Abstract
The oxidation ditch process is one of the most economical approaches currently used to simultaneously remove organic carbon, nitrogen, and also phosphorus (P) from wastewater. However, limited information is available on biological P removal in this process. In the present study, microorganisms contributing to P removal in a full-scale oxidation ditch reactor were investigated using culture-dependent and -independent approaches. A microbial community analysis based on 16S rRNA gene sequencing revealed that a phylotype closely related to Dechloromonas spp. in the family Rhodocyclaceae dominated in the oxidation ditch reactor. This dominant Dechloromonas sp. was successfully isolated and subjected to fluorescent staining for polyphosphate, followed by microscopic observations and a spectrofluorometric analysis, which clearly demonstrated that the Dechloromonas isolate exhibited a strong ability to accumulate polyphosphate within its cells. These results indicate the potential key role of Dechloromonas spp. in efficient P removal in the oxidation ditch wastewater treatment process.
Nutrients such as organic carbon (C), nitrogen (N), and phosphorus (P) have to be removed from sewage/wastewater to avoid the eutrophication of aquatic water systems. Diverse types of wastewater treatment plants (WWTPs) that rely on microbial metabolic activities have been developed and utilized to achieve the efficient removal of nutrients. Aerobic treatment processes, such as activated sludge plants, are commonly utilized as sewage treatments. Aerobic processes are generally efficient for C removal, but not for N or P removal. Since the biological removal of N and P involves aerobic and anaerobic metabolism, conventional aerobic processes require additional reactors with different dissolved oxygen concentrations. The simultaneous removal of C, N, and P may be achieved in one reactor by alternating between aerobic and anaerobic phases. These processes are often referred to as simultaneous nitrification, denitrification, and phosphate removal (SNDPR) (54, 55).
Microbial metabolism related to N removal in WWTPs has been extensively examined (35). Ammonia-N in wastewater is primarily converted into nitrate (nitrification) under aerobic conditions by the combination of ammonium-oxidizing bacteria (e.g., Nitrosomonas spp.) and nitrite-oxidizing bacteria (e.g., Nitrospira spp.) or potentially by complete ammonia-oxidizing (comammox) bacteria (46). Nitrate is subsequently converted to N2 (denitrification) under anoxic conditions by diverse nitrate-reducing microorganisms. In contrast, studies on the microbiology of P removal are less comprehensive. Microbial P removal is generally performed by a group of microorganisms referred to as polyphosphate (polyP)-accumulating organisms (PAOs) (28, 53). PAOs have the ability to utilize intracellular polyP as an energy source during the anaerobic phase, during which they sequester available carbon sources. They then take up phosphate in wastewater and accumulate polyP in their cells under subsequent aerobic conditions, which results in efficient P removal from wastewater. The most well-known PAO group is Candidatus Accumulibacter phosphatis (hereafter called Accumulibacter spp.), classified into the family Rhodocyclaceae of class Betaproteobacteria, which has yet to be isolated in axenic cultures. Microbial community analyses based on 16S rRNA gene sequences, and also polyP staining combined with fluorescent in situ hybridization (FISH) and fluorescence-activated cell sorting (FACS) demonstrated that Accumulibacter spp. are the main contributors in diverse full-scale and lab-scale P-removing WWTPs (32, 52, 57, 58). In addition, a diverse range of species, including Actinobacteria, uncultured Halomonadaceae, and Rhodocyclaceae bacteria (not Accumulibacter spp.) have been identified as putative PAOs in full-scale SNDPR WWTPs (4, 22, 30, 31, 50). However, the paucity of pure-culture experiments has hampered furthering understanding on the ecophysiology of these microorganisms.
The oxidation ditch process is one of the most economical and efficient SNDPR techniques that simultaneously remove C, N, and P from actual sewage with repetitive aerobic/anaerobic treatment phases (7, 25, 36). Although biological C and N removal in oxidation ditch WWTPs has been extensively investigated (9, 12, 15), limited information is currently available on biological P removal. In the present study, we investigated microorganisms related to P removal in a full-scale oxidation ditch WWTP in Japan with culture-dependent and -independent approaches. Microbial community analyses based on 16S rRNA gene sequencing revealed that bacteria classified into the family Rhodocyclaceae, particularly Dechloromonas spp., predominated in the oxidation ditch WWTP. Some Rhodocyclaceae bacteria, including the dominant Dechloromonas strains, were isolated from the oxidation ditch WWTP, and their polyP accumulation abilities were examined in order to demonstrate their possible involvement in P removal from actual sewage.
