Original Articles
Human skeletal remains from Hasankeyf Höyük, a sedentary hunter-gatherer site in southeast Anatolia
2022 Volume 130 Issue 2 Pages 121-134

Details
2022 Volume 130 Issue 2 Pages 121-134
Although early Neolithic Anatolia is a key region for the development of sedentary society by modern humans, osteological studies are limited to specimens from the later period or from other regions such as southern Levant. We examined nearly 100 human skeletal remains from Hasankeyf Höyük, a Pre-Pottery Neolithic A (PPNA) site in southeastern Anatolia, which were uncovered during an excavation between 2011 and 2015. A total of 124 individuals were identified as belonging to the Neolithic period. The adult/subadult ratio was close to 1:1, and the sex proportion was skewed towards an abundance in males. A few stature estimates fell within the variation range for Natufian and Neolithic peoples in the southern Levant and Anatolia. The estimated life expectancy was below 30, from 27 to 30 years old, based on age criteria related to dental wear stage. On the basis of dentognathic evidence, the skeletons exhibited heavy occlusal wear for their age, with a lot of obliquely slanted occlusal surfaces and enamel chipping. The observed oral health and dental abrasion patterns are discussed with regards to the people’s biological/behavioral adaptations to the environment, such as the nutritional quality of their diet, alimentary customs, or any indications that the teeth were used as a ‘third hand.’ The Hasankeyf Höyük people are considered to represent very early sedentary villagers in southeast Anatolia who would not have initiated the domestication of plants and animals.
Neolithic Anatolia is a key region for understanding the domestication of wild plants and animals, and the region saw the transition from foraging to farming subsistence at the beginning of the Holocene. Recent reports have argued that while pre-domestication cultivation occurred during the Pre-Pottery Neolithic A (PPNA) (11500–10600 cal BP) time period (Weiss et al., 2006; Willcox et al., 2008; Riehl et al., 2012), domesticated types of cereal appeared in the central-southern Levant at 10700–10200 cal BP (Arranz-Otaegui et al., 2016), then variably spread onto the Anatolian plateau in Turkey at around the same time (Baird et al., 2018). People in Neolithic Anatolia are assumed to have been able to incorporate these waves of subsistence shifts into their sedentary lifestyles, although archeological evidence suggests that the degree to which farming was adopted was not uniform among indigenous communities (Baird et al., 2018).
From a biocultural point of view, the foraging-to-farming transition, as found elsewhere around the world, was presumably related to various climatic, demographic, social, behavioral, and evolutionary factors arising during the terminal Pleistocene and into the Holocene (Larson et al., 2014). Furthermore, the Neolithic transition must have involved an assembly of adaptations, involving changes in population size, nutrition, workload, mobility, and lifestyle, and leading to alterations in health, well-being, and living circumstances (Cohen and Armelagos, 1984; Larsen, 2015).
A large and growing record of studies of human skeletons from archeological settings shows that both the foraging-to-farming transition and the early intensification of agricultural production occasioned a general increase in morbidity and compromised health, along with significant changes in activity, workload, and lifestyle (Cohen and Armelagos, 1984; Larsen, 2015). For example, research based on the human skeletal remains of the Natufian and Pre-Pottery Neolithic periods in the southern Levant have indicated a series of consequences arising from the foraging-to-farming transition that occurred from 15100 to 8350 cal BP (Hershkovitz and Gopher, 2008).
Although plenty of human skeletal remains have been recovered from Anatolia, human bioarcheological studies have primarily focused on burial customs or human behaviors associated with the dead (Kuijt, 2008; Erdal and Erdal, 2012; Erdal, 2015). Secondary treatments of dead bodies or skeletons have been well documented in the regions of Levant and Anatolia, including painting, plastering, defleshing, and skull and/or body part removal (Kuijt, 2008; Özbek, 2009; Erdal, 2015; Gresky et al., 2017). On the other, osteological approaches to the burial customs such as bone preservation or representation of the individual (Bello et al., 2006) have been limited, although they provide a new line for discussions on burial customs.
As for osteological research on early Neolithic human remains in Anatolia, specimens from the time of cereal farming acquisition, i.e. Pre-Pottery Neolithic B (PPNB) onwards, are abundant as found in Çayönü (Özbek, 2004a), Aşıklı Höyük (Erdal, 2018), and Çatalhöyük (Larsen et al., 2015, 2019). In contrast, paleobiological data from the earliest stage of the Neolithic (PPNA), which have been recently reported in Körtik Tepe (Özbek, 2004b; Koruyucu et al., 2018; Erdal, 2021), are still insufficient in both quantity and quality to address the hypotheses and questions surrounding the development of subsistence transitions and the consequences regarding phenotypic changes in human osteology. In this context, a number of human skeletal remains uncovered during the 2011–2015 excavations at Hasankeyf Höyük, a PPNA site in southeast Anatolia, are important because they are one of the biggest human skeletal remains from the earliest Neolithic period in Anatolia. Archeological data from the Hasankeyf site have indicated significant hunting and gathering subsistence from wild resources and a minimal level of plant cultivation (Miyake et al., 2012; Uluçam and Miyake, 2018; Hongo et al., 2019). Osteological approaches to the skeletal remains can provide a new key for their burial customs and also clarify their health status, patterns of mortality, and oral condition, which illuminate the biological basis of how Hasankeyf Höyük people adapted to their regional and temporal environment in the wider context of the Neolithic.
Hasankeyf Höyük is located on the left bank of the Tigris, approximately 2 km east of the well-known medieval site of Hasankeyf in Batman province, Turkey (Figure 1a, b). Excavations of this site were first carried out by a Turkish team in 2009 and have been taken over by a Japanese team from the University of Tsukuba since 2011 (Miyake et al., 2012; Uluçam and Miyake, 2018).

Hasankeyf Höyük. (a) Map of the site and neighboring prehistoric sites along the upper Tigris. (b) Site view. (c) Burial of str73 showing the human bone preservation (%BR = 78.6, %WPB = 32.1).
The site forms a roughly circular mound of around 150 m in diameter and 8 m above the surrounding plain. Except for ephemeral occupational evidence from the Iron Age and the Hellenistic periods in the form of pits dug into the prehistoric layers, all the archeological deposits are mostly from 11000–11500 cal BP as determined by radiocarbon dating of charcoal remains (Miyake et al., 2012). These dates correspond to the PPNA period.
More than 100 human burials have been found, mostly under the floors of subterranean round or rectangular buildings, which were discovered accumulated within a thick PPNA deposit (Figure 2). Most of the human bones were collected from the burials after mapping in situ positions of the bones with the use of dry sieving to collect smaller bone fragments. Among the mammalian fauna, there was a dominance of wild caprine bones, accounting for approximately 56% of the number of identified specimens (NISP) (Hongo et al., 2019). The bones of wild boar and red deer (11% and 6% of the total NISP, respectively) were also common. Rabbits, foxes, and tortoise shell fragments were also regularly uncovered, but aurochs remains were rare in the assemblage. Additionally, in line with large quantities of fish and bird bones recovered, stable nitrogen isotope analysis of human bones based on individual amino acids identified the importance of freshwater resources, although it was not significant and variable among the individuals recovered from different structures (Itahashi et al., 2017).

Excavation plan of Hasankeyf Höyük. Round or semi-rectangular stone walls of the interstratified buildings are indicated with the positions of human burials (stars).
There was little archaeobotanical evidence for the consumption of cereals, such as wheat and barley, but wild nuts, such as almonds, pistachio, hackberry, and lentils, were found (Miyake et al., 2012).
