Abstract
Domesticated camelids spread to Peru’s Northern Highlands by 500 BC. The complexity and development of the society that then developed in the region have been explained by social networks enabled by the use of the llama as a cargo animal. However, the actual use of domesticated camelids in the Formative Period remains unclear. This study analyzed camelid skeletal remains excavated from the Pacopampa site (1200–400 BC) to provide information on the actual use of these animals. Osteometry determined the camelid to be llama; alpaca was not identified in the samples. Body part frequency, butchering mark distribution, and mortality profile revealed ritual consumption of the meaty part of young individuals. Four sacrificed immature llamas were detected. In contrast to the great similarity with the consumption process of artiodactyls’ only llamas were sacrificed, while deer were not targeted. Although there were no features in the animal bone material indicating the use of secondary products, the increasing number of artifacts related to textile production suggest the fiber might be the result of llama herding.
Introduction
The domestication of the South American camelid and the influence of this process on prehistoric society have been a central issue in Andean zooarchaeology. Although it is accepted that the llama (Lama glama) was domesticated from the guanaco (Lama guanicoe) and the alpaca (Vicugna pacos) from the vicuña (Vicugna vicugna), there is an absence of general agreement on the location and date of this process—an age of 2000 BC has been claimed for the Central Andes, and 2500–2000 BC for the South–Central Andes (Goñalons and Yacobaccio, 2006; Wheeler, 2012). The subsequent spread of domesticated forms varies from region to region. However, in the Northern Highlands of Peru, the domesticated camelid arrived around 500 BC (Shimada, 1985; Uzawa, 2008, 2019). In this region, the introduction of the domesticated camelid coincided with the development of social networks, and so the contribution of the llama as a pack animal was valued (Miller and Burger, 1995).
Modern pastoralists focus on the secondary products of camelids. In the highlands, both animals are grown—alpacas for their fiber, while llamas are taken to the valleys to be used as pack animals during crop harvest (Bonavia, 2009). This resource development using altitude differences is known as vertical control (Murra, 1972). Although the agropastoral complex using camelids has been recognized as the key to the development of ancient civilizations, there is little evidence that this system existed in c. 500 BC in the wider range of the Central Andes.
The ancient Andean civilization developed across a wide geography. Its ceremonial centers were established in the coastal deserts, along the rivers, and in the mountain valleys after 3000 BC (Burger, 1992; Haas and Creamer, 2006). Societies in each region developed using local resources acquired by hunting, gathering, fishing, and farming (Onuki, 1985; Chicoine and Rojas, 2013; Pozorski and Pozorski, 2008). Recent research has shown that the appearance of the ceremonial center on the coast preceded that of the mountains (Shady, 2014). Therefore, the role of camelid herding in the highlands during the early stages of civilization can hardly be overestimated. To explain the simultaneous dispersal of camelid herding and social development after 500 BC, we need to better understand, through archaeological data, animal use in the Formative Period.
We conducted zooarchaeological research at two ceremonial center sites in the Northern Highlands of Peru, namely Pacopampa and Kuntur Wasi, and showed that the domesticated camelid emerged in the late Formative Period at both sites, when the society of the region became remarkably complex (Burger, 1992; Uzawa, 2008, 2019; Seki, 2014). It is unclear how camelid use was linked to social complexity. The main argument is that the Formative societies in the region were supported by rich agricultural production and stable deer hunting even after the introduction of the domestic animal, making it inappropriate to apply the model that emphasizes a shift to a mixed agropastoral economy with vertical control. The widespread dispersal of domesticated camelids during the late Formative Period was undoubtedly an important event that resulted in the systematic livestock management later observed at Wari and Inca. More concrete archaeological evidence is needed to clarify the role that these animals played in social development.
Formative Period societies are recognized as small groups linked together by shared rituals in the ceremonial center (Burger, 1992). Nevertheless, activities in the ceremonial center were diverse, including the construction of cemeteries for ancestor worship and the production of bone tools, copper products, and textiles (Seki et al., 2019). The contribution of camelids needs to be examined in relation to the ritual activities at the site. For this purpose, clarifying the use of camelids in the ceremonial center, in contrast to the use of other artiodactyls, will provide basic data for assessing the effect of camelid introduction on society.
Materials and Methods
Pacopampa Site
The Pacopampa site is at 2500 masl on the eastern slope of the Andean mountains in the Northern Highlands of Peru, Department of Cajamarca (Figure 1). In this region, the mountains gradually decrease their height toward the border with Ecuador and open pathways to the Pacific coast and the Amazonian forest. The site is situated in the connecting area of the quechua zone and the warmer temple zone, one of the geographical regions of Peru, characterized as cool but farmable, and a warmer area suited for tropical crops such as manioc and cotton, respectively (Pulgar, 1981). Modern villagers grow soybeans, maize, and other root vegetables around the site.
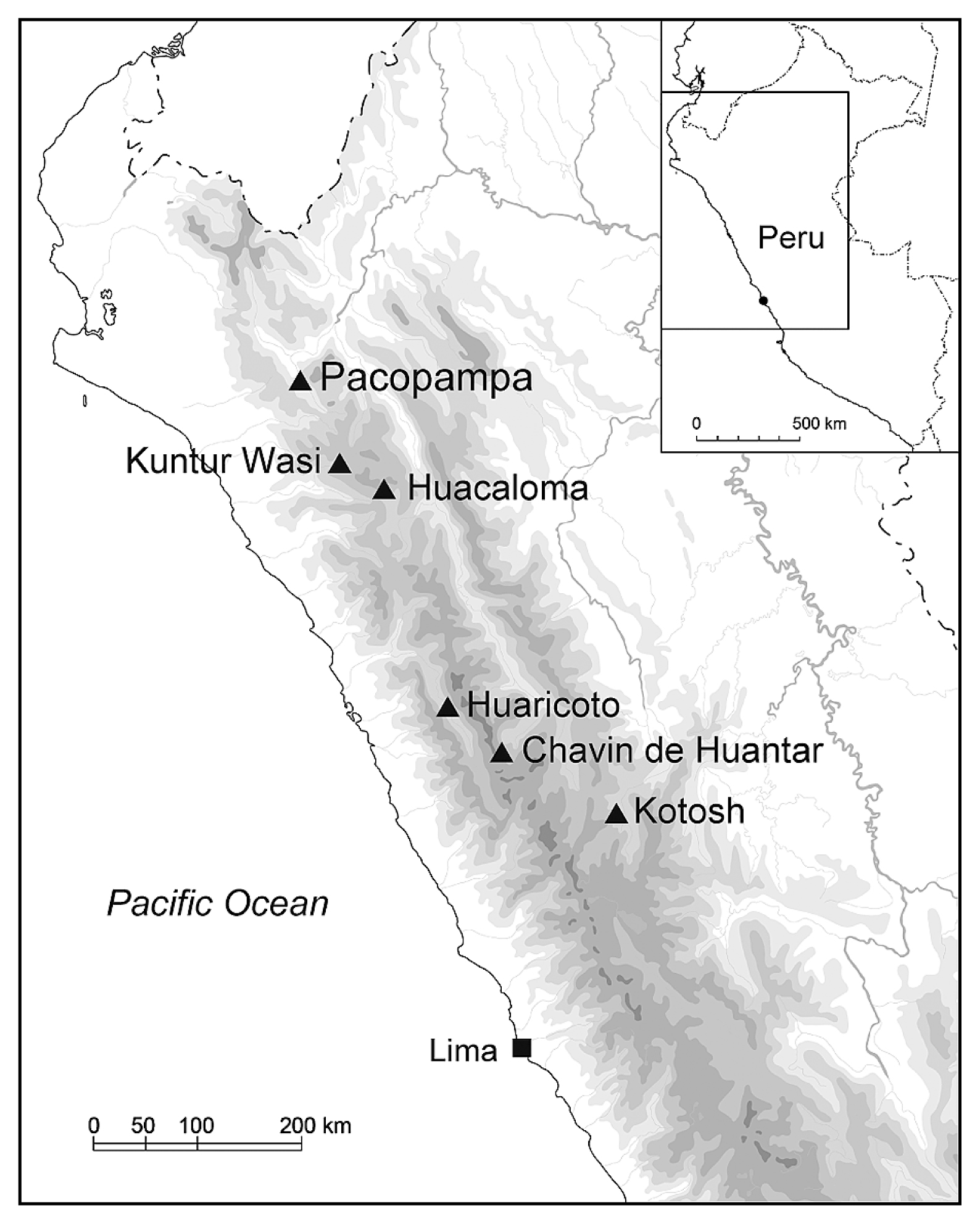
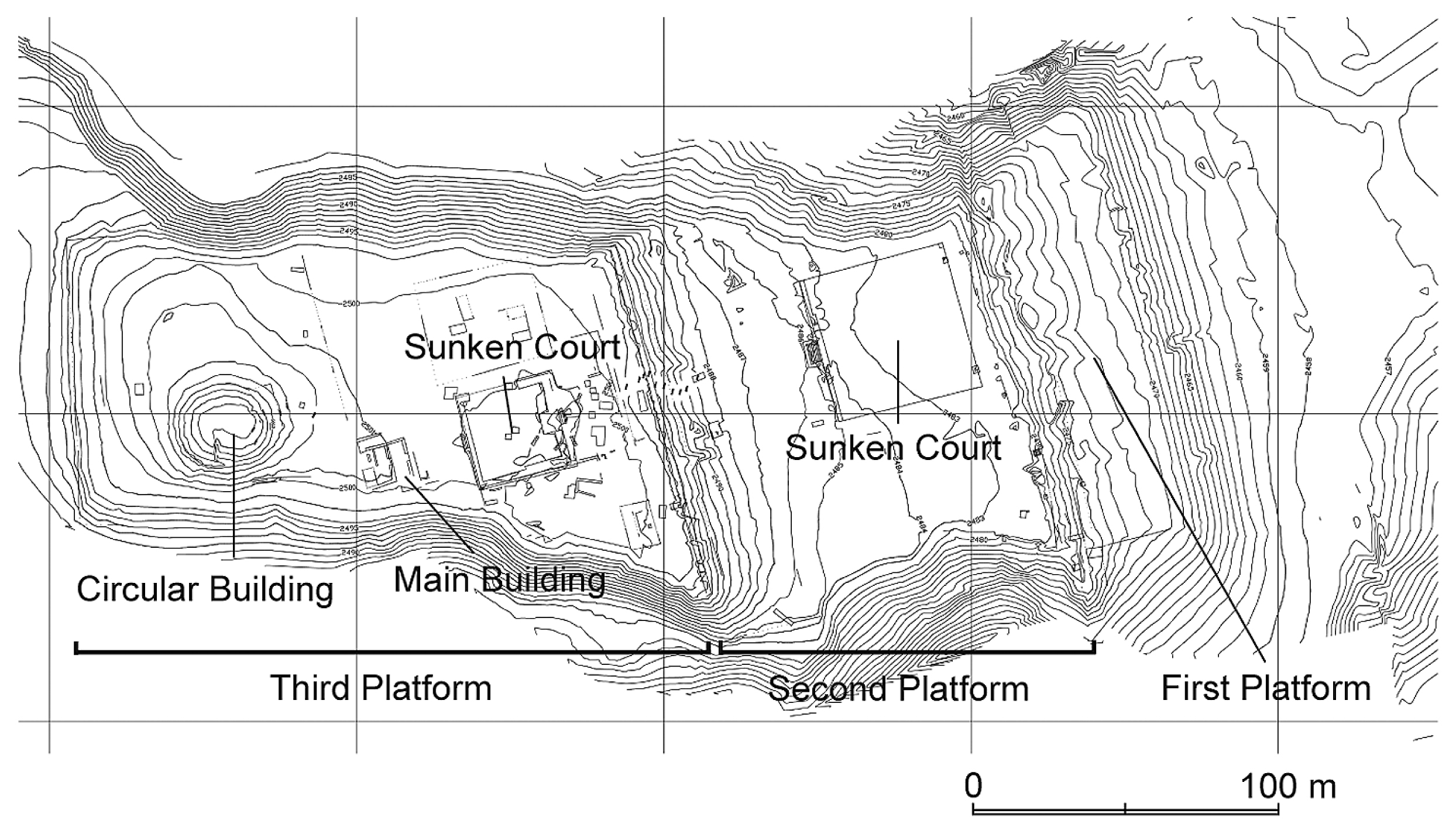