Materials and Methods
Sample collection from the oxidation ditch WWTP
Sludge samples for the microbial community analysis and isolation of microorganisms were collected from the oxidation ditch WWTP in Okishima, Omihachiman, Shiga, Japan. This plant receives domestic wastewater with an annual average influent flow of 210 m3 d−1. The plant is generally operated with a repetitive cycle of 4-h aerobic and 6-h anaerobic phases. The aerobic (dissolved oxygen of >0.5 mg L−1) and anaerobic (dissolved oxygen of <0.5 mg L−1) phases are switched by the operation of mechanical aerators. Mixed liquor suspended solids are maintained at 5.0–8.0 g L−1. The annual averages of biological oxygen demand, total nitrogen, and total phosphorus in the influent/effluent are 167±93/5.0±1.2 mg L−1, 28.8±6.2/4.3±1.8 mg L−1, and 3.9±1.6/0.6±0.2 mg L−1, respectively, which suggests that the efficient removal of C, N, and P is achieved in the oxidation ditch WWTP. Sludge samples were recovered from the reactor during the aerobic phase.
Microbial community and phylogenetic analyses
Microbial genomic DNA was extracted from sludge samples using the FAST DNA Spin Kit for soil (MP Biomedicals, Santa Ana, CA, USA) according to the manufacturer’s instructions. The PCR amplification of 16S rRNA gene fragments for clone library analyses was conducted as described previously with a primer set of 27F and U533R (18, 19). PCR products were purified with a QIAquick PCR Purification Kit (QIAGEN, Hilden, Germany), ligated into the pGEM-T Easy Vector (Promega, Madison, WI, USA), and cloned into E. coli JM109 competent cells (Promega). The sequences of the cloned PCR products were elucidated at the Biomedical Center of TAKARA Bio.
The sequences obtained were assigned to phylotypes using BLASTClust (2) with a cut-off value of 97% sequence identity. Phylotypes were phylogenetically classified with the Classifier program in the Ribosomal Database Project (47) and were compared to sequences in the GenBank nucleotide sequence database using the BLAST program (2). Phylogenetic trees were constructed using the neighbor-joining method (37) with the program MEGA7 (24). Bootstrap resampling was conducted with 1,000 replicates to validate the robustness of the phylogenetic trees. The nucleotide sequence data obtained in this study have been submitted to the GenBank database under Accession Nos. LC145217–LC145287.
Isolation of Rhodocyclaceae bacteria
The isolation of Rhodocyclaceae bacteria was performed using an agar (1.5%)-solidified basal medium (pH 7.0) consisting of the following (L−1 of distilled water): 1 g of NH4Cl, 0.1 g of MgCl2·6H2O, 0.08 g of CaCl2·6H2O, 0.6 g of NaCl, 9.52 g of 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 0.1 g of yeast extract, and 10 mL each of vitamin solution and trace metal solution (19). Organic substrates (sodium acetate or sodium lactate, final concentration of 10 mM) and phosphate buffer solution (KH2PO4 and NaHPO4·12H2O, final concentration of 10 mM, pH 7.0) were separately sterilized and supplemented to the basal medium after autoclaving to avoid unintended reactions (43). Sludge samples were serially diluted and inoculated onto the medium and incubated at 25°C under aerobic conditions. After a 10-d incubation, colonies were randomly selected, and further purified with the same medium at least three times. The whole length of the 16S rRNA gene sequence was measured by direct sequencing of the DNA fragment derived from PCR amplification using the primer pair of 27F and 1492R as described previously (17).
Microscopic observations and quantification of microbial polyP accumulation
The isolated strains and E. coli K-12 (as a reference strain) were cultivated to the early stationary phases in the basal media described above and supplemented with 10 mM sodium lactate. In the microscopy analysis, 1 mL of cell suspension was collected by centrifugation (8,000×g, 5 min), resuspended in 1 mL of phosphate-buffered saline (PBS), and stained with 4′6-diamidino-2-phenylindole (DAPI) at a final concentration of 10 μg mL−1 for 30 min. After staining, cells were collected by centrifugation and resuspended in PBS. PolyP fluorescence and DNA fluorescence were observed using a fluorescence microscope (DMI 4000 B, Leica) with 340–380 nm excitation and 425 LP emission filters. In the spectrofluorometric polyP analysis, 1 mL of cell suspension was stained with DAPI following the same procedure used for microscopy. After staining and centrifugation, cells were resuspended in 200 μL PBS and loaded onto a 96-well flat-bottom microplate. Cells were excited at 355 nm, polyP fluorescence was captured at 535 nm emission, and DNA fluorescence was captured at 450 nm emission using bottom fluorescence reading on a SPECTRAmax Gemini XS (Molecular Devices, Tokyo, Japan) microplate reader. In the emission scan analysis, cells were excited at 355 nm and fluorescence emission was recorded between 425 nm and 595 nm at 5-nm intervals. All excitation and emission wavelengths had a bandwidth of 9 nm. Relative polyP fluorescence values were obtained by normalizing polyP fluorescence with DNA fluorescence (21). Fluorometric experiments to monitor DNA and polyP fluorescence were conducted in triplicate and the Student’s t-test was used for statistical analyses.