Bioanthropological studyAmong the 107 burial pits belonging to the Neolithic period, more than half were recognized as being primary interments based on skeletal identification and the in situ positions of the bones, i.e. many of the individual bones were arranged in the correct anatomical position (Tashiro, 2014). However, while identifying each bony element, we sometimes encountered more than one skeletal individual within a single burial pit, varying from the intrusion of a single bone or a portion of the body to multiple interments of two or three individuals. After omitting cases of minor bone intrusion, we counted a total of 124 individuals during the 2011–2015 excavation.
The uncovered human skeletal remains from each burial pit were first cleaned and restored with the usual restoration processes (White et al., 2012). During the process of cataloging, we performed individual identification by sorting all identifiable human bones, recording their preservation status, and counting the number of individuals. Bilateral limb bones were identified for each side separately and confirmed as being from the same individual or from an intrusion by another individual. Bone preservation was recorded, using the selected criteria of skeletal parts (Table 1), as belonging to three categories: ‘absent,’ ‘less than half present (<1/2),’ and ‘more than half present (>1/2).’ The pattern of bone preservation was then assessed using two indexes: percentages of well-preserved bone (%WPB) and bone representation (%BR) (Bello et al., 2006). We considered bones for which more than half of the bone survived as well preserved and calculated the %WPB as the ratio of the number of well-preserved (>1/2) bones to the total hypothetical number of bones that should have been present. Bones of the cranium, face, mandible, teeth, vertebrae, ribs, hands, and feet were counted as single elements of the body. Postcranial long bones were counted bilaterally, and unfused epiphyses for immatures were considered as a lump with the respective diaphysis. We considered those bones that were frequently identified among all the recovered individuals as well represented, and calculated the %BR as the ratio of the total number of the identified bones to the total hypothetical number of them. An example of the bone preservation is shown in Figure 1c.
| %BR | %WPB (>1/2) | |
|---|---|---|
| Skull | 61.29 | 22.31 |
| Teeth | 73.39 | 39.52 |
| Vertebra | 54.03 | 4.84 |
| Rib | 63.71 | 9.68 |
| Sacrum | 23.39 | 2.42 |
| Os coxae | 50.81 | 10.48 |
| Clavicle | 32.26 | 18.55 |
| Scapula | 34.27 | 9.27 |
| Humerus | 56.45 | 32.26 |
| Ulna | 51.21 | 27.02 |
| Radius | 47.98 | 23.39 |
| Hand | 71.77 | 28.23 |
| Femur | 62.10 | 34.27 |
| Patella | 18.15 | 16.13 |
| Tibia | 58.06 | 27.02 |
| Fibula | 47.18 | 24.19 |
| Foot | 70.97 | 29.84 |
Sex was initially determined by observing the sexual dimorphic morphologies of the pelvis and skull (White et al., 2012), and we secondarily supplemented the sex identity by metric assessments of canine size and several postcranial dimensions. For the secondary sex assessment measurements, we used the mesiodistal (MD) and labiolingual (LL) diameters of the upper and lower permanent canines and the following postcranial metrics: minimal circumference of the humerus (Martin no. 7), length of metacarpals (M2) (1st–5th), femoral head circumference (M20), femoral midshaft circumference (M8), mediolateral diameter of the patella (M2), minimal circumference of the tibia (M10b), length of the talus (M1), and length of metatarsals (M2) (1st–5th). Some of these measurements have been already acknowledged as having moderate to high validity for sex assessment (see Bass, 1995 for review). Thus, we assigned the sex of unknown individuals by comparing it among the known sex variations (Jidoi, 1997). First, we plotted the quartile ranges of each measurement for males and females initially diagnosed with the pelvic and/or skull criteria, and then we assigned the sex to unknown individuals according to whether the canine and/or postcranial sizes were over the female or under the male ranges. Individuals were assigned as ‘male’ when the measurement of the target specimen was above the known ‘female’ range, and they were assigned as ‘female’ when the size was under the known ‘male’ range. The assignment ‘possible male’ was given to those above the third quartile ‘female’ range and that of ‘possible female’ was given to those below the first quartile ‘male’ range.
Ages at death were estimated via two steps. We first applied morphological age criteria using the tooth eruption sequence for immature individuals (Ubelaker, 2008) and using long bone epiphyseal closure and spheno-occipital synchondrosis closure for subadult–adult criteria (White et al., 2012). We then used three age-related changes: the pubic symphysis surface for adults (Brooks and Suchey, 1990), the auricular surface of ilium (Buckberry and Chamberlain, 2002), and the costal ends of ribs (İşcan et al., 1984; 1985). Combining these with the degree of occlusal dental wear (Smith, 1984), we were able to seriate more individuals into a series of age categories, because dental remains are the most durable of the age-related markers and occlusal wear correlates well with age within populations sharing a similar biocultural setting (Lovejoy et al., 1985). In practice, after recording the dental wear stages of the molars (M1 and M2) according to Smith (1984), we matched the individual scores for the sum of the wear stages (M1 plus M2) with the developmental stage of the pubic symphysis, the auricular surface of the ilium, and the sternal end of the ribs. On the basis of the age profile of individuals, we attempted life table reconstruction and estimated a lifespan for the Neolithic Hasankeyf Höyük people. To compensate for the relatively large variability in age–wear relationships, we used two sets of age-wear patterns. Additionally, we reconstructed life tables with 5- and 10-year intervals to check the feasibility of the reconstructed age profiles.
In addition to lifespan, the health status of the Hasankeyf Höyük people was evaluated based on several skeletal indicators. We estimated the stature from the long bone lengths and used regression by Pearson (1899) and those by Trotter based on European-American and African-American peoples (Trotter, 1970). Because many of the long bone epiphyses were crushed and fragmented, despite the generally well-preserved bone status, the number of specimens available with measurable long bones was quite limited.
Other health indicators were mainly based on the dental remains. We observed the wear of teeth, including wear associated with habitual customs or behaviors, such as occlusal groove or lingual surface attrition (LSAMAT), caries, enamel chipping, cupped teeth, linear enamel hypoplasia (LEH), and calculus attachment. On the postcranial bones, several individuals exhibited trauma and pathological changes to the bone surface (e.g. periostitis), which are indicative of an individual’s health status. However, observations of bone surfaces were limited because of the heavy matrix attachment to the bone surface of a substantial number of specimens.
Lastly, the biological affiliation of the Neolithic people of Hasankeyf was assessed based on their dental crown diameters. We measured both the mesiodistal (MD) and buccolingual (BL) diameters of the maxillary and mandibular permanent teeth with a digital caliper (Mitutoyo Co. Ltd., Kawasaki, Japan) accurate to 0.1 mm. Dental measurements had to be selective because of the degree of occlusal wear or chipping of the enamel, and the data for each sex were pooled. We calculated the total crown area (TCA) or molar crown area (MCA) for the left side of the dentition (Haddow and Lovell, 2003). On the basis of the MD and BL diameters averaged with sides, we then calculated the Penrose shape distances (Constandse-Westermann, 1972) and compared them with those from several prehistoric and historic samples.