Pacopampa is a large ceremonial center with a total area of 4 ha; it consists of three platforms. Its major architectural features are located on the third platform, which is the uppermost part of the site. After several surveys by different archaeologists (Rosas La Noire and Shady, 1970; Fung, 1975; Morales, 1980), the Pacopampa Archaeological Project, directed by Yuji Seki as a joint project of National Museum of Ethnology in Japan and Universitad de National major de San Marcos in Peru, began excavating the site in 2005. This project has helped illuminate the construction process of the architectures and established the existence of two sequential phases Pacopampa I (1200 cal BC–700 cal BC) and Pacopampa II (700 cal BC–400 cal BC) (Seki, 2014, Seki et al., 2019). In the Andean archaeological chronology, Pacopampa I corresponds to the Middle Formative Period and Pacopampa II to the late Formative Period. Although each phase is subdivided into subphases, namely IA and IB, and IIA and IIB, respectively, the 14C dates for the subphases are not yet available. Therefore, in this study, samples were compiled and analyzed in two time periods, Pacopampa I and II.
Bone assemblage
The material we analyzed comprises mammalian bone remains recovered from the excavation of the third platform, the core of the Pacopampa site (Figure 2). The third platform is about 100 m wide and at a depth of 200 m, where the construction structures are concentrated. The excavation was conducted based on a grid system divided into east–west and north–south squares of 20 m on each side. The material was collected by careful hand-picking. No dry or wet sieving was carried out due to the wet soil condition and the location constraints of the site. Approximately 20000 pieces of animal bone remains recovered from ten grids covering the central part of the third platform were identified to anatomical element and taxonomic affiliation up to the present. The bone remains were found on the floor surface and also mixed in the cover soil. For camelids, there were also buried individuals. The number of identified specimens (NISP) was 6246, with a high proportion of artiodactyls throughout Pacopampa I and II phases, and 60% of the material, or 80% if we exclude humans, being occupied by camelids and white-tailed deer (Odocoileus virginianus). Regardless of the provenance, the materials were compiled and quantitatively evaluated by period.
Methods
Osteometry
The bone morphology of the four South American camelid species is very similar, and there are no diagnostic features for each species except for the incisor teeth morphology, which is unique to the alpaca and vicuña. Hence, the differences in body size among them are important for the identification of taxa in fragmented archaeological materials (e.g. Izeta et al., 2009). In this study, we performed discriminant analysis using the maximum width (BFp) and maximum depth (DFp) of the proximal facet of proximal phalanges, which are the most commonly used measures to classify South American camelids. Measurements were made according to the method of Muños and Mondini (2014). Data on modern llamas and alpacas were taken from the literature (Kent, 1982; Cartajena, 2009; Izeta et al., 2009; Gasco and Marsh, 2015). These data include comparative specimens collected in Peru, Bolivia, Chile, and Argentina. Using the measurements of these modern camelids, discriminant analysis of the Pacopampa samples were conducted to determine the taxonomic attribute.
Body part frequency
We compare the relative frequency of the camelid skeletal elements with those of deer to confirm the selective transport of the body. Some fragments of vertebrae and ribs were not distinguishable between the two animal species and were identified as coming from artiodactyls, and thus excluded from the analyses. Since the preservation bias may lead to the differential occurrence of skeletal parts during the deposition processes, the bone structural density (BMD) of the skeletal elements and their frequency of occurrence were correlated to evaluate the effect of deficiencies caused by physicochemical factors. Each element’s frequency was calculated using %MNI (where MNI refers to minimum number of individuals), which indicates how many individuals of each bone element were found in the material relative to the maximum MNI calculated in the material. The BMD value for camelids was taken from Stahl (1999) and that for deer from Lyman (1984). When one element showed multiple BMD values, the more significant value was used.
Butchering mark
We observed the butchering marks to determine the consumption and use process of camelids brought to the site. This study took butchering marks in a broad sense and recorded seven types of them. Cut marks, chop marks, and peeling marks indicate dismembering and filleting; the percussion pit and striae reflect bone-cracking and marrow extraction; scrape marks signify skinning or periosteal removal. The frequency of each mark was calculated for each body part. The identification of each mark was made following White (1992).
Mortality profile
The age of the animals’ death reflects the purpose of livestock management (Greenfield et al., 1988; Yacobaccio, 2007). Camelid age estimation required a skeletal fusion schedule to calculate the camelid’s survivor curve at the Pacopampa site. There are several methods pertaining to the skeletal fusion sequences in camelids (Kent, 1982; Wing, 1972), and this study followed the system of Miller (2003). Miller reported the camelid mortality profile at Chavín de Huántar, a representative ceremonial center in the Central Andes. Using the same method enables us to compare the Pacopampa result with those from Chavín de Huántar.
The skeletal fusion process in camelids is normally complete in 5 years, even in the most slowly fusing elements. On the other hand, the life span of camelids ranges from 8 to 10 years (Bonavia, 2009). Age assessment based on bone fusion only covers the first half of the camelid life span. Thus, we also conducted an age assessment based on tooth eruption and wear to determine the life stage of older animals. The mortality profile of deer from Pacopampa II phase was also estimated to compare their use pattern to that of the camelids. Considering the animals’ different maximum life spans, the three age classes were used following Stiner (1994).