Results and Discussion
Microbial community analysis of the oxidation ditch WWTP
A clone library analysis based on the bacterial 16S rRNA gene was conducted in order to elucidate the microbial community structure in the full-scale oxidation ditch WWTP in Japan, in which the efficient removal of C, N, and P from domestic wastewater has been achieved. All phylotypes detected from the WWTP are listed in Table S1. Overall phylogenetic trends with phylum/class and family levels are shown in Fig. 1A and B, respectively. In the phylum/class level analysis (Fig. 1A), Betaproteobacteria (31.0% of all clones) was dominant, followed by Chloroflexi (11.1%), Alphaproteobacteria (10.3%), and Bacteroidetes (10.3%). The predominance of these phylogenetic groups is a common trait of SNDPR WWTPs including oxidation ditch systems (13, 15, 20, 39, 48, 49, 56).

In the family level analysis (Fig. 1B), the family Rhodocyclaceae (class Betaproteobacteria) was dominant (27.8% of all clones). Rhodocyclaceae bacteria are facultatively anaerobic, generally have nitrate-reducing abilities, and include some polyP-accumulating species (11, 23, 32), suggesting that Rhodocyclaceae bacteria contribute to the removal of C, N, and also P in the oxidation ditch WWTP. The results of further analyses on Rhodocyclaceae bacteria are presented in the next section.
Other dominant groups were the families Anaerolineaceae (phylum Chloroflexi) (10.3% of all clones), Planctomycetaceae (phylum Planctomycetes) (7.9%), Chitinophagaceae (phylum Bacteroidetes) (4.8%), and Nitrospiraceae (phylum Nitrospira) (4.8%). The family Anaerolineaceae includes strictly anaerobic bacteria that have the ability to degrade a wide range of organic compounds in various anaerobic environments, including anaerobic WWTPs (29, 51). Most members of Planctomycetaceae are strictly aerobic, chemoorganotrophic bacteria, and are often detected in aerobic WWTPs such as activated sludge plants (5, 41). Chitinophagaceae bacteria are aerobic or facultatively anaerobic, and generally have the ability to assimilate diverse sugars and amino acids (16). The family Nitrospiraceae as well as the family Nitrosomonadaceae (phylum Betaproteobacteria), which are aerobic nitrite- and ammonia-oxidizing bacterial groups (14), were dominant, suggesting that these bacteria contribute to the aerobic ammonia nitrification step in the oxidation ditch WWTP. Overall, the microbial community analysis revealed the co-existence of aerobic and anaerobic microorganisms in the oxidation ditch WWTP, which may be requisite for the simultaneous and efficient removal of C, N, and P.
Rhodocyclaceae phylotypes detected from the oxidation ditch WWTP
Since the family Rhodocyclaceae was the dominant bacterial group in the oxidation ditch WWTP, a detailed phylogenetic analysis was performed on the Rhodocyclaceae phylotypes. The phylogenetic positions of the Rhodocyclaceae phylotypes detected in the present study were analyzed by comparisons with sequences of representative Rhodocyclaceae strains in the databases (Fig. 2). The first and second most dominant phylotypes, Oki-11 and Oki-12 (20.6 and 2.4% of all clones, respectively), were closely related to Dechloromonas spp. (99.0% sequence identity to Dechloromonas aromatica strain RCB and 98.1% to Dechloromonas sp. MissR, respectively). Dechloromonas spp. are facultatively anaerobic, nitrate-reducing bacteria, and were originally isolated as aromatic compound-degrading bacteria (6). Dechloromonas spp. have frequently been detected in aerobic/anaerobic WWTPs (13, 20, 39, 49, 56), suggesting their contribution to the degradation of organic compounds under aerobic and anaerobic conditions and also to N removal via nitrate reduction. Furthermore, Dechloromonas spp. have been detected in P-removing WWTPs (10, 26, 45), suggesting their potential contribution to P removal. However, it remains controversial whether Dechloromonas spp. actually function as PAOs. The results of the genome analysis showed that D. aromatica strain RCB harbored genes required for polyP accumulation, including polyphosphate kinase and exopolyphosphatase (38). Furthermore, polyP staining coupled with FISH (23) and FACS (11) revealed that Dechloromonas spp. have the ability to store polyP in full-scale WWTPs. In contrast, some research groups reported opposite findings (1, 27, 40); Dechloromonas-related bacteria that dominated in P-removal WWTPs did not accumulate polyP in situ. Furthermore, there have been no studies on the polyP-accumulating abilities of Dechloromonas spp. using pure-culture experiments with isolated strains.