Most of the human bones showed good preservation. However, the proportion of intact bone surfaces varied among body parts (Figure 3). This variable durability can be interpreted as being due to taphonomic effects or the selection of bones or body parts (intentional or unintentional) for secondary burial customs. When we counted the presence of bones for all individuals (%BR in Table 1), teeth were most abundant (73.4%), followed by bones of the hand (71.8%), then the foot (71%), and the ribs (61.7%). When we considered the bones that were more than half preserved (%WPB > 1/2 present in Table 1), the descending order changed to teeth (39.5%), femur (34.3%), humerus (32.3%), foot (29.8%), and hand (28.2%). The percentages for vertebra, ribs, sacrum, and os coxae were low. The difference of %BR and %WPB values indicates relative durability of the bone, i.e. the value would decrease greatly if the bone was less preserved although it was well identified among the individuals discovered. In particular, the value for ribs decreased greatly from the %BR value (63.7%) to the %WPB value (9.7%), which was due to the poorer durability of the rib bones during the taphonomic or recovery process. Similar wide gaps between the two counts were seen for hand and foot bones, possibly due to intentional or accidental removal/ ignorance of the bones during secondary burials. In contrast, the gap for the patella was minimal, i.e. patellae were infrequently recovered (18.2%) from the burials, but when they appeared, preservation was almost intact (16.1%). In terms of age differences for bone preservation, the general pattern of %WPB was similar among the different age categories (Figure 4). We noticed that subadult individuals retained more well-preserved bone parts than adults. Furthermore, the %WPB values were higher for younger individuals, including immatures and juveniles, than for adults for many bone parts, except the hand and foot bones. Different %WPB values have previously been interpreted in terms of the durability of the bones or extrinsic factors such as secondary reburial practices (Bello et al., 2006), and immature and sub-adult skeletons were generally less well preserved and their bones less well represented than those of adults in three historical osteological collections. The present data for the Hasankeyf skeletons clearly show different patterns, in which subadult skeletons, which should be less durable, were the best preserved, and even the immature and juvenile skeletons were better preserved than adults for many bone parts, except the hands and feet.

Percentages of bone representation (%BR) and well-preserved bones (%WPB).

Percentage of well-preserved bones (%WPB) among age categories (immature, juvenile, subadult, and adult).
On the basis of the results of the sex diagnoses and the estimation of ages at death, we assigned the individuals to four age categories (Table 2, Figure 5). First, we used age categories of immature (less than c. 6 years old), juvenile (c. 6–12 years), subadult (c. 12–20 years), and adults (over c. 20 years). Concerning the age distribution, more than half were adults, a quarter were immature (less than 6 years old), and many fewer individuals were juveniles or subadults. Of the 124 individuals, we succeeded in allocating sex to 63 of the 81 adults and subadults. The male:female sex ratio was 35:28, which was almost in the normal range of variation but with a few more males than females. For sex diagnoses, we employed metric comparisons of selected bones and teeth in addition to the normal sexual dimorphic observations (Appendix Figures 1–3). The selected metrics generally confirmed the dimorphic distribution of the predetermined sexes. Thus, we were able to sex-diagnose more individuals according to their metric values.
| Immature (<6 years) | 28 | Fetus-neonate | 8 |
| Child | 20 | ||
| Juvenile (6–12 years) | 15 | ||
| Subadult (>12 years) | 15 | Male | 35 |
| Female | 28 | ||
| Adult | 66 | Unknown | 18 |
| Total | 124 |
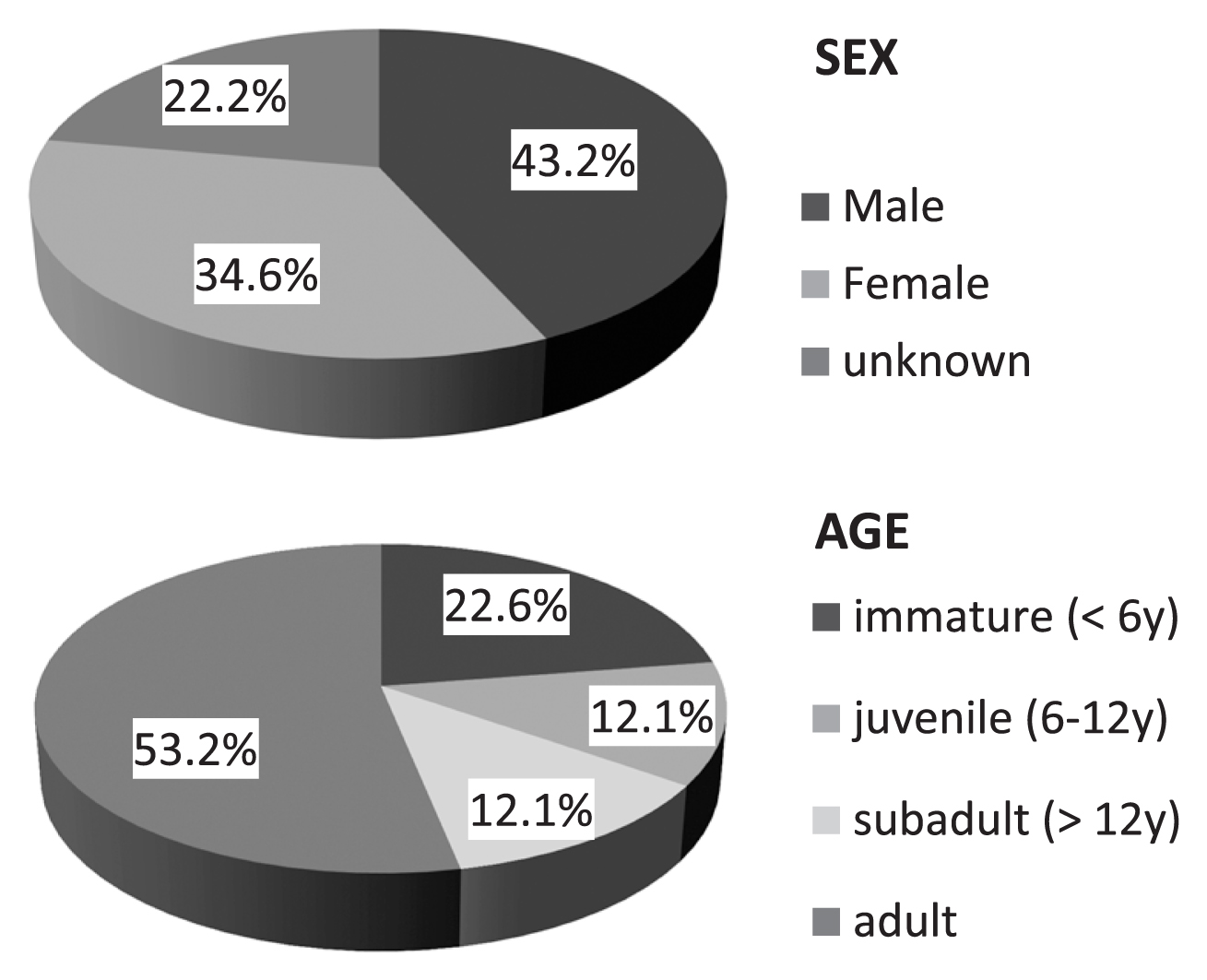
Pie chart of sex and age-at-death frequency.