A juvenile, defined by the presence of deciduous teeth in the dentition or by permanent teeth in the process of eruption but not worn out, is an individual before reproductive maturity. Prime adulthood, marked by the replacement of the dp4 with the P4, corresponds to the stage wherein females are sexually mature and able to give birth, and with an increased ability to store fat. The transition from prime adult to old was determined by tooth wear patterns and was bounded by 61–65% of the maximum life span. There is a decrease in the reproduction rate in old adults, and fat accumulation is also lower than in their prime. In camelids, the boundary was set at 6 years of age in this study. The three age categories reflect the growth process of most artiodactyls, although each duration varies depending on the animal species and its life span (Stiner, 1994).
Buried camelid
Four camelid burial remains had been detected in the excavation up until 2019. Particular attention was given to the age, body part frequency, and butchering practices.
Data analysis and statistics
Spearman’s rank correlation coefficiency analysis was conducted without assuming a normal distribution between the skeletal parts’ frequency of occurrence and the density of the bone structure. Tests for independence were carried out for the occurrence of skeletal elements in deer and camelids using Fisher’s exact test. All analyses were performed with R version 4.0.3. P values are indicated in the text, and <0.05 was considered a statistically significant difference.
Results
Overview of the material and species composition
The ratio of camelids to deer changed between the Pacopampa I and the Pacopampa II phases. As we reported in an earlier study, camelids were almost absent in Pacopampa I and increased in Pacopampa II (Uzawa, 2008). In this study, we reanalyzed the data with additional materials from the excavation, conducted up to 2019, and confirmed the same tendency. Camelid herding was introduced in the Pacopampa II period (Table 1).
Table 1
Mammal remains from the Pacopampa site: camelids, which were almost non-existent in the Pacopampa I period, rapidly increased in the Pacopampa II period
| Phase |
Pacopampa I |
Pacopampa II |
| Taxa |
NISP |
%NISP |
MNI |
NISP |
% NISP |
MNI |
| Odocoileus virginianus |
970 |
59.9 |
34 |
1707 |
36.9 |
44 |
| Camelidae |
22 |
1.4 |
4 |
1331 |
28.8 |
25 |
| Artiodactyla indt. |
114 |
7.0 |
— |
639 |
13.8 |
— |
| Homo sapiens |
304 |
18.8 |
9 |
308 |
6.7 |
5 |
| Cavia porcellus |
139 |
8.6 |
17 |
461 |
10.0 |
56 |
| Sylvilagus brasiliensis |
53 |
3.3 |
8 |
78 |
1.7 |
11 |
| Canis familiaris |
7 |
0.4 |
1 |
76 |
1.6 |
4 |
| Aguti paca |
0 |
0.0 |
0 |
2 |
0.0 |
2 |
| Caviomorph |
3 |
0.2 |
— |
6 |
0.1 |
— |
| Lagidium peruanum |
4 |
0.2 |
2 |
1 |
0.0 |
1 |
| Rodencia |
3 |
0.2 |
— |
12 |
0.3 |
— |
| Didelphidae |
0 |
0.0 |
0 |
1 |
0.0 |
1 |
| Cebus sp. |
0 |
0.0 |
0 |
3 |
0.1 |
1 |
| Muridae |
0 |
0.0 |
0 |
2 |
0.0 |
1 |
| Total |
1619 |
|
|
4627 |
|
|
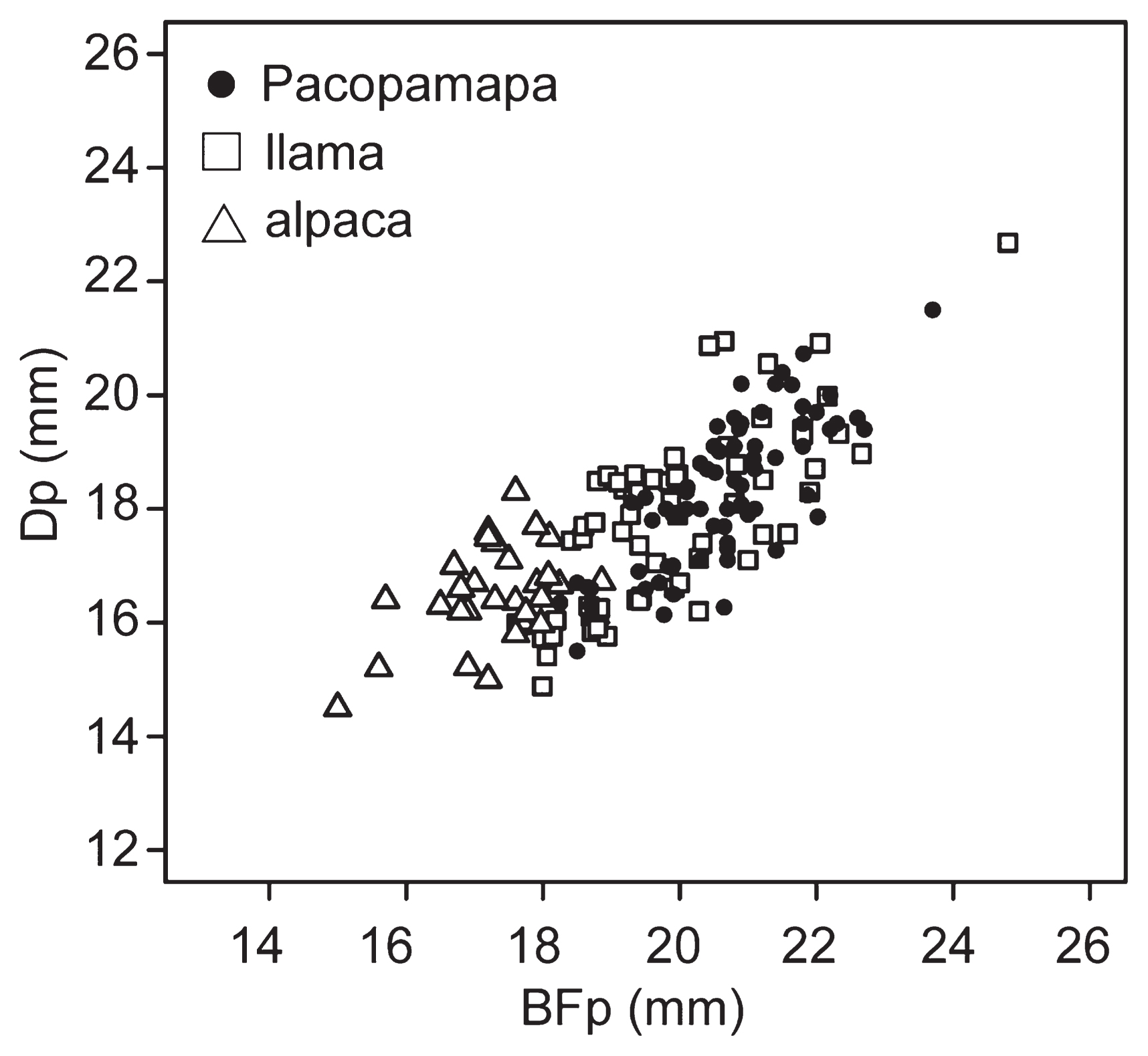
A total of 69 camelid proximal phalanges from Pacopampa were measured. BFp ranged between 18.3 and 23.7 mm, with a mean of 20.6 mm, and a standard deviation of 1.09 mm; DFp ranged between 15.5 and 21.5 mm, with a mean of 18.3 mm and a standard deviation of 1.28 mm (Figure 3). Most samples showed sizes corresponding to llamas, and some of the values overlapped with those of larger individuals of the alpaca herd. Discriminant analysis showed that all the materials from the Pacopampa site were judged as coming from llama. We performed the Shapiro–Wilk normality test and found that the measurements were normally distributed (BFp: W = 0.986, P = 0.637, DFp: W = 0.985, P = 0.600). This result is consistent with the result of discriminant analysis, where all samples consisted of a single taxon.
Skeletal frequency of camelids
An overview of the frequency of body parts of camelids and deer is shown in Figure 4, with details in Table 2. In both species, whole body parts were present, but there was a wide variation in the number of parts recovered for each element. Large bones of the proximal limb elements such as the humerus and femur were more frequent, and the head, spine, and distal limbs were less frequent. The number of vertebrae and ribs may have been underestimated because we excluded the fragments that could not be identified at species level from the analysis.