The other Rhodocyclaceae phylotypes (Oki-13 to -18, Fig. 2 and Supplementary Table S1) were also detected in the oxidation ditch WWTP. It is important to note that two phylotypes (Oki-16 and -17) are related to Candidatus Accumulibacter phosphatis (sequence identities of 96.9 and 93.0%, respectively), suggesting their ability to accumulate polyP. However, these phylotypes only comprise a minor part of the population, indicating that their contribution to P removal in the oxidation ditch WWTP is marginal.
Isolation of Rhodocyclaceae bacteria from the oxidation ditch WWTP
The isolation of Rhodocyclaceae bacteria (particularly the dominant Dechloromonas phylotypes) was undertaken in order to investigate their physiology using pure-culture experiments. Based on previous findings showing that most Rhodocyclaceae bacteria prefer short chain organic acids as growth substrates (32), media supplemented with acetate or lactate were utilized for isolation. Colonies appearing on plate media were purified, and the partial sequences of the 16S rRNA gene were obtained. Among the 290 isolates tested, 121 isolates were classified into the family Rhodocyclaceae. Among them, 43 isolates were closely related to Dechloromonas spp. with 98–100% sequence identities, and some of them showed identical sequences to the dominant phylotype Oki-11 (e.g., the isolate LOPT-10–19 in Fig. 2). Furthermore, some isolates closely related to the other genera of Rhodocyclaceae were also obtained. Four representative Rhodocyclaceae isolates classified into different genera were subjected to further phylogenetic analyses by sequencing the whole length of the 16S rRNA gene (Fig. 2). These four strains, namely Dechloromonas sp. LOPT-10–19, Zoogloea sp. AOPS-03–15, Thauera sp. LOPT-10–14, and Dechlorosoma sp. ANPS-21–24, were subjected to physiological analyses.
PolyP-accumulating abilities of Rhodocyclaceae isolates
The ability to accumulate polyP by Rhodocyclaceae isolates was assessed through fluorescence staining approaches. DAPI is a fluorescent chemical that is widely used to visualize polyP stored in microbial cells (42, 44). DAPI stains DNA in addition to polyP, but with distinct fluorescence emission (blue and yellow-green, respectively) when excited with UV light (3, 33). The representative Rhodocyclaceae isolates and E. coli were aerobically cultivated in basal medium supplemented with lactate. Cells in the stationary phases were collected, stained with DAPI, and subjected to fluorescence microscopy (Fig. 3). Yellow-green fluorescence was not observed in DAPI-stained E. coli cells (Fig. 3F). In contrast, almost all cells of Dechloromonas sp. LOPT-10–19 exhibited yellow-green fluorescence derived from polyP granules (Fig. 3G). A previous study reported that the numbers and cellular localization of polyP granules differ between PAOs, ranging from a single large granule at one end of a cell to multiple granules scattered within a cell (8). The accumulation of polyP at polar ends of the rod-shaped cell in Dechloromonas sp. LOPT-10–19 is very similar to the accumulation patterns observed for Corynebacterium (34); however, the mechanisms underlying particular accumulation patterns are currently unknown. The other Rhodocyclaceae isolates also exhibited yellow-green fluorescence, while the manner of polyP accumulation was dissimilar to that of Dechloromonas sp. LOPT-10–19. Regarding Zoogloea sp. AOPS-03–15 and Dechlorosoma sp. ANPS-21–24, only minor fractions of cells exhibited yellow-green fluorescence (Fig. 3H and J). As for Thauera sp. LOPT-10–14 (Fig. 3I), while most cells exhibited yellow-green fluorescence, the intensity and size of polyP granules relative to the cell area appeared to be smaller than those of Dechloromonas sp. LOPT-10–19. Thauera sp. LOPT-10–14 has a larger cell size (~3–5 μm in length and ~2 μm in width) than Dechloromonas sp. LOPT-10–19 (~1.5–3 μm in length and ~0.8 μm in width). However, polyP granules were of a similar size for both strains (~0.5–0.8 μm in diameter).

