction error. CA1: C1 of Dagosto and Terranova (1992); CA2: approximately equal to C2 of Dagosto and Terranova (1992); CA3: C4 of Dagosto and Terranova (1992); CA6: C7 of Dagosto and Terranova (1992); CA7: C3 of Dagosto and Terranova (1992); CA8: C6 of Dagosto and Terranova (1992); CA9: C5 of Dagosto and Terranova (1992).
I am grateful to the following individuals involved in access to the specimens used in this research: Masanaru Takai, Takeshi Nishimura, and Naoko Egi (Primate Research Institute, Kyoto University, Inuyama, Japan); and Shin-ichiro Kawada (National Museum of Nature and Science, Tsukuba, Japan). I am also grateful to the reviewer for the useful comments. This work was supported by the Cooperation Program (2011-A-3, 2012-B-2, 2013-B-15, and 2014-B-2) of the Primate Research Institute (Kyoto University, Inuyama, Japan) and by JSPS KAKENHI grant numbers 21770265, 25840172, and 16K07534.