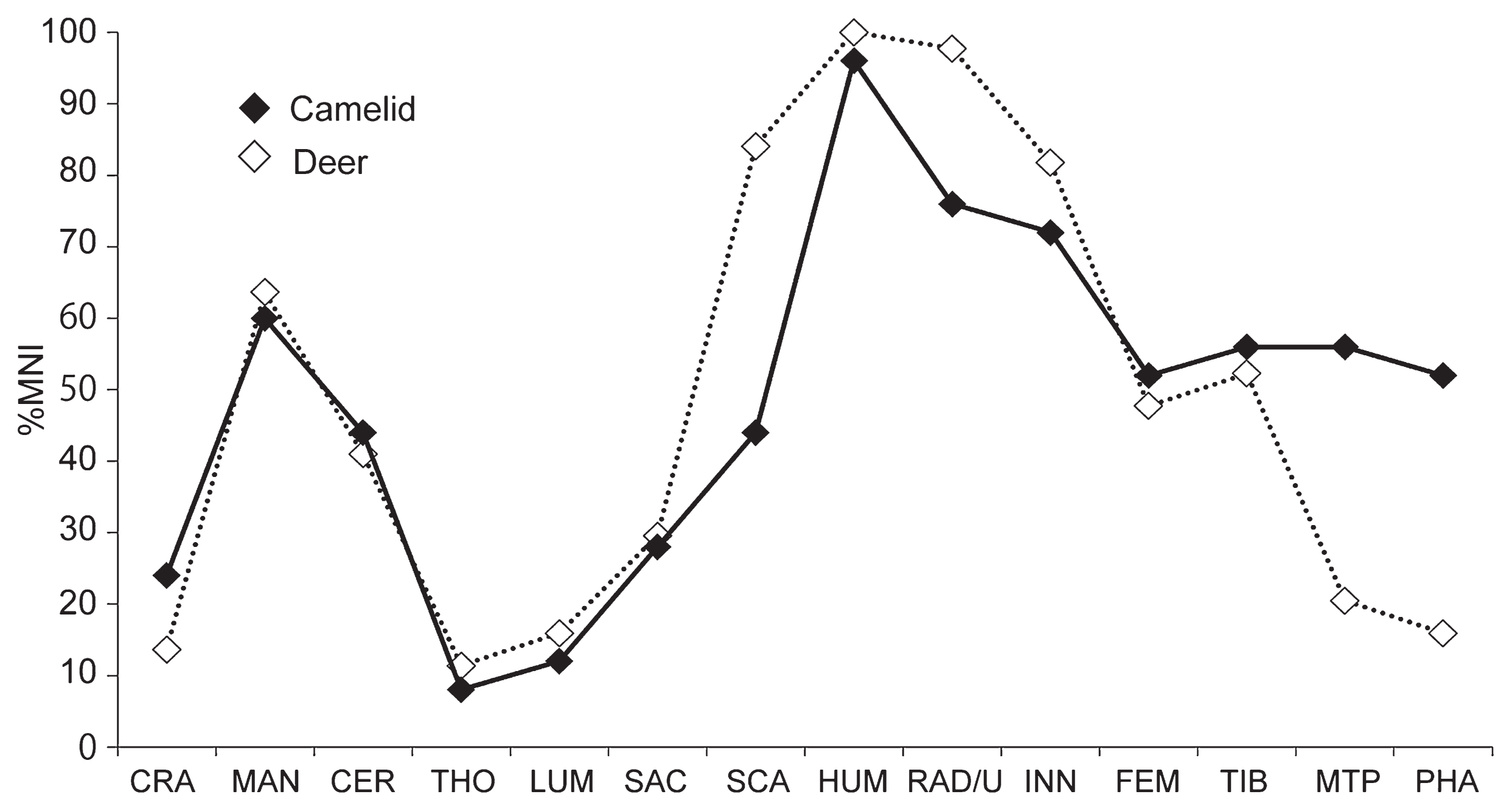
Table 2
Skeletal frequencies of camelid and deer remains. The skeletal frequencies of camelids and deer are shown by period, based on NISP and MNI. In the camelid, the radius and ulna are joined as one bone, whereas in the deer, they are separated. For comparison, the radius and ulna are shown as a combined in part (a), and the breakdown value of each element is shown in part (b)
| (a) Camelid and deer |
| Phase |
Camelid |
Deer |
| Pacopampa I |
Pacopampa II |
Pacopampa I |
Pacopampa II |
| Skeletal part |
NISP |
MNI |
%MNI |
NISP |
MNI |
%MNI |
NISP |
MNI |
%MNI |
NISP |
MNI |
%MNI |
| Crania |
2 |
1 |
33.3 |
23 |
6 |
24.0 |
42 |
5 |
14.7 |
51 |
6 |
13.6 |
| Mandible |
0 |
0 |
0.0 |
40 |
15 |
60.0 |
38 |
15 |
44.1 |
67 |
28 |
63.6 |
| Teeth |
1 |
1 |
33.3 |
54 |
3 |
12.0 |
25 |
2 |
5.9 |
116 |
6 |
13.6 |
| Atlas |
1 |
1 |
33.3 |
7 |
3 |
12.0 |
18 |
17 |
50.0 |
17 |
14 |
31.8 |
| Axis |
0 |
0 |
0.0 |
11 |
11 |
44.0 |
12 |
12 |
35.3 |
22 |
18 |
40.9 |
| Cervical vertebrae |
1 |
1 |
33.3 |
45 |
5 |
20.0 |
51 |
9 |
26.5 |
55 |
10 |
22.7 |
| Thoracic vertebrae |
0 |
0 |
0.0 |
24 |
2 |
8.0 |
56 |
3 |
8.8 |
70 |
5 |
11.4 |
| Lumber vertbrae |
1 |
1 |
33.3 |
19 |
3 |
12.0 |
68 |
10 |
29.4 |
49 |
7 |
15.9 |
| Sacrum |
0 |
0 |
0.0 |
9 |
7 |
28.0 |
13 |
10 |
29.4 |
16 |
13 |
29.5 |
| Ribs |
2 |
1 |
33.3 |
59 |
3 |
12.0 |
54 |
3 |
8.8 |
58 |
3 |
6.8 |
| Scapula |
0 |
0 |
0.0 |
28 |
11 |
44.0 |
48 |
23 |
67.6 |
68 |
37 |
84.1 |
| Humerus |
5 |
3 |
100.0 |
81 |
24 |
96.0 |
114 |
34 |
100.0 |
133 |
44 |
100.0 |
| Radius–ulna |
0 |
0 |
0.0 |
108 |
19 |
76.0 |
101 |
24 |
70.6 |
196 |
43 |
97.7 |
| Metacarpal |
1 |
1 |
33.3 |
36 |
14 |
56.0 |
0 |
0 |
0.0 |
23 |
9 |
20.5 |
| Innominate |
0 |
0 |
0.0 |
60 |
18 |
72.0 |
46 |
15 |
44.1 |
140 |
36 |
81.8 |
| Femur |
1 |
1 |
33.3 |
73 |
13 |
52.0 |
115 |
20 |
58.8 |
143 |
21 |
47.7 |
| Tibia |
2 |
1 |
33.3 |
74 |
14 |
56.0 |
69 |
12 |
35.3 |
112 |
23 |
52.3 |
| Astragalus |
0 |
0 |
0.0 |
23 |
13 |
52.0 |
17 |
11 |
32.4 |
33 |
19 |
43.2 |
| Calcaneum |
0 |
0 |
0.0 |
54 |
25 |
100.0 |
11 |
6 |
17.6 |
50 |
24 |
54.5 |
| Metatarsal |
0 |
0 |
0.0 |
38 |
14 |
56.0 |
8 |
4 |
11.8 |
23 |
6 |
13.6 |
| Metapodial |
2 |
1 |
33.3 |
70 |
8 |
32.0 |
8 |
|
0.0 |
23 |
|
0.0 |
| Carpal |
1 |
1 |
33.3 |
75 |
8 |
32.0 |
3 |
1 |
2.9 |
21 |
3 |
6.8 |
| Tarsal |
1 |
1 |
33.3 |
60 |
12 |
48.0 |
1 |
1 |
2.9 |
11 |
5 |
11.4 |
| First phalanx |
1 |
1 |
33.3 |
135 |
13 |
52.0 |
8 |
1 |
2.9 |
74 |
7 |
15.9 |
| Second pharanx |
0 |
0 |
0.0 |
63 |
8 |
32.0 |
5 |
1 |
2.9 |
33 |
5 |
11.4 |
| Third phalanx |
0 |
0 |
0.0 |
9 |
2 |
8.0 |
2 |
1 |
2.9 |
12 |
2 |
4.5 |
| (b) deer |
| Phase |
Pacopampa I |
Pacopampa II |
| Skeletal part |
NISP |
MNI |
%MNI |
NISP |
MNI |
%MNI |
| Antler |
9 |
2 |
5.9 |
13 |
2 |
4.5 |
| Radius |
65 |
24 |
70.6 |
128 |
38 |
86.4 |
| Ulna |
36 |
18 |
52.9 |
68 |
43 |
97.7 |
Spearman’s rank correlation coefficient analysis was used to check the relationship between each body part and BMD frequency showed no correlation (camelids: ρ = 0.247, P = 0.415, cervids: ρ = 0.412, P = 0.163). The uneven distribution of body parts cannot be explained by taphonomic bias alone, wherein robust bones are well preserved during the formation of the bone samples. It was suggested that the bias in body parts reflected cultural selection, such as selective delivery of body parts to the site and differences in the way body parts were consumed.