Table 3 shows the correspondence of the wear stages of molars (M1 plus M2) with those of postcranial developmental signatures of the pubic symphysis, the auricular surface of the ilium, and the sternal end of the ribs. Although the available postcranial developmental stage data were limited in number, a relatively high correlation was found between dental wear and the stages of pubic symphysis and sternal rib end (Spearman’s ρ = 0.78 and 0.77, respectively). Taking into account the large variation in estimated ages within a certain developmental stage, we used two sets of age–wear relationships (age 1 and age 2) by assuming a linear relationship between wear stage and age. The range of dental wear scores (M1 + M2) was 6–31, and this corresponded to ranges of 15–39 years in the age 1 pattern and 15–45 years in the age 2 pattern. We reconstructed life tables for both patterns and for 5- and 10-year age intervals (Table 4). Following the conventional procedure (Chamberlain, 2006), mortality and survivorship parameters were calculated from the age-specific number of deaths. Four patterns of life tables were reconstructed to calculate the life expectancy at age 15 ranging from 11.6 to 15.2 years. Thus, the expected life span (life expectancy at age zero) was from 26.6 to 30.2 years.
| Human no. | Wear stage (M1 + M2) | Pubic symphysis | Auricular surface | Rib sternal end | Age1 (years)a | Age2 (years)a |
|---|---|---|---|---|---|---|
| 134 | 10 | 1 | 0 | 15 | 15 | |
| 201 | 10.5 | 1 | 1 | |||
| 250 | 14.5 | 2 | II | 3 | 20 | 20 |
| 211 | 15 | 3 | I | 1 | 25 | |
| 221 | 16 | 1 | ||||
| 39a | 17 | I | 25 | |||
| 39b | 17 | 4 | ||||
| 219 | 20.5 | 3 | 30 | |||
| 182 | 23 | 3 | 30 | 35 | ||
| 231 | 25 | 3 | ||||
| 25 (1) | 26 | 4 | ||||
| 25 (2) | 26 | 3 | ||||
| 147a | 26 | IV | ||||
| 212 | 27 | 3 | 35 | 40 | ||
| 290 | 27.5 | III | 5 | |||
| 123b | 29 | 3 |
| age1 | x | d’x | dx | lx | qx | Lx | Tx | ex |
|---|---|---|---|---|---|---|---|---|
| 15 | 11 | 25.00 | 100.00 | 0.25 | 437.50 | 1159.09 | 11.59 | |
| 20 | 8 | 18.18 | 75.00 | 0.24 | 329.55 | 721.59 | 9.62 | |
| 25 | 9 | 20.45 | 56.82 | 0.36 | 232.95 | 392.05 | 6.90 | |
| 30 | 10 | 22.73 | 36.36 | 0.63 | 125.00 | 159.09 | 4.38 | |
| 35 | 6 | 13.64 | 13.64 | 1.00 | 34.09 | 34.09 | 2.50 | |
| 40 | 0.00 | |||||||
| 10-year interval | ||||||||
| 15 | 19 | 43.18 | 100.00 | 0.43 | 784.09 | 1204.55 | 12.05 | |
| 25 | 19 | 43.18 | 56.82 | 0.76 | 352.27 | 420.45 | 7.40 | |
| 35 | 6 | 13.64 | 13.64 | 1.00 | 68.18 | 68.18 | 5.00 | |
| 45 | 0.00 | |||||||
| age2 | x | d’x | dx | lx | qx | Lx | Tx | ex |
|---|---|---|---|---|---|---|---|---|
| 15 | 9 | 20.45 | 100.00 | 0.20 | 448.86 | 1454.55 | 14.55 | |
| 20 | 6 | 13.64 | 79.55 | 0.17 | 363.64 | 1005.68 | 12.64 | |
| 25 | 9 | 20.45 | 65.91 | 0.31 | 278.41 | 642.05 | 9.74 | |
| 30 | 5 | 11.36 | 45.45 | 0.25 | 198.86 | 363.64 | 8.00 | |
| 35 | 9 | 20.45 | 34.09 | 0.60 | 119.32 | 164.77 | 4.83 | |
| 40 | 5 | 11.36 | 13.64 | 0.83 | 39.77 | 45.45 | 3.33 | |
| 45 | 1 | 2.27 | 2.27 | 1.00 | 5.68 | 5.68 | 2.50 | |
| 50 | 0.00 | |||||||
| 10-year interval | ||||||||
| 15 | 15 | 34.09 | 100.00 | 0.34 | 829.55 | 1522.73 | 15.23 | |
| 25 | 14 | 31.82 | 65.91 | 0.48 | 500.00 | 693.18 | 10.52 | |
| 35 | 14 | 31.82 | 34.09 | 0.93 | 181.82 | 193.18 | 5.67 | |
| 45 | 1 | 2.27 | 2.27 | 1.00 | 11.36 | 11.36 | 5.00 | |
| 55 | 0.00 | |||||||
d’x = number of deaths in the age x, dx = proportion of deaths, lx = survivorship, qx = probability of death per year, Lx = number of person-years lived between the age x and x + 1, Tx = number of person-years lived after the age x, ex = average years of life remaining at the age x.
Stature was estimated based on the limited number of long bone length measurements. The application of three regression as described above (Pearson, 1899; Trotter, 1970) resulted in slightly different estimates, giving means of 167–174 cm in males and 155–159 cm in females (Table 5). These stature estimates were mostly equivalent to those of Mesolithic to Neolithic populations from adjacent regions in Anatolia (Koca Özer et al., 2011), the eastern Mediterranean (Angel, 1984), western Europe (Meiklejohn et al., 1984), south Levant (Smith et al., 1984), and Iran/Iraq (Rathbun, 1984) (Table 5).
| Site | Calculation method | Male | Female | Reference/region | ||
|---|---|---|---|---|---|---|
| n | Mean (cm) | n | Mean (cm) | |||
| Hasankeyf | Pearson (1899) | 6 | 166.97 | 3 | 154.95 | This study |
| Trotter (1970) European-American | 4 | 173.72 | 3 | 159.22 | ||
| Trotter (1970) African-American | 4 | 168.37 | 3 | 156.15 | ||
| Neolithic | Trotter and Gleser (1952) | 29 | 170.85 | 41 | 155.95 | Anatolia in Turkey |
| Chalcolithic | 7 | 164.96 | 6 | 153.25 | Koca Özer (2011) | |
| Bronze | 46 | 165.87 | 32 | 157.18 | ||
| Upper Paleolithic | Trotter and Gleser (1952) | 35 | 177.10 | 28 | 166.50 | Eastern Mediterranean |
| Mesolithic | 61 | 172.50 | 35 | 159.70 | Angel (1984) | |
| Early Neolithic | 39 | 169.60 | 31 | 155.50 | ||
| Late Neolithic | 6 | 161.30 | 13 | 154.30 | ||
| Late Upper Paleolithic | 19 | 170.40 | 10 | 156.70 | Western Europe | |
| Mesolithic | 46 | 167.70 | 36 | 155.60 | Meiklejohn et al. (1984) | |
| Neolithic | 102 | 167.30 | 88 | 154.10 | ||
| Natufian | NA (based on femur length) | 10 | 167.00 | 3 | 158.00 | South Levant |
| Neolithic | 55 | 168.70 | 43 | 157.50 | Smith et al. (1984) | |
| Chalcolithic | 9 | 168.30 | 10 | 156.20 | ||
| Neolithic | NA | 4 | 172.00 | 5 | 158.00 | Iran/Iraq |
| Bronze | 66 | 174.00 | 35 | 159.00 | Rathbun (1984) | |
The degree of dental wear was scored for M1 and M2 (Smith, 1984). When we compared the seriated molar wear scores with developmental stages of pubic symphysis, the auricular surface, and the sternal end of ribs, we found the degree of molar wear was relatively strong compared with developmental stage according to postcranial indicators. For example, individual Str39a, who was assigned to the 25–29 year age category via auricular surface developmental stage I, showed moderate to strong molar wear (summed score = 17) with a conspicuous decrease in wear along the M1–M3 molar row (wear stages of M1, M2, and M3 were 5, 3, and 1 on the right and 5, 4, and 2 on the left, respectively) (Figure 6). Furthermore, many individuals’ teeth showed signs of habitual use: 40.5% of individuals (15/37) exhibited slanted occlusal surfaces in the molar row, including saddle-shaped wear (Figure 7a). A few cases showed an incised occlusal groove in the anterior teeth and LSAMAT on the upper incisors (Figure 7b).
