Abstract
To produce tomato fruits with 6% average Brix without decreasing yield, we investigated the effect of moderate salinity stress on Brix and yield in a single-truss, high-density tomato production system. Because tomato fruit development can be predicted from cumulative temperature, we also assessed cumulative temperature after anthesis as a potential indicator for determining the starting points of salinity stress treatments. When transverse diameters of the first fruit reached 4 cm (i.e., early increase treatment) or the first fruits were at the mature green stage (i.e., late increase treatment), nutrient solution electrical conductivity was slowly increased until the breaker stage from 1.8 dS·m−1 to 6.0 dS·m−1. Plants subjected to the late increase treatment produced tomato fruits with Brix values of 6% without reductions in marketable yield. We also increased nutrient solution electrical conductivity based on cumulative temperature after anthesis and found that early-treated plants produced tomatoes with higher Brix levels and yields than late-treated plants. In summary, moderate salinity stress to avoid excessive stress on plants increased sugar concentrations without decreasing fruit yield and resulted in tomato fruits with average Brix of about 6% when nutrient solution electrical conductivity was increased at a rate of 0.1 dS·m−1·day−1. Because seasonal differences in cumulative temperature influence the appropriate timing of salinity stress applications, further study is needed to optimize year-round growth under moderate salinity stress in single truss, high-density tomato production systems.
Introduction
Recent consumer interest in tomatoes has focused on flavor quality, with fruits having high Brix (soluble solid) levels in great demand. Because market transaction prices are high, production of high-Brix tomato fruit is potentially very profitable. Sugar concentrations of tomato fruit can generally be increased by applying water or salt stress to plant root zones before harvest (Adams and Ho, 1992; Ehret and Ho, 1986; Saito et al., 2006). Although increased sugar concentrations can be achieved by beginning stress treatments after early fruit set, reductions in fruit weight due to the effects of strong stress are observed (Okano et al., 2002). In addition, earlier application of salt stress to tomato plants, such as immediately after flowering during periods of high temperature, frequently results in blossom-end rot (BER) (Watanabe, 2006). In multi-truss tomato production, advanced techniques are required to minimize BER occurrence on the upper trusses and maintain plant vigor under the stress conditions needed to produce fruit with high sugar concentrations.
Single-truss, high-density tomato production is designed for year-round short-term cultivation and production. Compared with multi-truss tomato production, the single-truss system is simple and eases cultivation. The production period of single-truss plants is very short, as only one fruit truss is harvested. Because Brix levels fluctuate in fruits from multi-truss production with salinity stress, the single-truss system is also advantageous for uniform production of high-sugar fruits. Tomatoes with high soluble solids have in fact been successfully obtained using this system (Araki et al., 2009; Sakamoto et al., 1999).
Increases in sugar and total soluble solids in tomato fruits subjected to salt or water stress are due to decreased water uptake (Bolarin et al., 2001; Chretien et al., 2000; Ehret and Ho, 1986; Li et al., 2001). Because of the tradeoff between increased sugar concentration and fruit size, however, final yields per plant are markedly reduced. Saito et al. (2006) and Sakamoto et al. (1999) have also reported that addition of sodium chloride (NaCl) to nutrient solutions increases tomato fruit sugar concentrations, but tends to decrease fruit yield. BER and cracking also increase, further reducing the final yield per plant. Development of a method to boost tomato fruit sugar concentrations without decreasing fruit size is thus a desirable goal.
Current techniques for high-sugar tomato production subject plants to excessive water or salt stress, which results in decreased fruit yield. Previous studies have examined the effect of salinity stress at different growth stages in a low-truss, high-density tomato production system (Saito et al., 2006; Sakamoto et al., 1999), but a successful method for increasing sugar concentration without inhibiting fruit growth has not been identified. In the present study, we investigated the effect of moderate salinity stress, achieved by gradually increasing nutrient solution concentration, on sugar concentration and fruit yield. For stable year-round production, we also examined the potential use of cumulative temperature after flowering as a starting point indicator for nutrient solution increase. Our goal was the production of tomato fruit with average Brix of 6% without decreases in fruit yield.