To investigate whether there exists a difference in the selectivity of body parts between camelids and deer, we compared the two species’ skeletal element frequencies. The skeletons were grouped into four parts, namely head, axial, proximal limbs, and distal limbs, and their frequencies were calculated based on the MNI. The head included the skull and mandible, while the axial included the vertebrae, sternum, and ribs. The proximal portion of the limb bones consisted of the scapula, humerus, innominate, and femur. The distal portion of the limb bones comprised the radius and ulna, tibia, metacarpus, carpus, tarsus, and phalanges. The result of the test for independence using Fisher’s exact test showed no significant difference in the ratio of the four parts between camelids and deer (P = 0.894).
In addition, to confirm chronologic differences in the body part frequency of deer, we compared the data of the Pacopampa I phase and the Pacopampa II phase and found no significant difference (P = 0.498). From Pacopampa I to Pacopampa II, the body part frequency of artiodactyls shows a common feature that proximal limb bones are dominant.
Butchering
The artiodactyls’ skeletal remains were broken and fragmented. However, they were well preserved, with more than 90% of the material falling within Behrensmeyer’s (1978) weathering stage index 1, and butchering marks on the bone surface could be observed. A wide variety of butchering marks were observed in all parts of the body. Cut marks, chop marks, and peeling are associated with skinning, dismembering the joints, and stripping the flesh, while pit, striae, and scrapes are associated with marrow extraction (White, 1992). The carcass was dismembered in both camelids and deer, meat stripped, and the bones were crushed to take marrow.
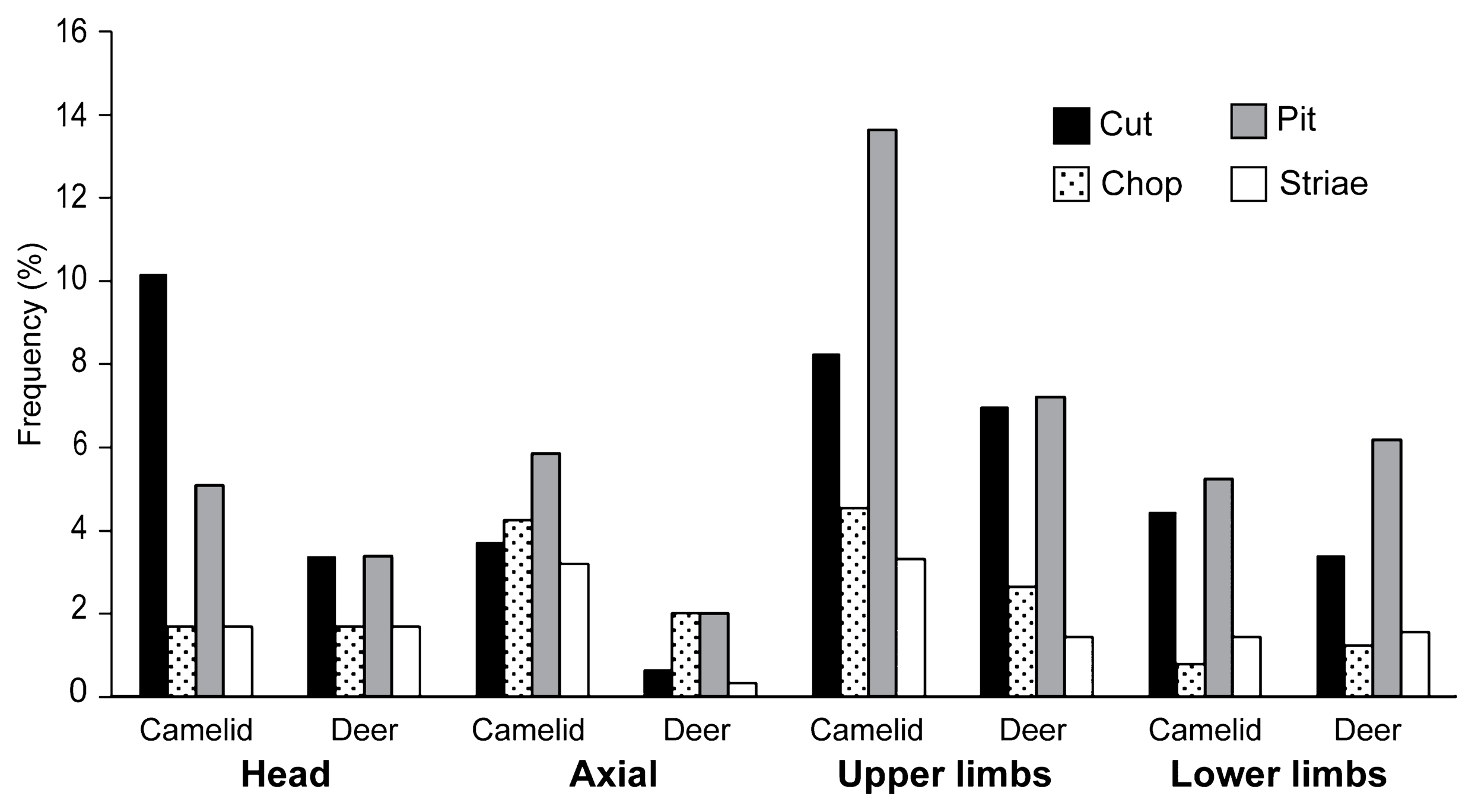
To compare the consumption patterns of camelids and deer, the body was again divided into four parts, namely head, axial, proximal, and distal limbs, and the butchering marks were observed. The elements included in the four groups are the same as those used in the analysis of body part frequency. Table 3 shows the frequencies and distribution of butchering marks. Figure 4 summarizes the frequency of cut, chop, pit, and striae observed in llamas and deer during Pacopampa II. The cut mark and pit distribution are prominent in the proximal and distal limbs of both camelids and deer, indicating that dismembering of the joints, filleting, and bone marrow extraction was concentrated on this part. In the head, cuts were more frequent on the camelids. The cut marks on the maxillary and mandibular bones, where thin muscles are attached, are judged to be skinning traces. However, the NISP for the camelid head was only 59, and there was no significant difference in the frequency of butchering marks between the camelids and deer (χ2 = 2.981, P = 0.084). In the axial (spine and ribs combined), dismantling marks were found more frequently in camelids than in deer (χ2 = 8.798, P = 0.003). It is suggested that the dismantling and use of the spinal column and thorax may have differed between deer and camelids, but it is unclear whether this is due to the anatomic characteristics of camelids or to cultural behavior such as cooking methods. There was a high commonality in the dismemberment patterns of deer and camelids in the limb bones, meat filleting, and marrow extraction. There was no significant difference in butchering mark frequency between camelids and deer in the proximal limb bones (χ2 = 1.76, P = 0.185) and distal limb bones (χ2 = 0.014, P = 0.907). No significant differences were detected when comparing the total body frequency of all elements combined (χ2 = 0.434, P = 0.510).
Table 3
Distribution and frequency of butchering marks observed on artiodactyl bones
|
Dismembering/Filetting |
Marrow extracting |
Total |
% Total |
| NISP |
Cut |
% Cut |
Chop |
% Chop |
Peeling |
% Peeling |
Pit |
% Pit |
Striae |
% Striae |
Crushing |
% Crushing |
Scrape |
% Scrape |
| Head |
| Camelid |
59 |
6 |
10.2 |
1 |
1.7 |
2 |
3.4 |
3 |
5.1 |
1 |
1.7 |
1 |
1.7 |
0 |
0.0 |
13 |
22.0 |
| Deer |
118 |
4 |
3.4 |
2 |
1.7 |
0 |
0.0 |
4 |
3.4 |
2 |
1.7 |
1 |
0.8 |
1 |
0.8 |
13 |
11.0 |
| Axial |
| Camelid |
188 |
7 |
3.7 |
8 |
4.3 |
4 |
2.1 |
11 |
5.9 |
6 |
3.2 |
10 |
5.3 |
0 |
0.0 |
32 |
17.0 |
| Deer |
298 |
2 |
0.7 |
6 |
2.0 |
5 |
1.7 |
6 |
2.0 |
1 |
0.3 |
7 |
2.3 |
1 |
0.3 |
30 |
10.1 |
| Upper limb |
| Camelid |
242 |
20 |
8.3 |
11 |
4.5 |
2 |
0.8 |
33 |
13.6 |
8 |
3.3 |
10 |
4.1 |
3 |
1.2 |
59 |
24.4 |
| Deer |
416 |
29 |
7.0 |
11 |
2.6 |
6 |
1.4 |
30 |
7.2 |
6 |
1.4 |
12 |
2.9 |
0 |
0.0 |
73 |
17.5 |
| Lower limb |
| Camelid |
764 |
34 |
4.5 |
6 |
0.8 |
2 |
0.3 |
40 |
5.2 |
11 |
1.4 |
1 |
0.1 |
0 |
0.0 |
83 |
10.9 |
| Deer |
647 |
22 |
3.4 |
8 |
1.2 |
0 |
0.0 |
40 |
6.2 |
10 |
1.5 |
5 |
0.8 |
5 |
0.8 |
74 |
11.4 |
The frequency of butchering marks in deer was compared between the Pacopampa I and II phases. A significantly high number of butchering marks on the proximal limb bones from the Pacopampa II phase was confirmed (χ2 = 5.411, P = 0.020). There was no significant difference in head (χ2 = 0.892, P = 0.345), trunk (χ2 = 1.349, P = 0.245), and distal limb bones (χ2 = 0.360, P = 0.548).