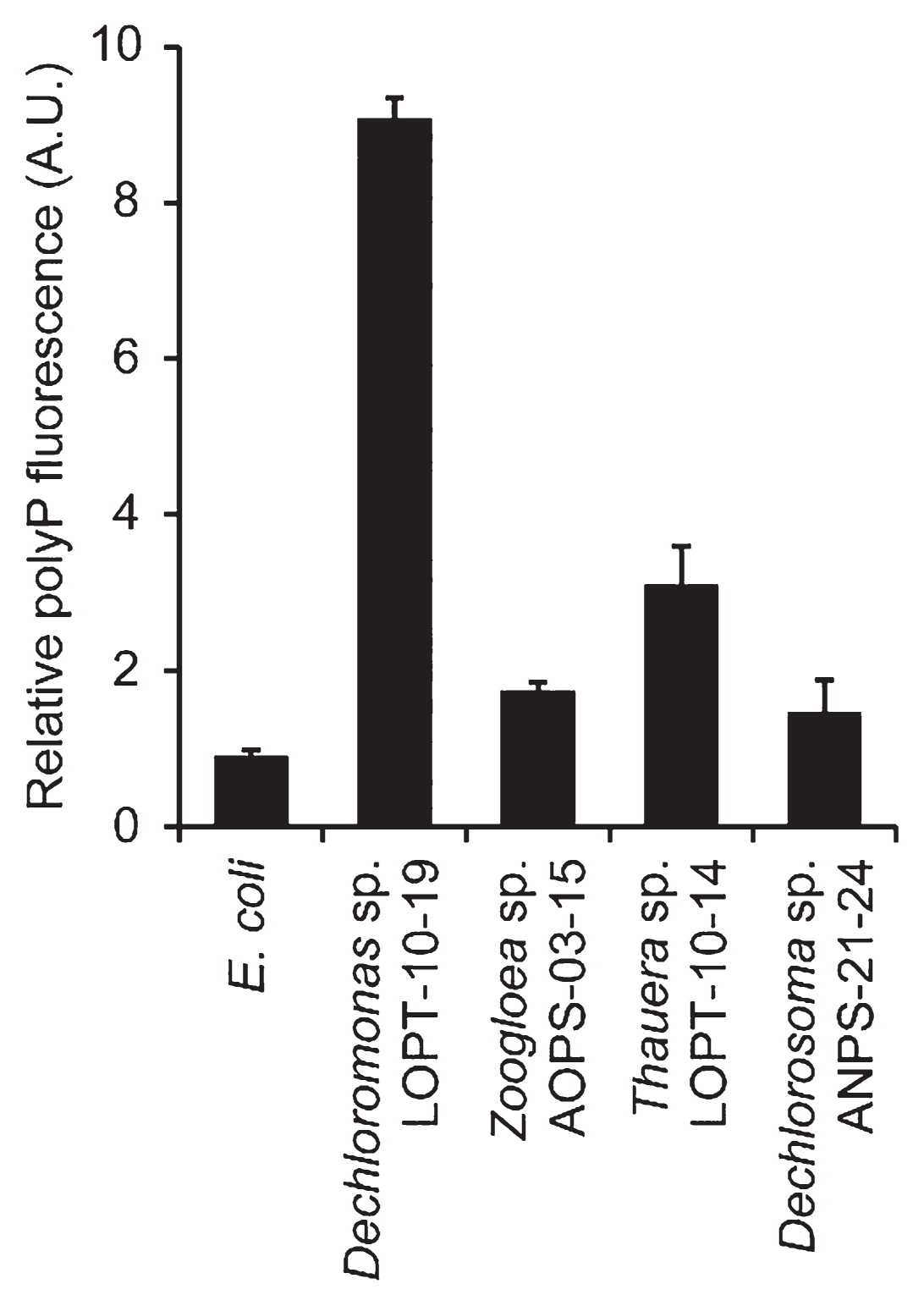
In order to quantitatively compare polyP-accumulating activities, DAPI-stained cell suspensions were subjected to spectrofluorometry. The relative amounts of polyP stored in cells were assessed by normalizing polyP-derived fluorescence (535 nm emission) using DNA-derived fluorescence (450 nm emission) (Fig. 4 and Supplementary Fig. S1). Relative polyP fluorescence from Dechloromonas sp. LOPT-10–19 was more than 10-fold higher than the background levels observed in the reference strain (E. coli K-12). The other Rhodococcaceae strains also exhibited significantly higher polyP fluorescence than E. coli; however, it was still markedly less than that of Dechloromonas sp. LOPT-10–19. These results were consistent with the microscopy images (Fig. 3), in which Zoogloea sp. AOPS-03–15 and Dechlorosoma sp. ANPS-21–24 both had fewer polyP granules than Dechloromonas sp. LOPT-10–19, and, as discussed above, Thauera sp. LOPT-10–14 had larger cells, making relative polyP fluorescence lower. The results of these microscopic and fluorometric analyses demonstrated that Dechloromonas spp. dominantly detected in the oxidation ditch WWTP have strong abilities to intracellularly accumulate polyP. This study is the first to demonstrate significant polyP accumulation by Dechloromonas spp. in pure-culture experiments, and provides an insight into the contribution of Dechloromonas spp. to microbial P removal in diverse WWTPs. The future challenge is to understand the role of Dechloromonas spp. in P removal in the WWTP, e.g., collecting polyP-accumulating cells from in situ microbial communities using DAPI staining combined with flow cytometry, followed by phylogenetic identification. By carefully following polyP accumulation in Dechloromonas spp. in the WWTP, P removal activity within the greater microbial community may be more clearly understood.