Materials and Methods
Plant materials
Seeds of tomato plants (Solanum lycopersicum L. ‘Momotaro York’; Takii Seed, Kyoto, Japan) were sown in a 128-cell tray filled with commercial substrate (Natera; Mitsubishi Plastics Agri Dream, Tokyo, Japan). The seeds were germinated in the dark and grown in a temperature-controlled chamber equipped with fluorescent lamps (Nae Terrace; Mitsubishi Plastics Agri Dream) for 21 days. The chamber was operated under a photosynthetic photon flux density of 280 μmol·m−2·s−1, with a 12 h photoperiod, 22/17°C day/night temperature, and 1200 μmol·mol−1 CO2. The tray was sub-irrigated daily with a commercial nutrient solution (High Tempo; Sumitomo Chemical, Tokyo, Japan) containing 15.4 mM NO3-N, 1.0 mM NH4-N, 39.3 mM PO43−, 9.4 mM K+, 13.1 mM Ca2+, and 5.4 mM Mg2+.
Experiments were carried out in a Venlo-type greenhouse (18 m × 18 m × 4.5 m; NS-oriented) covered with an ethylene tetrafluoroethylene film (F-CLEAN; AGC Grenn-Tech, Tokyo, Japan) at Chiba University, Japan. Three weeks after raising seedling, 40 seedlings were transferred to a greenhouse-based recirculating nutrient film technique (NFT) system (Tomatori-na; Mitsubishi Plastics Agri Dream) with a slope of 0.5% and a nutrient solution flow rate of 5 L·min−1. Intra- and inter-row spacings were 10 and 110 cm, respectively, with a planting density of 9.09 plants·m−2. The Enshi formula nutrient solution used in this study contained 12.0 mM NO3-N, 1.0 mM NH4-N, 3.0 mM P, 6.0 mM K, 3.0 mM Ca, and 1.5 mM Mg, and had an electrical conductivity (EC) of 1.8 dS·m−1. The solution was held in 100-L tanks, each supplying two beds. Plants were pinched off just below the second truss, and all lateral shoots were removed. After pinching, planting beds were wired opposite the fruit truss, and leaves were pulled back and attached to the wire to expose the first-truss fruits (Johkan et al., 2013). At anthesis, the flower truss was sprayed with 4-chlorophenoxyacetic acid (Tomato Tones; ISK Biosciences, Tokyo, Japan). The greenhouse was maintained at a minimum temperature of 10°C. Five fruits matured on each truss and were harvested.
Leaf areas were measured as projected areas using a digital camera and image analysis software (Scion Image, National Institutes of Health, Frederick, MD, USA). Fruits at the red stage were harvested from 20 plants and classified as marketable or unmarketable, with the unmarketable fruit further categorized for BER, core rot (CR), cracking, hollowing, and catfacing. Ascorbic acid (AsA) concentrations were measured on an RQ-Flex plus reflectometer (Merck, Darmstadt, Germany), and sugar concentrations were determined using a refractometer (IPR-101; Atago, Tokyo, Japan). Titratable acidity of fruits was examined by the titration method. Tomato fruit was homogenized and then passed through filter paper. The filtrate was diluted 10-fold with distilled water. That sample was then titrated with 0.1N NaOH. Titratable acidity was expressed as the citric acid concentration.
Effect of timing of nutrient concentration increase on tomato yield and quality (Exp. 1)
Tomato seeds were sown on 25 May, 2012 and placed in the chamber 3 days later, with seedlings transferred to the recirculating NFT system on 19 June. Seedlings were subjected to one of two treatments: when transverse diameters of more than half of first fruits in a bed reached 4 cm (i.e., early increase treatment) or more than half of first fruits in a bed were at the mature green stage (i.e., late increase treatment), the EC of the nutrient solution was slowly increased until the breaker stage from 1.8 dS·m−1 to 6.0 dS·m−1 using 100× stock of Enshi formula nutrient solution (Fig. 1A). Early increase treatments were started on 13 July, and late increase treatments on 25 July, with targeted nutrient solution rate increases of 0.34 and 0.65 dS·m−1 per 2 days, respectively. Average maximum and minimum temperatures were 35.6 and 22.0°C, respectively.