Mortality profile
The estimated survival curve of the camelids is shown in Figure 5. The use of camelids at Pacopampa was mainly immature individuals. In addition to the data from Pacopampa, the graph also shows the data from the Chavín de Huántar site reported by Miller and Burger (1995) for comparison. The survival rate of camelids excavated from the Pacopampa site was 87% at 1 year of age and 41% at 3 years of age. The majority of the animals die before they reach adulthood. Only 22% survived by the age of 4 years, the upper age limit estimated with the analysis, which was about half of the life span.
We observed the teeth eruption and wear patterns to determine the age composition, paying particular attention to the adult animals, which could not be examined by bone fusion. The results show seven juveniles (<3 years old), two prime adults (<6 years old), and one old adult (>6 years old). Although a significant portion of the animals were slaughtered in the juvenile stage, those beyond the adult stage were long lived.
The age structure of the camelids was then compared with that of the deer to determine whether the camelid mortality pattern reflects this animal’s husbandry management or is just showing the general use of artiodactyls at the site. Deer showed a composition of 17 juveniles, six prime adults, and no old adult specimens. The predominance of a juvenile was also confirmed in the deer. Notably, some materials contained unerupted deciduous teeth, indicating the use of newborn or neonate individuals. The age structure of camelids and deer was tested by the Fisher exact probability test, and no significant difference was found (P = 0.455).
Buried camelids
Four pits with camelid skeletons were found in the Pacopampa II period. One other pit with a camelid skeleton was also excavated in the Cajamarca period that followed the Formative Period and when Pacopampa had ceased its function as a ceremonial center. Based on the conditions of excavation, we conclude that these individuals were killed as sacrifices. The characteristics of each individual are described below.
10PC-C-Ent.01
The whole skeleton remains. The bones are well preserved. Based on the teeth eruption and the state of occlusal wear, the age was estimated to be 6 months. Two cut marks were found on the ventral side of the thoracic vertebrae. Since the individual was found in the anatomically normal position, we concluded that these wounds were not caused by dismemberment but by slaughtering (Figure 6).
10PC-D-Ent.10-2
The whole skeleton remains. Bones are well preserved and present no evidence of human-induced damage. Based on tooth eruption and the state of occlusion, the age was estimated to be 3–6 months.
10PC-D-Ent.10-3
The occipital bone, the first cervical vertebra, metacarpals, metatarsals, and phalanges remain. All four metacarpals and phalanges are present, with no evidence of artificial damage. Only the head and podiums were selectively buried. Based on the state of the metacarpal and second phalange epiphyses, the individual was estimated to be up to 24 months old.
11PC-C-Ent.04
This individual lacks head and cervical vertebrae. All the limb bones, including phalanges, were intact. No artificial damage was observed in the remaining parts. Based on the unfused proximal epiphysis of the radius and the scapula, the age was estimated 9–12 months old.
15PC-A-Ent.11
This specimen was unearthed from the deposit of the Cajamarca period. The entire skeleton has remained. No artificial damage was recognized. The age of the individual was estimated to be 6–9 months old based on tooth eruption.
Discussion
Incompatibility with the vertical control model
The camelids used at Pacopampa comprise mainly juvenile llamas, which do not participate in caravans generally. Alpacas raised in the highlands were not found in the samples. As we have shown in an earlier study, the strontium isotope ratios from the camelid teeth corresponded to the values around the site (Takigami et al., 2020). Taking all of this information together, we believe that llamas were raised in the site’s neighboring area.
Previously, camelid bones recovered from Formative Period sites in valleys were recognized as the remains attached to dried meat (ch’arki), traded from the higher zone (Miller and Burger, 1995). This idea was generated from the observations of modern pastoralists who reside in the puna zone. They breed llama and alpaca at over 4000 masl, where the animals are slaughtered, and carcasses processed into freeze-dried meat with bones. Herders then bring the ch’arki to lower canyons and coasts to be exchanged for crops. Miller (1979) found that the less meaty parts of the animal, such as the head and feet, were left behind at the breeding sites, while the meaty parts of the limbs were brought to the consumption places. The disproportionate distribution of camelid bones owing to dried meat production and trade is known as the ‘ch’arki effect,’ and has been used to interpret bone assemblages from archaeological sites. Miller and Burger (1995) noticed the ch’arki effect on the camelid remains from the Chavín de Huántar site and claimed that there had been trading between highlands and valleys. This idea of an agropastoral economy based on the vertical control model at the most prominent ceremonial center of the Formative Period has become a central model in Andean archaeology.
The llama skeletal remains from the Pacopampa site were composed of fleshy proximal limb elements, which seemed to correspond to the ch’arki effect. However, many heads and digits, which should have been left at production sites, were recovered. In particular, calcaneus was the most frequently found element for camelids. Some bones could have been brought in as ch’arki, but it is unlikely that dried meat was a principal source of camelids at the site. Recent investigation at Chavín de Huántar revealed the corresponding result of this study and has led to claims of the possibility of camel breeding in the valley (Rosenfeld and Bautista, 2017).
Consumption of artiodactyls at Pacopampa
Interestingly, the body part frequency of camelids was consistent with that of hunted deer. Usually, game animals are dismembered at the point of capture, and the selected parts are brought to the consumption place (Perkins and Daly, 1968). In contrast, it is easier for livestock to be accompanied alive to where they are slaughtered and consumed. Despite the difference in the mode of acquisition, camelids and deer nevertheless shared a body part profile at Pacopampa. There must have be a standard manner in which both animals were consumed.
The similarity in age structure between camelids and deer was also noteworthy. It is known that human hunters generally target adult artiodactyls who can provide more nutrition (e.g. Binford, 1979; Stiner, 1994). Deer hunting at Pacopampa was more focused on young animals, which is inconsistent with the general strategy of a hunter-gatherer economy. In the samples, immature individuals, including neonates, indicated an essential purpose for targeting juvenile animals.
The age structure of camelids, where humans can control the animals’ age of death, should further reflect cultural selection. A juvenile-centric composition of camelids has been observed at other Formative Period sites. Shimada (1985) reported that 37% of camels died before 1 year, and 86% were lost by 3.5 years. Although direct comparison was difficult because of the difference in age assessment methods, Wing (1972) noted that the number of adult camelids decreased with the introduction of domesticated forms and that 42% of the animals were killed young in the Higueras period at the Kotosh site. Even more interesting is the age structure at Chavín de Huántar. The age structure of the Pacopampa llamas showed a very similar pattern to that of material from high-status settlements during the Janabarriu period at Chavín de Huántar (Figure 7). Miller and Burger (1995) implied that high-status people consumed the meat of tender young animals. The same explanation can be applied to the case of the Pacopampa site.