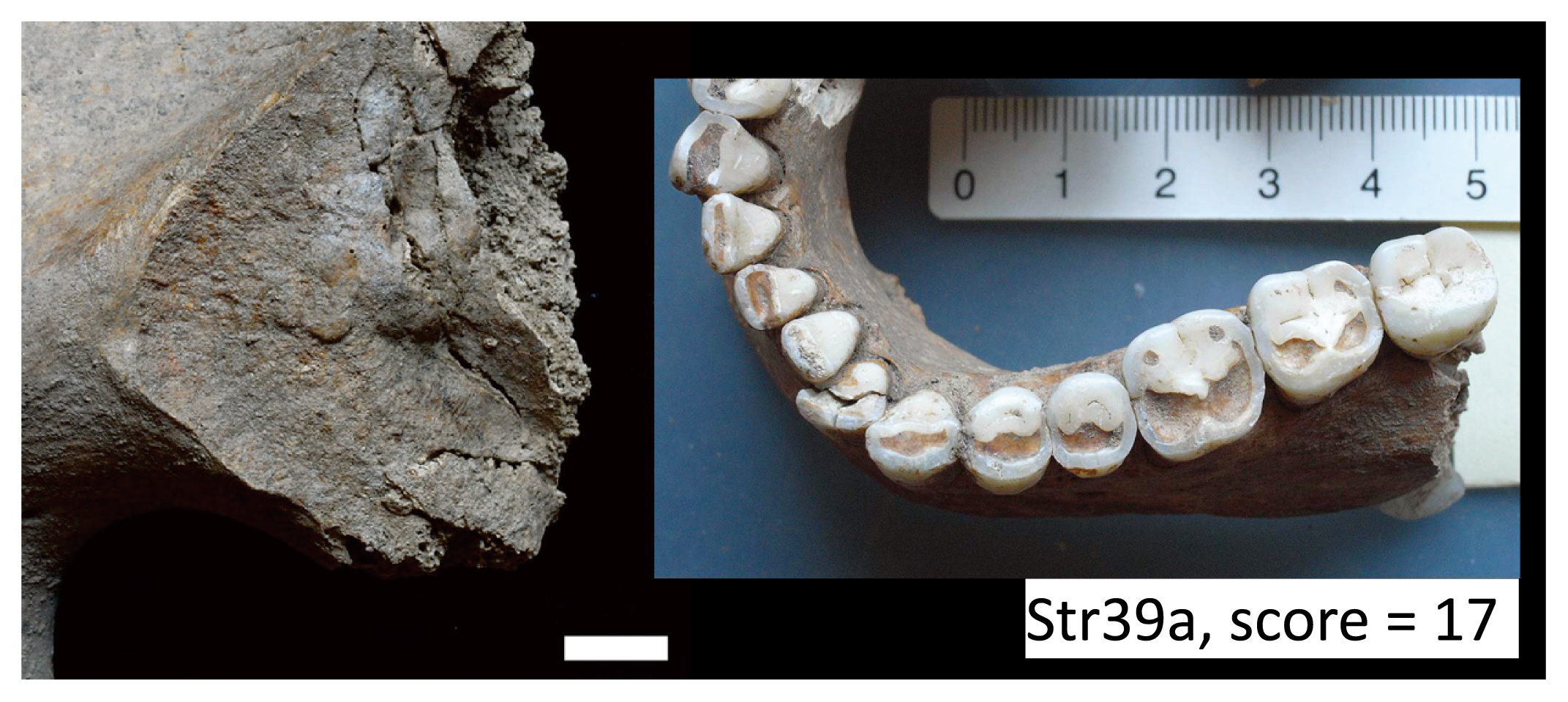
Individual Str39a. Auricular surface of right ilium (stage I) and occlusal wear of mandible (score = 17). Scale = 1 cm.
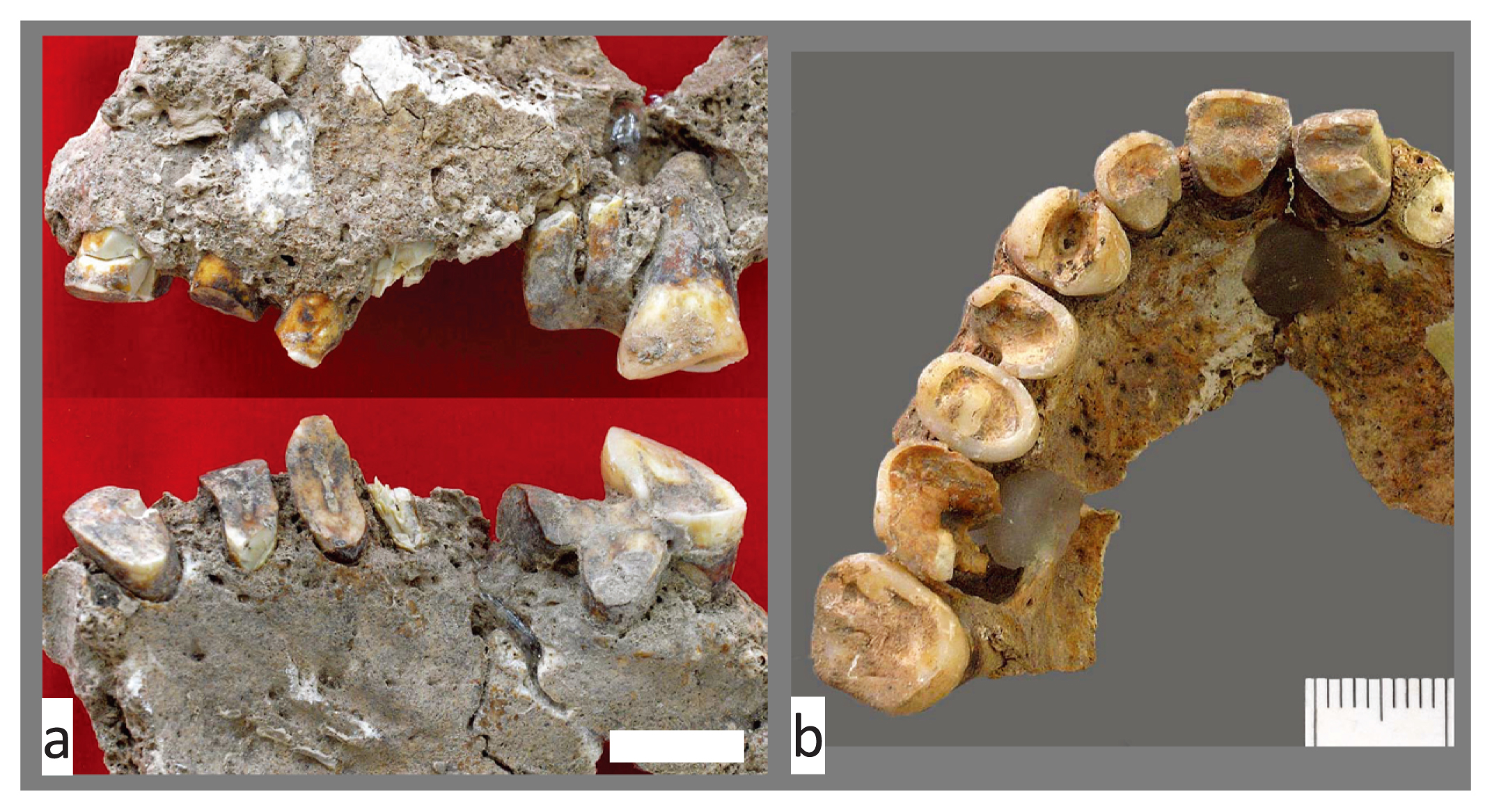
Dental wear and occlusal groove indicating habitual use of the teeth. (a) Slanted and saddle-shaped wear of maxillary teeth (Str27). (b) Occlusal groove and lingual surface attrition (LSAMAT) on anterior teeth (Str25). Scale = 1 cm.