Effect of cumulative temperature-based timing of nutrient concentration increase on tomato yield and quality (Exp. 2)
Tomato seeds were sown on 20 August and placed in the chamber 3 days later. On 11 September, seedlings were transferred to the recirculating NFT system. In Exp. 1, post-anthesis cumulative temperatures at the start of early and late increase treatments were 287 and 650°C·days, respectively (Fig. 1A). The EC of the nutrient solution was therefore slowly increased from 1.8 dS·m−1, beginning at either 287 or 650°C·days, to 6.0 dS·m−1 at 1000°C·days (breaker stage) using 100× stock of Enshi formula nutrient solution (Fig. 1B). Early and late increase treatments were started on 13 and 30 October; targeted nutrient solution increases for early and late increase treatments were 0.10 and 0.17 dS·m−1·day−1, respectively. Average maximum and minimum temperatures were 28.6 and 15.7°C, respectively. First through fifth tomato fruits were harvested separately.
Statistical analysis
Tukey’s multiple range test was used to test differences among more than two means at the 0.05 significance level using XLSTAT software (Esumi, Tokyo, Japan).
Results
Effect of timing of nutrient concentration increase on tomato yield and quality (Exp. 1)
Tomato fruit yield, marketable fruit number, and marketable fruit fresh weight (FW) were unaffected by the timing of nutrient solution EC increase, but marketable fruit yields from plants subjected to the late increase treatment were significantly higher than those from early-treated plants (Table 1). Percentages of marketable fruit from control, early, and late increase treatments were 76, 75.6, and 88%, respectively. The total incidence of BER and CR in fruit from treated plants was higher than that of controls, while the highest percentage of fruit cracking was recorded from early-treated plants (Table 2). Fruit BER under each treatment was less than 3%. In addition, irregular fruit and hollow fruit were observed on control and late-treated plants.

Elevated Brix levels were observed in tomato fruits from treated plants, with the 5.9% measured from late-treated plants being significantly higher than the 5.1% control level (Table 3). Titratable acidity of tomato fruit from early increase treatments was significantly higher than that of controls, whereas AsA concentration, leaf area, and shoot and root dry weights from the different treatments were statistically unchanged (data not shown).
Effect of cumulative temperature-based timing of nutrient concentration increase on tomato yield and quality (Exp. 2)
Fruit yields of plants subjected to the early increase treatment were significantly lower than those of control plants, whereas the yield, number, and FW of marketable fruit were unaffected by early and late EC nutrient solution increases whose timing was based on cumulative temperature (Table 1). The percentages of marketable fruit from control, early and late treatments were very high: 90.9, 94.8, and 98.7%, respectively. Hollow fruit percentages from control and early treatments were 7.8 and 3.9%, respectively, while 1.3% of fruits from each treatment were catfaced (Table 2).