Conclusion
In the present study, we demonstrated the dominance of Rhodocyclaceae bacteria, particularly Dechloromonas spp., in the full-scale oxidation ditch WWTP in Japan, in which C, N, and also P are stably and efficiently removed. Some strains closely related to Dechloromonas spp. were successfully isolated and proven to exhibit strong abilities to intracellularly accumulate polyP. This is the first study to demonstrate polyP accumulation by Dechloromonas spp. in pure-culture experiments. These results suggest that Dechloromonas spp. is an important player in the removal of C, N, and also P in the oxidation ditch WWTP.
Acknowledgements
The sludge samples and operation information of the oxidation ditch WWTP were kindly provided by Hiyoshi Corporation. We thank Hiromi Ikebuchi and Asuka Tanaka for their technical assistance. This research was partly supported by the Institute of Fermentation, Osaka. The authors declare no conflict of interest.
References
- 1. Ahn, J., S. Schroeder, M. Beer, S. McIlroy, R.C. Bayly, J.W. May, G. Vasiliadis, and R.J. Seviour. 2007. Ecology of the microbial community removing phosphate from wastewater under continuously aerobic conditions in a sequencing batch reactor. Appl Environ Microbiol. 73:2257-2270.
- 2. Altschul, S.F., T.L. Madden, A.A. Schäffer, J. Zhang, Z. Zhang, W. Miller, and D.J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389-3402.
- 3. Aschar-Sobbi, R., A.Y. Abramov, C. Diao, M.E. Kargacin, G.J. Kargacin, R.J. French, and E. Pavlov. 2008. High sensitivity, quantitative measurements of polyphosphate using a new DAPI-based approach. J Fluoresc. 18:859-866.
- 4. Beer, M., and H. Stratton. 2006. Which are the polyphosphate accumulating organisms in full-scale activated sludge enhanced biological phosphate removal systems in Australia? J Appl Microbiol. 100:233-243.
- 5. Chouari, R., D. Le Paslier, P. Daegelen, P. Ginestet, J. Weissenbach, and A. Sghir. 2003. Molecular evidence for novel planctomycete diversity in a municipal wastewater treatment plant. Appl Environ Microbiol. 69:7354-7363.
- 6. Coates, J.D., R. Chakraborty, J.G. Lack, S.M. O’Connor, K.A. Cole, K.S. Bender, and L.A. Achenbach. 2001. Anaerobic benzene oxidation coupled to nitrate reduction in pure culture by two strains of Dechloromonas. Nature. 411:1039-1043.
- 7. Daigger, G., and H. Littleton. 2000. Characterization of simultaneous nutrient removal in staged, closed-loop bioreactors. Wat Environ Res. 72:330-339.
- 8. Docampo, R. 2006. Acidocalcisomes and polyphosphate granules, p.53-70. In J.M. Shively (ed.), Inclusions in Prokaryotes, Microbiology Monographs. Springer, Berlin, Heidelberg.
- 9. Gao, S.Y., Y.Z. Peng, S.Y. Wang, and J. Yan. 2006. Novel strategy of nitrogen removal from domestic wastewater using pilot Orbal oxidation ditch. J Environ Sci. 18:833-839.