We found only two cases of caries at the neck of maxillary M3 among 1274 teeth (0.16%) from 70 individuals (2.86%). The number of individuals whose molars suffered antemortem enamel chipping was 16 out of 58 individuals (25.9%), and those with concave dentine wear on the occlusal surface (cupped teeth) numbered 32 (55.2%). LEH was observed on the anterior teeth and was individually counted as 32 positive cases out of 54 individuals (59.3%). Individuals with calculus attachment numbered 23 out of 53 (43.4%).
We could not determine the percentages of postcranial bones affected by traumas and pathological changes of the bone surface (e.g. periostitis) because there was insufficient time to remove the attached matrix from many of the post-cranial bones. However, we found several cases of traumatic and pathological bone alterations (Table 6). A relatively severe periostitis-induced bony swelling was observed on the right femoral shaft (Figure 8a, b). A healed fracture of the second metacarpal of the left foot (Figure 8c) and clear joint arthritis of the left ankle (Figure 8d) indicated accidental injury or accumulated injuries from carrying heavy loads. Joint arthritis (Figure 8e) and joint ankylosing (Figure 8f) in the hand phalanges were also indicative of infectious or behavior-related trauma.
| Trauma | Affected parts |
|---|---|
| Periostitis | Tibia and femur |
| Antemortem fracture | Metatarsal |
| Joint arthritis | Ankle (tarocalcaneal joint) and interphalangeal joint (proximal–intermediate) in the hand |
| Joint ankylosing | Hand phalanx (intermediate–distal) |

Paleopathological and traumatic alterations of postcranial bones. (a) Osteoperiostitis on the right femur (Str28). (b) Fractured cross-section of the same (Str28) specimen. (c) Healed fracture of second metatarsal of the left foot (Str29). (d) Talocalcaneal joint arthritis of the left foot (Str46). (e) Joint arthritis between the proximal and middle hand phalanx (Str179a). (f) Joint ankylosis between the distal and middle hand phalanx (Str182).
Tooth sizes of the Hasankeyf skeletons were assessed using MD and BL crown diameters and crown areas (Table 7). We summed the crown areas for all 16 teeth (TCA = 1263.1 mm2) and for the six molars (MCA = 698.2 mm2) on the left side and compared them with fossilized and modern samples (Figure 9). A decline with time has been detected for the summed tooth sizes of both TCA and MCA from Paleolithic to modern samples, and the Hasankeyf samples occupied an intermediate position between the Epi-Paleolithic (Natufian) and the Neolithic (Jarmo and Abou-Gosh) in the southern Levant.
| Maxilla | Mandible | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mesiodistal diameter (MD) | Buccolingual diameter (BL) | Crown area | Mesiodistal diameter (MD) | Buccolingual diameter (BL) | Crown area | |||||||||||||
| n | Mean | SD | n | Mean | SD | n | Mean | SD | n | Mean | SD | n | Mean | SD | n | Mean | SD | |
| I1 | 37 | 9.11 | 0.69 | 40 | 7.49 | 0.46 | 32 | 68.20 | 7.97 | 33 | 5.68 | 0.41 | 29 | 6.33 | 0.49 | 24 | 35.96 | 4.00 |
| I2 | 30 | 7.04 | 0.56 | 32 | 6.64 | 0.49 | 23 | 47.32 | 5.70 | 39 | 6.20 | 0.48 | 36 | 6.66 | 0.52 | 31 | 41.60 | 5.15 |
| C | 37 | 7.76 | 0.41 | 41 | 8.62 | 0.63 | 33 | 66.73 | 7.98 | 38 | 7.05 | 0.43 | 39 | 8.06 | 0.65 | 33 | 56.87 | 7.03 |
| P1 | 34 | 6.97 | 0.37 | 38 | 9.28 | 0.61 | 33 | 64.66 | 6.43 | 39 | 7.12 | 0.39 | 43 | 8.08 | 0.54 | 38 | 58.10 | 5.93 |
| P2 | 37 | 7.01 | 0.69 | 40 | 9.22 | 0.84 | 37 | 64.32 | 8.63 | 39 | 7.29 | 0.39 | 43 | 8.48 | 0.58 | 37 | 62.43 | 6.54 |
| M1 | 42 | 10.73 | 0.65 | 40 | 11.93 | 0.76 | 39 | 128.73 | 14.95 | 54 | 11.45 | 0.76 | 53 | 11.01 | 0.60 | 50 | 127.24 | 14.49 |
| M2 | 43 | 9.75 | 0.63 | 42 | 11.78 | 0.82 | 41 | 115.50 | 13.66 | 52 | 11.01 | 0.72 | 51 | 10.48 | 0.64 | 50 | 115.81 | 13.82 |
| M3 | 20 | 8.78 | 0.63 | 21 | 10.97 | 1.03 | 19 | 96.08 | 14.77 | 26 | 10.88 | 0.77 | 26 | 10.24 | 0.70 | 26 | 111.81 | 14.27 |

Temporal changes in dental crown area (TCA, MCA) from Middle Paleolithic to modern samples. Comparative data are from Haddow and Lovell (2003).
We then estimated the biological relationships among several prehistoric and historic samples from the Levant and Pakistan based on the Penrose shape distance. The Hasankeyf samples were positioned near the Epi-Paleolithic Natufian and Pakistan Neolithic samples (Mehrgarh N) and were distant from the Levantine samples from the PPNB period (Figure 10).