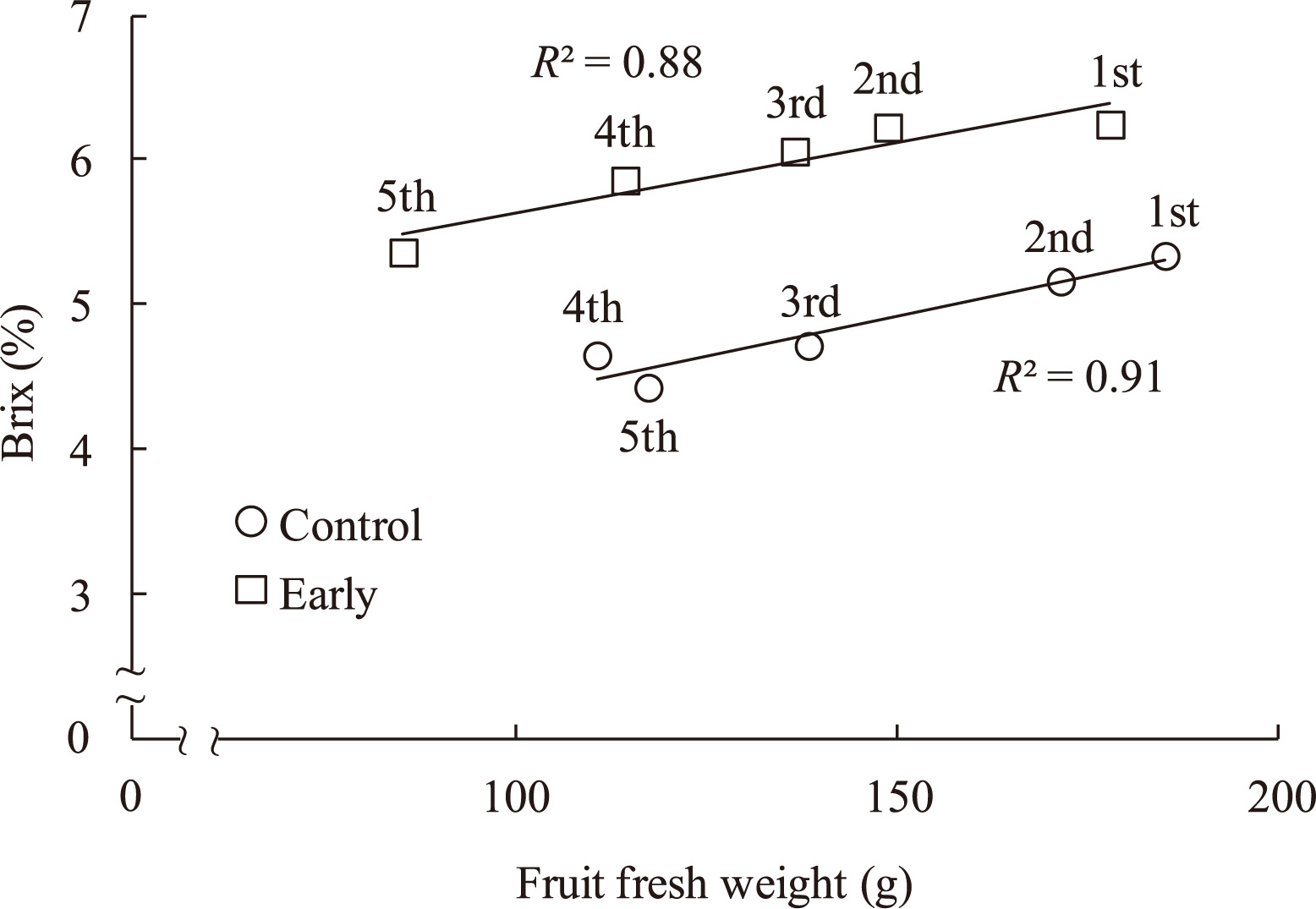
The highest soluble solids concentration, 6.4%, was measured in tomato fruit from early-treated plants. Brix levels of control and late-treated plants were 5.4 and 5.7%, respectively (Table 3). Sugar concentration and fruit weight were positively correlated, with the highest sugar concentrations and fruit weights observed in early-set fruit (Fig. 2). The largest fruits had higher soluble solids concentrations, and the sizes and soluble solids concentrations of first and second fruits were larger and higher than those of other fruits. Titratable acidities of tomato fruit from the two treatments were significantly higher than control levels. The highest AsA concentration, 20.7 mg/100 g FW, was observed in fruit from early-treated plants; AsA concentrations of control and late-treated plants were 15.5 and 17.6 mg/100 g FW, respectively (Table 3). Leaf area and shoot and root dry weight were unaffected by increases in nutrient solution concentration (data not shown).