- 10. Goel, R., P. Sanhueza, and D. Noguera. 2005. Evidence of Dechloromonas sp. participating in enhanced biological phosphorous removal (EBPR) in a bench-scale aerated-anoxic reactor. Proc Water Environ Fed. 41:3864-3871.
- 11. Günther, S., M. Trutnau, S. Kleinsteuber, G. Hause, T. Bley, I. Röske, H. Harms, and S. Müller. 2009. Dynamics of polyphosphate-accumulating bacteria in wastewater treatment plant microbial communities detected via DAPI (4′,6′-diamidino-2-phenylindole) and tetracycline labeling. Appl Environ Microbiol. 75:2111-2121.
- 12. Guo, C.Z., W. Fu, X.M. Chen, D.C. Peng, and P.K. Jin. 2013. Nitrogen-removal performance and community structure of nitrifying bacteria under different aeration modes in an oxidation ditch. Water Res. 47:3845-3853.
- 13. Hu, M., X. Wang, X. Wen, and Y. Xia. 2012. Microbial community structures in different wastewater treatment plants as revealed by 454-pyrosequencing analysis. Bioresour Technol. 117:72-79.
- 14. Isobe, K., and N. Ohte. 2014. Ecological perspectives on microbes involved in N-cycling. Microbes Environ. 29:4-16.
- 15. Jin, P., X. Wang, X. Wang, H.H. Ngo, and X. Jin. 2015. A new step aeration approach towards the improvement of nitrogen removal in a full scale Carrousel oxidation ditch. Bioresour Technol. 198:23-30.
- 16. Kämpfer, P., N. Lodders, and E. Falsen. 2011. Hydrotalea flava gen. nov., sp. nov., a new member of the phylum Bacteroidetes and allocation of the genera Chitinophaga, Sediminibacterium, Lacibacter, Flavihumibacter, Flavisolibacter, Niabella, Niastella, Segetibacter, Parasegetibacter, Terrimonas, Ferruginibacter, Filimonas and Hydrotalea to the family Chitinophagaceae fam. nov. Int J Syst Evol Microbiol. 61:518-523.
- 17. Kato, S., S. Haruta, Z.J. Cui, M. Ishii, A. Yokota, and Y. Igarashi. 2004. Clostridium straminisolvens sp. nov., a moderately thermophilic, aerotolerant and cellulolytic bacterium isolated from a cellulose-degrading bacterial community. Int J Syst Evol Microbiol. 54:2043-2047.
- 18. Kato, S., R. Nakamura, F. Kai, K. Watanabe, and K. Hashimoto. 2010. Respiratory interactions of soil bacteria with (semi)conductive iron-oxide minerals. Environ Microbiol. 12:3114-3123.
- 19. Kato, S., E. Goya, M. Tanaka, W. Kitagawa, Y. Kikuchi, K. Asano, and Y. Kamagata. 2016. Enrichment and isolation of Flavobacterium strains with tolerance to high concentrations of cesium ion. Sci Rep. 6:20041.
- 20. Kim, B.C., S. Kim, T. Shin, H. Kim, and B.I. Sang. 2013. Comparison of the bacterial communities in anaerobic, anoxic, and oxic chambers of a pilot A2O process using pyrosequencing analysis. Curr Microbiol. 66:555-565.
- 21. Klauth, P., S.R. Pallerla, D. Vidaurre, C. Ralfs, V.F. Wendisch, and S.M. Schoberth. 2006. Determination of soluble and granular inorganic polyphosphate in Corynebacterium glutamicum. Appl Microbiol Biotechnol. 72:1099-1106.
- 22. Kong, Y., J.L. Nielsen, P. Halkjær, and P.H. Nielsen. 2005. Identity and ecophysiology of uncultured actinobacterial polyphosphate-accumulating organisms in full-scale enhanced biological phosphorus removal plants. Appl Environ Microbiol. 71:4076-4085.
- 23. Kong, Y., Y. Xia, J.L. Nielsen, and P.H. Nielsen. 2007. Structure and function of the microbial community in a full-scale enhanced biological phosphorus removal plant. Microbiology. 153:4061-4073.
- 24. Kumar, S., G. Stecher, and K. Tamura. 2016. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol. 33:1870-1874.
- 25. Littleton, H., G. Daigger, and P. Strom. 2007. Application of computational fluid dynamics to closed-loop bioreactors: II. Simulation of biological phosphorus removal using computational fluid dynamics. Wat Environ Res. 79:613-624.