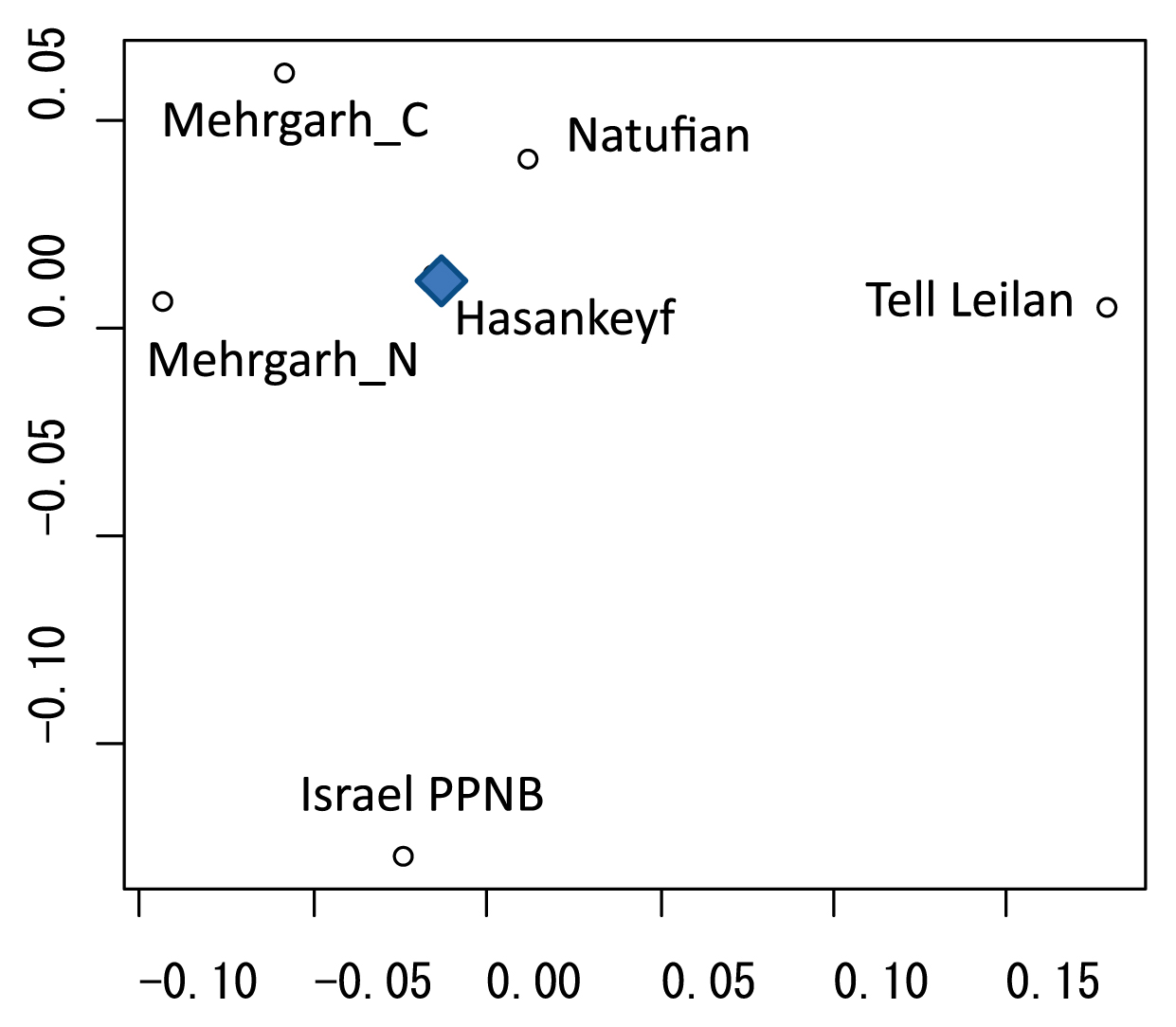
Classic multidimensional scaling (MDS) of Penrose shape distances based on MD and BL diameters of upper and lower permanent dentition (28 variables in total). Comparative samples are Tell Leilan from Bronze Age Syria (Haddow and Lovell, 2003), Natufian in south Levant (Dahlberg, 1960), Mehrgarh N (Neolithic) and C (Chalcolithic) in Pakistan (Lukacs and Hemphill, 1991), and Israel PPNB (data combined from Arensburg et al., 1978 and Hershkovitz et al., 1986).
The %BR and %WPB of the Hasankeyf human skeletal assemblage are characterized by well-represented hand and foot bones and well-preserved younger specimens compared with adults (Figure 3, Figure 4). Both parameters can be considered to change with various factors, including the intrinsic durability of the bones and extrinsic factors during taphonomic or excavation processes. The use of dry sieving yielded a high bone recovery rate for the Hasankeyf specimens; therefore, other causal factors for the observed phenomena in %BR and %WPB are plausible.
The intrinsic durability of bone usually depends on its mechanical properties, i.e. its hardness. Thus, it is assumed that the intrinsic durability of bone varies with age and with the anatomic structures of the body. The bones of infants and juveniles survive less well than those of adults and tend to be underrepresented in skeletal assemblages because they are less dense, more porous, and have smaller hydroxyapatite crystals (Guy et al., 1997; Bello et al., 2006). The Hasankeyf remains have a peculiar feature, in that the bones in the subadult age category are the best preserved, and immature and juvenile specimens are better preserved than those of adults. Anatomic or structural explanations can be considered for the relatively high %BR values of the hand and foot bones from Hasankeyf. Smaller bones and/or bones with high trabecular and low cortical bone contents, such as those of the hands and feet, are intrinsically less well represented in general. On the basis of well-preserved and well-represented skeletal assemblages from medieval adult burials in England, the percentage of hand and foot bones, which is assumed to be equivalent to %BR, ranges from approximately 40% to 65% (Mays, 2010). The even higher %BR values of the Hasankeyf specimens (71–72%, Table 1) need another extrinsic explanation.
To consider the plausible extrinsic factors, we hypothesized possible scenarios involving burial practices. Among other early Neolithic burials in Anatolia, we have found a wide variety of burial practices for both primary and secondary burials, including painting, plastering, defleshing, and skull and/or body part removal (Kuijt, 2008; Özbek, 2009; Erdal, 2015; Gresky et al., 2017). Most of the Hasankeyf burials were considered to be primary burials that were dug into the floors of circular or rectangular stone-walled buildings (Miyake et al., 2012), and immature to subadult individuals were intimately buried and frequently painted with black and red pigments (Tashiro, 2014). This intensive burial practice for subadult individuals might have led to the increased bone preservation. In contrast, secondary burials or the accidental removal of bones (or body parts) would have occurred often in adult burials, whose hand and foot bones escaped removal and remained intact in primary burials.
Sex, age profiles, and life expectancyThe sex ratio of 35:28 (= 1.25) for the Hasankeyf Neolithic people indicates a slight dominance of males compared with females. Although a former pioneering paleodemographic study demonstrated a remarkable bias toward a female-dominant sex ratio in Çatalhöyük human remains (Angel, 1969, 1971), recent excavations have reported moderate female dominance ratios (male:female = 11:13 or 54:63) (Molleson et al., 2005; Hillson et al., 2013). A preponderance of females has been reported for the Neolithic period in Abu Hureyra in Syria (Molleson, 2000), where the recovery of remains inside houses suggested selective burial practices for females. At the Hasankeyf site, where the burial situation seems to have been similar to that of Abu Hureyra, even with the different chronological settings, the apparent male preponderance should be scrutinized in terms of burial contexts, such as chronology, funerary customs, or associated burial goods, because most of the human remains were discovered under the floors and in the vicinity of the walls of circular or rectangular structures (Miyake et al., 2012).
The same arguments can be used for the age profile of the Hasankeyf Höyük people. The detected age profile was not very conspicuous. More than half of the specimens were from adults (c. over 20 years old), 23% were from immatures (less than 6 years), and 12% were from either juveniles or subadults (Figure 5). In contrast, a large number of immature individuals, more than two-thirds of the death assemblage, were reported in Çatalhöyük (Molleson et al., 2005), for which several interpretations were discussed, including selective infanticide, regional and temporal variation in burials, and various burial customs (Andrews et al., 2005; Molleson et al., 2005; Hillson et al., 2013). As already discussed for the preservation of the human skeletal remains, the data from the Hasankeyf specimens should be scrutinized in the light of several aspects, including temporal or customary variations in human burials.
Although we strove to maximize the number of successfully sexed and aged specimens, this may not have been sufficient for successful life table reconstruction. For example, among the several parameters calculated in the life tables, age-specific mortality rates (qx in Table 4) fluctuate for the 5-year intervals, while those in the 10-year interval table increase monotonically after age 15. In a hypothetical natural and stable population, the death rates are expected to increase with age after maturation. In this sense, the reconstructed life tables with 10-year intervals may be more reasonable. Four of the life tables presented variable estimated life expectancies at age 15, ranging from 11.6 to 15.2, indicating an expected lifespan of around 27–30 years. This range is acceptable, at least in comparison with the life table reconstructions for several previous Neolithic findings. For example, Weiss and Wobst (1973) developed model life tables using various anthropological data including those based on historical/prehistorical human skeletal remains, and an expected lifespan (life expectancy at age 0) was calculated for humans from hunter-gatherer societies to Neolithic people that ranged from 19 to 25 years. These life expectancy estimates may be checked and revised with the use of Bayesian and maximum likelihood approaches, which methodologically overcome the long-discussed ‘age mimicry’ problem (Hoppa and Vaupel, 2008).