Discussion
In conventional high-sugar tomato fruit production using salt stress, the EC of the nutrient solution is rapidly increased during the growing period by adding NaCl or high concentrations of stock solution. Saito et al. (2006) reported that tomato fruit Brix increased from 6.1 to 9.9% in a third-truss, high-density tomato production system when nutrient solution EC was increased and maintained at 8.0 dS·m−1 from first truss anthesis until fruit harvest by adding NaCl. Wu and Kubota (2008) also reported that tomato fruit Brix increased from 5.3% to 6.1% when nutrient solution EC was increased to 4.5 dS·m−1 at the pink stage. Although fruit sugar concentrations were increased by limiting water inflow to the fruit using salt stress, fruit yield was reduced as tomato fruits became smaller. Our results demonstrate that it is possible to increase sugar concentrations without significantly reducing fruit yield using moderate salinity stress. In addition, our results indicate that moderate salinity stress can be used to produce tomato fruit with 6% Brix.
During high-sugar tomato production, physiological disorders can easily occur because water inflow to fruit is limited. High salinity stress increases the incidence of BER and CR by interfering with Ca2+, decreasing its uptake by roots and its transport via xylem to fruits (Belda and Ho, 1993; Belda et al., 1996; Ho et al., 1993). Adams and Ho (1992) reported that BER incidence in tomatoes cultivated at EC 13.6 dS·m−1 was 3.7–4.4-fold higher than in those grown at EC 2.4 dS·m−1. In Exp. 1 of our study, CR incidence in tomatoes from early- and late-treated plants under salinity stress was 3.0 and 2.3 times that of the controls, respectively, while total incidence of BER and CR in early- and late-treated fruit was slightly increased, to 1.9 and 1.3 times that of the controls, respectively. Moreover, in Exp. 2, the percentage of tomatoes with physiological disorders after salinity stress treatments was less than 6%, and the yield of unmarketable fruit was not increased by high nutrient solution concentrations. These results suggest that a consequence of avoiding rapid increases in nutrient solution concentration could allow the normal growth of tomato fruit because of stabilized water inflow. Our results also suggest that the incidence of unmarketable fruits from plants treated with moderate salinity stress is less than that obtained from tomato production using conventional salinity stress.
To ensure stable production of high-sugar tomato fruit regardless of the season, the timing of nutrient solution concentration increase is important. In Exp. 1, the starting points of early and late salinity stress treatments were when first fruit had 4-cm transverse diameter and at mature green stages, respectively. In commercial production, however, objective management indicators are preferable to visual monitoring methods. The process of maturation in tomato fruits can be predicted from cumulative temperature: in tomatoes, about 1000°C·days are required from flowering to fruit maturation (Kita, 1987). We therefore examined the effect of applying nutrient solution concentration increases according to cumulative temperature. Because cumulative temperatures at early and late treatment starting points in Exp. 1 were 287 and 650°C·days, respectively, the required daily concentration increase in Exp. 2 was calculated by predicting the date on which the 1000°C·day would be reached. Brix values of tomato fruits from early- and late-treated plants in Exp. 2 were increased with no reduction in fruit yield. It thus appears that salt stress treatment involving a gradual increase in nutrient solution EC at a rate determined by increases in cumulative temperature can minimize tomato fruit growth inhibition.
Seasonal differences in fruit development due to cumulative temperature are large. For example, diameters of fruits subjected to early and late treatments in Exp. 2 were 3 and 7 cm, whereas those in Exp. 1 were 4 and 6 cm, respectively. Moreover, in our study, the cumulative temperature at the breaker stage of tomato fruits in winter was more than 1500°C·days (data not shown). The inversion of Brix values in tomato fruit subjected to moderate salinity stress in Exps. 1 and 2 may therefore be attributed to differences in the degree of fruit development and salinity stress related to fruit developmental stage. It may thus be necessary to determine the cumulative temperature of fruit developmental stages for each season and to take into consideration effective temperature zones.