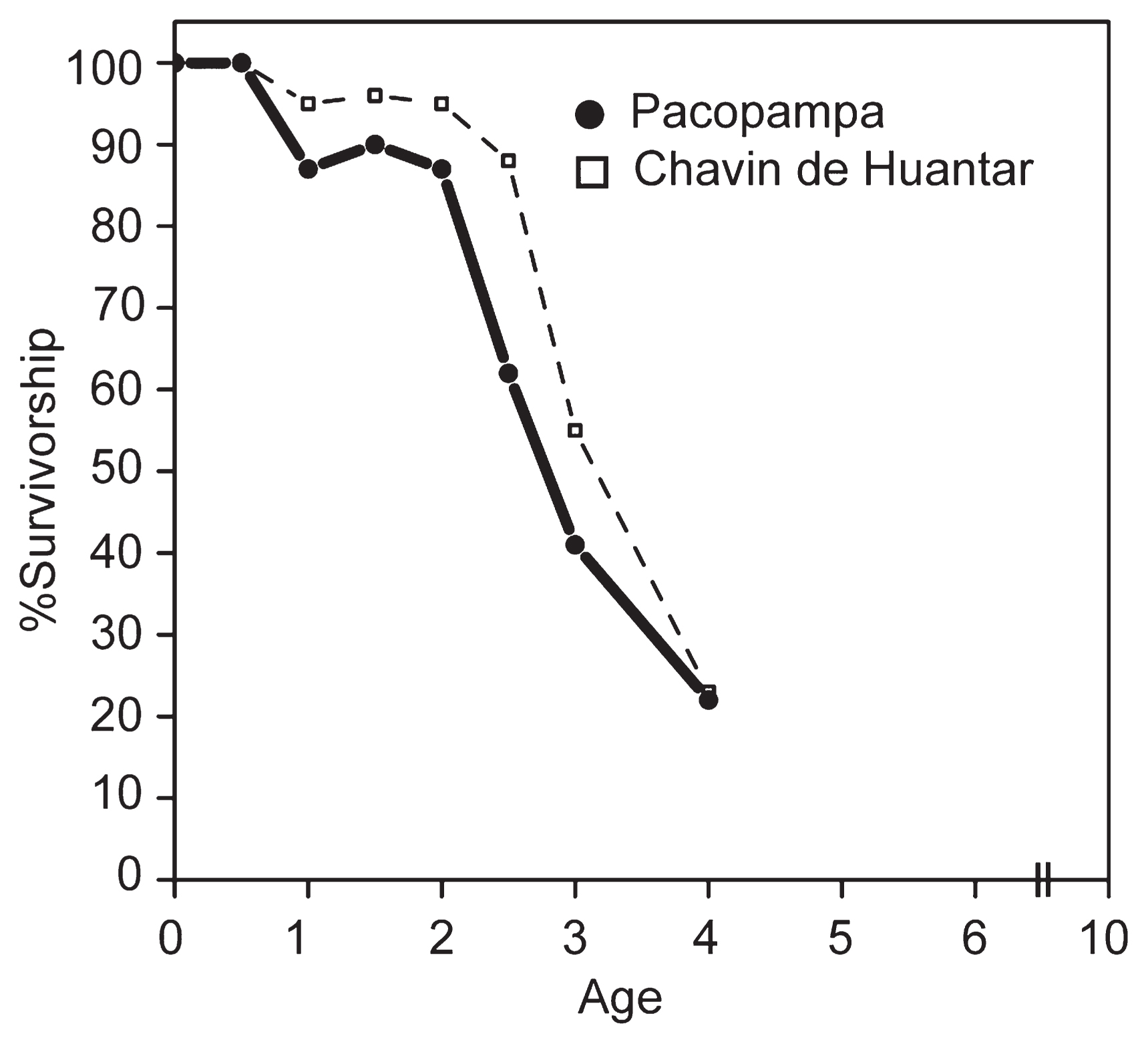
This study also revealed that llamas and deer at the site share the same age structure. The result suggests that the emphasis on young individuals in the Formative Period site was not unique to camelids but can be generalized as a characteristic of artiodactyls. Ritual practice, including feasting, might likely have required tender meat.
Llama sacrifice
What distinguishes regular slaughter from sacrifice is the butcher’s intent and this is difficult to determine through archaeological inquiry (Russell, 2011). Therefore, contextual association has been emphasized in camelid sacrifice in the Peruvian Andes. The five buried camelids found at the Pacopampa site were determined to be sacrifices for the following reasons: they were domestic llamas, buried in the most crucial ceremonial platform, complete or partial skeletons articulated, and included cases with kill marks.
Animal sacrifice played an important religious role in the ancient Andes, and much data have been reported since the Early Intermediate period (Bonavia, 2009). Large-scale camelid sacrifices have been detected in the Moche culture (AD 100–800) on the north coast (Goepfert, 2012; Szpak et al., 2016). However, only fragmentary, unverified references to the late Formative Period site, such as the Kotosh and Cupisnique culture, were reported (Bonavia, 2009). The Pacopampa cases date evident llama sacrifice to the late Formative Period, confirming that this practice began in conjunction with camelid herding at the site.
The age structure of the sacrificed camelid was generally young, as was the case with individuals excavated from other contexts. There was diversity in the sacrifice style, ranging from intact skeleton to partial elements. The variation of the offered parts may reflect a difference of opportunities and purposes for sacrifice. In this sense, the case of 10PC-D-Ent. 10-3 is noteworthy, wherein only the head and podium were offered. In the Moche, the same combination of the body parts is known to be dedicated to the dead. The purpose of Pacopampa llama sacrifice is unknown, but scholars suggest the continuity of the offering from the Formative to the Early Intermediate Period. The case of 10PC-C-Ent.01, which bears cut marks on the ventral side of thoracic vertebrae, even resembles the Inca’s practice of sacrificing llamas by cutting the abdomen (Poma de Ayala, 1980).
Purpose of camelid herding at Pacopampa
At the Pacopampa site, young llamas were consumed in the rituals. However, we must be cautious in concluding that the primary product was the only purpose of llama breeding. At the time of camelid breeding during the Pacopampa II period, we also observe the occurrence of textile-related artifacts (Arata, 2017). Besides ritual use, wool production may have been another purpose of camelid herding. This interpretation does contradict with previously assumed Andean domestic camelid management in two respects. First, alpaca is the primary choice for fiber, and the llama is not a standard choice. Second, contemporary pastoralists engaging in wool production rear the animals until the animals lose their productive capacity. This creates an adult-dominated age structure (Gobel, 2001).
These two dilemmas can be explained as follows. Llama wool can be used as raw material for textiles. The hair diameter, which affects its quality, is continuous between the two species and varies depending on the body part. Current camelid wool production in Argentina is much higher for llamas than for alpacas (Mueller et al., 2015).
As for the age composition, we need to consider the sample’s provenance, excavated from the ceremonial center’s central platform. At Chavín de Huántar, Miller and Burger (1995) reported that older individuals had been recovered from settlements of low-status people. The same phenomenon could have occurred at Pacopampa. In other words, the bodies of young animals were selectively brought to the center of the site for ritual purposes, while those that had grown to adulthood might have been consumed elsewhere and only the hair from them could have been brought to the site for textile production. If the hypothesis that adult animals were raised for wool harvesting is valid, then the possibility of caravanning using these animals must also be considered. Further investigation of the different sectors of the Pacopampa site is required to verify the breeding of adult individuals.
Conclusions
At the Pacopampa site, camelid herding began in Pacopampa II, the late Formative Period. The camelid skeletal remains were determined to be of llamas, and alpacas were not identified in the samples. Body part frequency, butchering mark distribution, and mortality profile revealed that ritual consumption of the meaty part of young individuals was practiced. This consumption pattern was common to deer and can be recognized as a general characteristic of artiodactyl use at the main platform of Pacopampa.
Buried immature llamas were determined to be sacrifices based on their archaeological context. These individuals trace the evident llama sacrifice to at least 400 BC, confirming that this practice began in conjunction with camelid herding. In contrast to the high similarity with the consumption process of artiodactyls, only camelids were sacrificed, while deer were not targeted.
As far as the present study is concerned, the use of camelids was largely limited to the ritual consumption of their bodies. However, the archaeological evidence suggests fiber was obtained as a secondary production.
Acknowledgments
The authors would like to express their gratitude to Dr. Tomohito Nagaoka, Ms. Nagisa Nakagawa, Ms. Megumi Arata, Mr. Juan Pablo Villanueva, Mr. Mauro Ordóñez Livia, Ms. Diana Alemán Paredes, and Dr. Mai Takigami for their invaluable help with the fieldwork and suggestions. This work was supported by JSPS KAKENHI grant numbers JP 16H05639, JP 20H01377, JP20H00050.
Conflict of interest
The authors declare no competing interests.
Author contributions
K.U. analyzed the samples and wrote the paper. Y.S. and D.M. co-directed the excavation, and Y.S. revised the draft.
References
- Arata M. (2017) Pacopampa iseki ni okeru seisan, shohi soshite haiki-sekki., kokkakuki, doseihin, kinzokuki no bunseki kara. In: Seki Y. (ed.), Andes bunmei, shinden kara yomitoru kenryoku no sekai. Rinsen shoten, Kyoto, pp. 133–160 (in Japanese).
- Behrensmeyer A.K. (1978) Taphonomic and ecologic information from bone weathering. Paleobiology, 4: 150–162.
- Binford L.R. (1979) Nunamiut Ethnoarchaeology, Academic Press, New York.
- Bonavia D. (2009) The South American Camelids: An Expanded and Corrected Edition. Cotsen Institute of Archaeology Press, University of California, Los Angeles,
- Burger R. (1992) Chavín and the Origins of Andean civilization. Thames & Hudson, New York.
- Cartajena I. (2009) Explorando la variabilidad morfométrica del conjunto de camélidos pequeños durante el Arcaico Tardío y el Formativo Temprano en Quebrada Tulán, norte de Chile. Revista del Museo de Antropología, 199–212.
- Chicoine D. and Rojas C. (2013) Shellfish resources and maritime economy at Caylán, coastal Ancash, Peru. Journal of Island and Coastal Archaeology, 8: 336–360.