Health statusStature can be assumed to represent an individual’s gross health status during their growth period. Mean estimated stature based on long bone lengths were c. 170 cm in males and c. 155 cm in females, which are generally comparable to several estimates for ancient peoples from adjacent regions (Table 5). In Anatolia, a decreasing trend in mean male stature has been reported from Neolithic (170.9 cm) to Chalcolithic (165.0 cm), which was discussed in a context of deterioration of health with plant and animal domestication (Koca Özer et al., 2011). In this respect, the health status of the Hasankeyf Höyük people can be considered ‘average’ and almost comparable to those of Neolithic people in the Levant and Anatolia. Or it may be attributable to a ‘pre-domestication’ status before decreasing the stature. A recent Bayesian approach to broader temporospatial variance of stature in the Near East and Europe has clarified the effect of farming transition depending on the region and the mode of Neolithization, and questioned an assumption of a general negative effect of Neolithic lifestyle (Rosenstock et al., 2019). If we accept the observed reduction in stature during the Pre-Pottery Neolithic (PPN) period in Anatolia as an indicator of health trend, its relationship to the change in life-ways should be carefully considered as more complex in view of a recent study detecting a large genetic contribution to body height (Cox et al., 2019).
Some of the oral pathological conditions can be considered indicative of ‘typical’ Neolithic characteristics, such as strong occlusal wear compared to age with obliquely slanted occlusal surfaces, rare caries (less than 3% individually), moderate enamel chipping (c. 30%), frequent cupped teeth (55%), frequent linear enamel hypoplasia (c. 60%), and frequent calculus attachment (43%).
In comparison to data from Neolithic and Natufian peoples of the southern Levant (Eshed et al., 2006), caries cases in Hasankeyf, of which there were only two in the neck of maxillary M3 (calculated to be 0.16% of the tooth count), were exceptionally low in frequency. The caries frequencies of south Levantine Natufian and Neolithic peoples were both c. 6.5% of the total tooth counts and even increased to 10–20% of M2/M3 (Eshed et al., 2006). In comparison to people of the Anatolia region, a trend of increase in dental caries has been reported; 0% from 58 teeth at Körtik Tepe, 4.3% from 1945 teeth at Çayönü, and 3.5% from 567 teeth at Aşıklı Höyük (Özbek, 2007) from PPN to the later Pottery Neolithic (PN) period of Çatalhöyük exhibited 6.6% in tooth count and 10–12% of the permanent dentition affected in both males and females (Molleson et al., 2005; Larsen et al., 2019). The caries frequency at Hakemi Use from the late PN period, however, was as low (3.5% in 342 teeth) as those in the earlier PPN period, where the combination of low caries but moderate-to-high chipping and abrasion status of Hakemi Use people was considered to be indicative of hard, coarse-grained, and fibrous food intake while wheat and barley were already domesticated (Erdal, 2013). Similarly, the lower frequency of caries in the Hasankeyf Höyük people may be attributable to a combination of habitual and dietary factors. Conspicuous degrees of dental wear and many obliquely slanted, sometimes saddle-shaped, wear surfaces on the molars indicate that the people used their teeth as tools. This is supported by the observation of occlusal grooves and labial surface attrition on the incisors (Figure 7). The rapid rate of wear and/or intensive use of teeth as tools quickly deteriorates dental material, which may lead to the loss of cariogenic materials. In terms of diet, it is commonly accepted that the onset of agriculture caused people to shift to diets rich in carbohydrates, promoting caries (Turner, 1979; Cohen and Armelagos, 1984; Meiklejohn et al., 1984), and the elevated levels of dental caries in the Çatalhöyük population are considered to reflect a focus on domesticated plant carbohydrates (Larsen et al., 2015, 2019). On the basis of archeobotanical studies from Iranian and Turkish sites, the size of wild cereal grains from the Iranian site increased during the pre-domestication time period after Younger Dryas, in contrast, the Turkish site of Körtik Tepe uncovered very few wild progenitor species, and shows a variety of cereal cultivation processes in the PPNA period (Riehl et al., 2012). The botanical remains from Hasankeyf included wild nuts, such as almond, pistachio, hackberry, and lentils, but cereals such as wheat and barley were rare (Miyake et al., 2012), which is similar to those from Körtik Tepe. In this context, the people of Neolithic Hasankeyf, who exhibited the lower caries frequency, were presumably conservative users of cereal domestication.
Biological affiliationA general trend of dental size reduction with time has been confirmed in populations of the Near East from Paleolithic to modern times, and the dental sizes of the Hasankeyf Höyük people are appropriate for the site’s position in the PPNA (Figure 9). The trend of reduction in dental size has previously been reported in Anatolian samples, where the rate of reduction from the Neolithic to the Islamic time period was more conspicuous in the molar crown area than in the anterior dentition, suggesting a plausible effect of subsistence transition to an agricultural lifestyle, diet, or food preparation techniques as well as a genetic mutation effect or relief of selective pressure on molar crown size (Erdal, 1999). Although this scenario may be feasible in Anatolia, no mechanisms for dental size reduction have been successfully identified in local post-Holocene cases in the Levant (Pinhasi et al., 2008) or in human evolutionary lineages (Gómez-Robles et al., 2017).
The biological affiliation of the Hasankeyf Höyük people was estimated based on the dental metrics for a handful of available comparative samples (Figure 10). Taking the temporal and regional variation into consideration, it is interesting to note that Neolithic Hasankeyf (PPNA in Anatolia) was close to Natufian in south Levant and distant from later Neolithic Israel (PPNB in south Levant). Although this picture cannot be indicative of the whole population history in this region due to the small sample size, it suggests a possibility of a relatively distant biological relationship between the peoples of Anatolian PPNA and those in south Levantine PPNB.
The paleobiology of the Hasankeyf Höyük people, who may represent the one of the earliest sedentary hunter-gatherers in Anatolia, was described based on human skeletal remains. Among the observed characteristics, noticeable contrasts were found in the rate of caries among temporal and regional comparative samples; the Hasankeyf Höyük people possessed only two M3 cervical caries among over 1000 teeth, whereas people from later PN Çatalhöyük and those from the Natufian to Neolithic (PPNA–PPNB) south Levant exhibited a higher frequency of dental caries. This observation may be relevant to the dietary habits of the Hasankeyf Höyük people, whose subsistence would not have incorporated plant domestication (Miyake et al., 2012).
Infrequent use of cereals has been reported for neighboring contemporary sites, including Körtik Tepe, Hallan Çemi, and Demirköy Höyük (Savard et al., 2006; Coşkun et al., 2010; Riehl et al., 2012). This picture is very different from that for the Middle Euphrates, where large-seeded grasses were extensively exploited as early as the PPNA and for which ‘pre-domestication cultivation’ has been discussed (Willcox et al., 2008). Together with archeological evidence from other contemporary sites in the upper Tigris Valley, further investigations of human skeletal remains around this region would contribute to our understanding of the biology of PPNA people in Anatolia and its relationship to the origin of sedentism in this area, for which a different scenario from that prevailing in the south Levant may be documented.
We are grateful to Prof. Dr. Abdüsselam Uluçam (Director of Hasankeyf Excavations and Conservation Projects) and the General Directorate of Antiquities and Museums, Ankara for providing the opportunity and permission to excavate Hasankeyf Höyük. We thank Prof. Yilmaz Erdal (Hacetepe University) for providing storage for the human skeletal remains, free access to them, and valuable discussion and comments. Thanks also to Prof. Hitomi Hongo (Sokendai University) for checking the faunal records. We also thank all the members and collaborators of the Hasankeyf excavations for their consistent work and help during this research. This study was supported by Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (grant numbers 24251013, 16H05775, and 18H00735). We thank Suzanne Leech, PhD, from Edanz Group (https://en-author-services.edanz.com/ac) for editing a draft of this manuscript. We have no conflict of interest to declare.