Although the relationship between cumulative temperature and tomato fruit development from flowering to maturity is strong (Kita, 1987), fruit in Exps. 1 and 2 were at different developmental stages at the starting points of early and late salinity treatments. Consequently, we are unable to provide a definitive answer regarding the starting points of salinity stress in this study. Nevertheless, tomato fruit Brix values were significantly increased by gradual increases in nutrient solution EC with no corresponding significant decreases in fruit yield. Acceptable Brix values and fruit yields were achieved under early salinity treatment in Exp. 2, suggesting that increasing EC by 0.1 dS·m−1·day−1 at this time may be suitable for inducing moderate salinity stress.
In general, Brix values of later-harvested fruit in a truss are higher than those harvested earlier because later-harvested fruit have been subjected to a longer period of salinity stress. In our study, however, the opposite was the case, and this was also true in the controls. Plant growth is also typically decreased under salinity stress, but this was not observed in our study, suggesting that tomato plants had become acclimatized to salinity stress while nutrient solution EC was gradually increased. These results indicate that the mechanism of high-sugar tomato production under moderate salinity stress treatment may be different from that of currently practiced methods.
Consistent fruit quality in fruit trusses is a problem during the production of high-sugar tomatoes. In this study, fruit sugar concentrations were increased by salt stress, with about 6% Brix measured up to the fourth fruit on each truss. The fifth fruit had the lowest FW of fruits from salt-treated plants; its Brix was also low, similar to that of the control. This result suggests that the low sugar concentration of the fifth fruit was due to its low sink strength, i.e., the sink strength of earlier-maturing fruit in the truss might be higher than that of young fruit. Flowering uniformity within the truss was affected by low temperatures and short-day conditions at the seedling stage (Ohyama et al., 2005). Consequently, we believe that optimization of seedling conditions to ensure uniform flowering within the first flower truss is important for stabilization of sugar concentration and fruit yield in the first fruit truss.
Ascorbic acid changes in tomato fruits mirrored increases in sugar content. Our results are consistent with previous reports that ascorbic acid, which is synthesized from a sugar, increases along with fruit sugar (Sato et al., 2006; Zushi and Matsuzoe, 1998). Our results are also consistent with previous reports that titratable acid in tomato fruit subjected to salt treatment is higher than in controls (Sato et al., 2006; Saito et al., 2009). In our study, soluble solids contents in tomato fruit were significantly increased by salt treatment: from 5.1 to 5.9 g/100 g FW in late-treated plants in Exp. 1, and from 5.4 to 6.4 g/100 g FW in early-treated plants in Exp. 2. Saito et al. (2009) have reported that salinity stress promotes sucrose translocation into fruit and increases sucrose synthase activity. These results indicate that the moderate salinity stress applied in this study also accelerated the translocation of assimilate into tomato fruit.
In conclusion, the use of moderate salinity stress, thereby avoiding excessive stress on plants, increased sugar concentrations without decreasing fruit yield, and led to tomato fruits with average Brix of approximately 6%. Manipulation of nutrient solution concentrations based on cumulative temperature may be difficult, however, because of seasonal differences in plant growth rate. Further study is needed on year-round growth in single truss, high-density tomato production systems, as well as research into management indicators for moderate salinity stress based on both solar radiation and cumulative temperature.
Literature Cited
- Adams, P. and L. C. Ho. 1992. The susceptibility of modern tomato cultivars to blossom-end rot in relation to salinity. J. Hort. Sci. 67: 827–839.
- Araki, T., S. Watanabe, T. Wajima, M. Kitano, Y. Nakano and K. Okano. 2009. Short-term application of the concentrated deep seawater for production of high quality tomatoes by single-truss and high density cultivation. Environ. Control Biol. 47: 37–46.
- Belda, R. M. and L. C. Ho. 1993. Salinity effects on the network of vascular bundles during tomato fruit development. J. Hort. Sci. 68: 557–564.