- Fung P.R. (1975) Excavaciones en Pacopampa, Cajamarca. Revista del Museo Nacional, XLI: 129–207.
- Gasco A.V. and Marsh E.J. (2015) Hunting, herding, and caravanning: osteometric identifications of camelid morphotypes at Khonkho Wankane, Bolivia. International Journal of Osteoarchaeology, 25: 676–689.
- Gobel B. (2001) El ciclo annual de la producción pastoril en Huancar (Jujuy, Argentina). In: Goñalons M., Olivera D.E., and Yacobaccio H.D. (eds.), El uso de los camélidos a través del tiempo. GZC/Ediciones del Tridente, Buenos Aires, pp. 91–115.
- Goepfert N. (2012) New zooarchaeological and funerary perspectives on Mochica culture (AD 100–800), Peru. Journal of Field Archaeology, 37: 104–120.
- Goñalons G.L.M. and Yacobaccio H.D. (2006) The domestication of South American camelids: a view from the South–Central Andes. In: Zeder M.A., Bradley D.G., Emshwiller E., and Smith B.D. (eds.), Documenting domestication. University of California Press, Berkeley, Los Angels and London, pp. 228–244.
- Greenfield H.J., Chapman J., Clason A.T., Gilbert A.S., Hesse B., and Milisauskas S. (1988) The origins of milk and wool production in the Old World: a zooarchaeological perspective from the Central Balkans [and Comments]. Current Anthropology, 29: 573–593.
- Haas J. and Creamer W. (2006) Crucible of Andean civilization: the Peruvian coast from 3000 to 1800 BC. Current Anthropology, 47: 745–775.
- Izeta A.D., Otaola C., and Gasco A. (2009) Osteometría de falanges proximales de camélidos sudamericanos modernos. Variabilidad, estándares métricos y su importancia como conjunto comparativo para la interpretación de restos hallados en contextos arqueológicos. Revista del Museo de Antropología, 169–180.
- Kent J. (1982) The Domestication and Exploitation of the South American Camelids: Methods of Analysis and Their Application to Circum-Lacustrine Archaeological Sites in Bolivia and Peru. University Microfilms International, Ann Arbor, MI.
- Lyman R.L. (1984) Bone density and differential survivorship of fossil classes. Journal of Anthropological Archaeology, 3: 259–299.
- Miller G.R. (1979) An introduction to the ethnoarchaeology of the Andean camelids. Ph.D dissertation, University of California, Berkeley. University Microfilms, Ann Arbor, MI.
- Miller G.R. (2003) Food for the dead, tools for the afterlife: zooarchaeology at Machu Picchu. In: Burger R. and Salazar L. (eds.), The 1912 Yale Peruvian Scientific Expedition Collections from Machu Picchu: Human and Animal Remains. Publications in Anthropology. Peabody Museum of Natural History, Yale University, New Haven, CT, pp. 1–63.
- Miller G.R. and Burger R.L. (1995) Our father the cayman, our dinner the llama: animal utilization at Chavín de Huántar, Peru. American Antiquity, 60: 421–458.
- Morales D.C. (1980) El dios felino de Pacopampa, departamento de Cajamarca. Seminario de Historia Andina, Universidad Nacional Mayor de San Marcos, Lima.
- Mueller J.P., Rigalt F., Lamas H., Sacchero D.M., Cancino A.K., and Wurzinger M. (2015) Fibre quality of South American camelids in Argentina: a review. Animal Genetics, 56: 97–109.
- Muñoz S. and Mondini M. (2014) Osteometrics of South–Central Andean camelids: new standards. 12th Conference of the International Council for Archaeozoology (ICAZ), 2014. DOI: 10.13140/2.1.3341.1843.
- Murra J.V. (1972) El ‘control vertical’ de un máximo de pisos ecológicos en la economía de las sociedades andinas, Huánuco, Perú. Universidad Hermilio Valdizán, Pilllco Marca.
- Onuki Y. (1985) The Yunga Zone in the Prehistory of the Central Andes: vertical and horizontal dimensions in Andean ecological and cultural processes. In: Masuda S., Shimada I., and Morris C. (eds.), Andean Ecology and Civilization: An Interdisciplinary Perspective on Andean Ecological Complementarity. Wenner-Grenn Foundation, No. 91, University of Tokyo Press, Tokyo, pp. 339–357.
- Perkins D. and Daly P. (1968) A hunter’s village in Neolithic Turkey. Scientific American 219, 96–106.
- Poma de Ayala F.G. (1980) Nueva corónica y buen gobierno [1615]. Tomo II. Biblioteca Ayacucho, Caracas.
- Pozorski S. and Pozorski T. (2008) Early cultural complexity on the coast of Peru. In: Silverman H. and Isbell W.H. (eds.), The Handbook of South American Archaeology. Springer, New York, pp. 607–631.
- Pulgar V.J. (1981) Geografía del Perú: Las ocho regiones naturales del Perú. Editorial Universo, Lima.
- Rosas La Noire E. and Shady S.R. (1970) Pacopampa: Un centro formativo en la sierra nor-peruana. Universidad Nacional Mayor de San Marcos, Lima.
- Rosenfeld S.A. and Bautista S.L. (2017) Rituals of the Past: Prehispanic and Colonial Case Studies in Andean Archaeology, University Press of Colorado, Boulder.
- Russell N. (2011) Social Zooarchaeology: Humans and Animals in Prehistory. Cambridge University Press, New York.
- Seki Y. (2014) La diversidad del poder en l a sociedad del Período Formativo: Una perspectiva desde la sierra norte. In: Seki Y. (ed.), El Centro Ceremonial Andino: Nuevas Perspectivas para los Períodos Arcaico y Formativo. Senri Ethnological Studies 89. National Museum of Ethnology, Osaka, pp. 175–200.
- Seki Y., Aleman P.D., Ordonez L.M., and Morales C.D. (2019) Emergence of Power during the Formative Period at the Pacopampa Site. In: Burger R., Salazar L., and Seki Y. (eds.), Perspectives on Early Andean Civilization in Peru: interaction, Authority, and Socioeconomic Organization during the First and Second Millennia BC. Yale University Publications in anthropology, Yale University Department of Anthropology, Yale Peabody Museum of Natural History, New Haven, CT, pp. 107–127.
- Shady S.R. (2014) La civilización Caral: Paisaje cultural y sistema social. In: Seki Y. (ed.), El Centro Ceremonial Andino: Nuevas Perspectivas para los Períodos Arcaico y Formativo. Senri Ethnological Studies 89, National Museum of Ethnology, Osaka, pp. 51–103.
- Shimada M. (1985) Continuities and changes in patterns of faunal resource utilization: formative through Cajamarca periods. In: Terada K. and Onuki, Y. (eds.), The Formative Period in the Cajamarca Basin, Peru: Excavations at Huacaloma and Layzon, 1982. University of Tokyo Press, Tokyo, pp. 289–310.
- Stahl P.W. (1999) Structural density of domesticated South American camelid skeletal elements and the archaeological investigation of prehistoric Andean ch’arki. Journal of Archaeological Science, 26: 1347–1368.
- Stiner M.C. (1994) Honor among Thieves. Princeton University Press, Princeton, NJ.
- Szpak P., Chicoine D., Millaire J.F., White C.D., Parry R., and Longstaffe F.J. (2016) Early horizon camelid management practices in the Nepeña Valley, north-central coast of Peru. Environmental Archaeology, 21: 230–245.
- Takigami M., Uzawa K., Seki Y., Chocano D.M., and Yoneda M. (2020) Isotopic evidence for camelid husbandry during the Formative Period at the Pacopampa site, Peru. Environmental Archaeology, 25: 262–278.
- Uzawa K. (2008) La difusión de los camélidos domesticados en el norte del Perú durante el Periodo Formativo. Boletín de Arqueología PUCP, 249–259.
- Uzawa K. (2019) A shift in the use of animals in the Northern Highlands Formative Period: Climate change or social adaptation? In: Burger R., Salazar L., and Seki Y. (eds.), Perspectives on Early Andean Civilization in Peru: Interaction, Authority, and Socioeconomic Organization during the First and Second Millennia BC. Yale University Publications in Anthropology, Yale University Department of Anthropology, Yale Peabody Museum of Natural History, New Haven, CT, pp. 97–105.
- Wheeler J.C. (2012) South American camelids—past, present and future. Journal of Camelid Science, 5: 1–24.
- White T.D. (1992) Prehistoric Cannibalism at Mancos 5MTUMR-2346. Princeton University Press, Princeton, NJ.
- Wing E. (1972) Utilization of animal resources in the Peruvian Andes. In: Izumi S. and Terada K. (eds.), Andes 4 : Excavations at Kotosh, Peru. University of Tokyo Press, Tokyo, pp. 327–352.
- Yacobaccio H.D. (2007) Andean camelid herding in the South Andes: ethnoarchaeological models for archaeozoological research, Anthropozoologica, 42, 143–154.