- Belda, R. M., J. S. Fenlon and L. C. Ho. 1996. Salinity effects on the xylem vessels in tomato fruit among cultivars with different susceptibilities to blossom-end rot. J. Hort. Sci. 71: 173–179.
- Bolarin, M. C., M. T. Estan, M. Caro, R. Romero-Aranda and J. Cuartero. 2001. Relationship between tomato fruit growth and fruit osmotic potential under salinity. Plant Sci. 160: 1153–1159.
- Chretien, S., A. Gosselin and M. Dorais. 2000. High electrical conductivity and radiation-based water management improve fruit quality of greenhouse tomatoes grown in rockwool. HortScience 35: 627–631.
- Ehret, D. L. and L. C. Ho. 1986. The effects of salinity on dry matter partitioning and fruit growth in tomatoes grown in nutrient film culture. J. Hort. Sci. 61: 361–367.
- Ho, L. C., R. Belda, M. Brown, J. Andrews and P. Adams. 1993. Uptake and transport of calcium and the possible causes of blossom-end rot in tomato. J. Hort. Sci. 44: 509–518.
- Johkan, M., M. Ishii, T. Maruo, L. Na, S. Tsukagoshi, M. Hohjo, A. Nakaminami and Y. Shinohara. 2013. Improved light conditions at the fruit truss accelerate a harvest time and enhance an ascorbic acid concentration in a low-truss, high-density tomato production system. J. Japan. Soc. Hort. Sci. 82: 317–321.
- Kita, N. 1987. Growth analysis of semi-forced tomato plants. Bull. Kanagawa Hort. Exp. Sta. 34: 22–26 (In Japanese with English abstract).
- Li, Y. L., C. Stanghellini and H. Challa. 2001. Effect of electrical conductivity and transpiration on production of greenhouse tomato (Lycopersicon esculentum L.). Sci. Hortic. 88: 11–29.
- Ohyama, K., Y. Omura and T. Kozai. 2005. Effects of air temperature regimes on physiological disorders and floral development of tomato seedlings grown under continuous light. HortScience 40: 1304–1306.
- Okano, K., Y. Nakano, S. Watanabe and T. Ikdeda. 2002. Control of fruit quality by salinity stress at various fruit development stages of single-truss tomato grown in hydroponics. Environ. Control Biol. 40: 375–382 (In Japanese with English abstract).
- Saito, T., N. Fukuda, C. Matsukura and S. Nishimura. 2009. Effects of salinity on distribution of photosynthates and carbohydrate metabolism in tomato grown using nutrient film technique. J. Japan. Soc. Hort. Sci. 78: 90–96.
- Saito, T., N. Fukuda and S. Nishimura. 2006. Effects of salinity treatment duration and planting density on size and sugar content of hydroponically grown tomato fruits. J. Japan. Soc. Hort. Sci. 75: 392–398.
- Sakamoto, Y., S. Watanabe, T. Nakashima and K. Okano. 1999. Effects of salinity at two ripening stages on the fruit quality of single-truss tomato grown in hydroponics. J. Hort. Sci. Biotech. 74: 690–693.
- Sato, S., S. Sakaguchi, H. Furukawa and H. Ikeda. 2006. Effects of NaCl application to hydroponic nutrient solution on fruit characteristics of tomato (Lycopersicon esculentum Mill.). Sci. Hortic. 109: 248–253.
- Watanabe, S. 2006. New growing system for tomato with low node-order pinching and high density planting. Proceed. Vege. Tea Sci. 3: 91–98 (In Japanese).
- Wu, M. and C. Kubota. 2008. Effects of high electrical conductivity of nutrient solution and its application timing on lycopene, chlorophyll and sugar concentrations of hydroponic tomatoes during ripening. Sci. Hortic. 116: 122–129.
- Zushi, K. and N. Matsuzoe. 1998. Effect of soil water deficit on vitamin C, sugar, organic acid, amino acid and carotene contents of large-fruited tomatoes. J. Japan. Soc. Hort. Sci. 67: 927–933.