- 26. Liu, Y., T. Zhang, and H.H. Fang. 2005. Microbial community analysis and performance of a phosphate-removing activated sludge. Bioresour Technol. 96:1205-1214.
- 27. McIlroy, S.J., A. Starnawska, P. Starnawski, A.M. Saunders, M. Nierychlo, P.H. Nielsen, and J.L. Nielsen. 2016. Identification of active denitrifiers in full-scale nutrient removal wastewater treatment systems. Environ Microbiol. 18:50-64.
- 28. Mino, T., M.C.M. Van Loosdrecht, and J.J. Heijnen. 1998. Microbiology and biochemistry of the enhanced biological phosphate removal process. Water Res. 32:3193-3207.
- 29. Narihiro, T., T. Terada, A. Ohashi, Y. Kamagata, K. Nakamura, and Y. Sekiguchi. 2012. Quantitative detection of previously characterized syntrophic bacteria in anaerobic wastewater treatment systems by sequence-specific rRNA cleavage method. Water Res. 46:2167-2175.
- 30. Nguyen, H.T.T., J.L. Nielsen, and P.H. Nielsen. 2012. ‘Candidatus Halomonas phosphatis’, a novel polyphosphate-accumulating organism in full-scale enhanced biological phosphorus removal plants. Environ Microbiol. 14:2826-2837.
- 31. Nielsen, P.H., A.T. Mielczarek, C. Kragelund, J.L. Nielsen, A.M. Saunders, Y. Kong, A.A. Hansen, and J. Vollertsen. 2010. A conceptual ecosystem model of microbial communities in enhanced biological phosphorus removal plants. Water Res. 44:5070-5088.
- 32. Oehmen, A., G. Carvalho, C.M. Lopez-Vazquez, M.C.M. van Loosdrecht, and M.A.M. Reis. 2010. Incorporating microbial ecology into the metabolic modelling of polyphosphate accumulating organisms and glycogen accumulating organisms. Water Res. 44:4992-5004.
- 33. Omelon, S., J. Georgiou, Z.J. Henneman, L.M. Wise, B. Sukhu, T. Hunt, C. Wynnyckyj, D. Holmyard, R. Bielecki, and M.D. Grynpas. 2009. Control of vertebrate skeletal mineralization by polyphosphates. PLoS One. 4:e5634.
- 34. Pallerla, S.R., S. Knebel, T. Polen, P. Klauth, J. Hollender, V.F. Wendisch, and S.M. Schoberth. 2005. Formation of volutin granules in Corynebacterium glutamicum. FEMS Microbiol Lett. 243:133-140.
- 35. Peng, Y., and G. Zhu. 2006. Biological nitrogen removal with nitrification and denitrification via nitrite pathway. Appl Microbiol Biotechnol. 73:15-26.
- 36. Peng, Y., H. Hou, S. Wang, Y. Cui, and Y. Zhiguo. 2008. Nitrogen and phosphorus removal in pilot-scale anaerobic-anoxic oxidation ditch system. J Environ Sci. 20:398-403.
- 37. Saitou, N., and M. Nei. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 4:406-425.
- 38. Salinero, K.K., K. Keller, W.S. Feil, H. Feil, S. Trong, G. Di Bartolo, and A. Lapidus. 2009. Metabolic analysis of the soil microbe Dechloromonas aromatica str. RCB: indications of a surprisingly complex life-style and cryptic anaerobic pathways for aromatic degradation. BMC Genomics. 10:351.
- 39. Saunders, A.M., M. Albertsen, J. Vollesen, and P.H. Nielsen. 2016. The activated sludge ecosystem contains a core community of abundant organisms. ISME J. 10:11-20.
- 40. Schroeder, S., J. Ahn, and R.J. Seviour. 2008. Ecophysiology of polyphosphate-accumulating organisms and glycogen-accumulating organisms in a continuously aerated enhanced biological phosphorus removal process. J Appl Microbiol. 105:1412-1420.
- 41. Snaidr, J., R. Amann, I. Huber, W. Ludwig, and K.H. Schleifer. 1997. Phylogenetic analysis and in situ identification of bacteria in activated sludge. Appl Environ Microbiol. 63:2884-2896.
- 42. Streichan, M., J.R. Golecki, and G. Schö1990Polyphosphate accumulating bacteria from sewage plants with different processes for biological phosphorus removal. FEMS Microbiol Ecol. 73:113-124.
- 43. Tanaka, T., K. Kawasaki, S. Daimon, W. Kitagawa, K. Yamamoto, H. Tamaki, M. Tanaka, C.H. Nakatsu, and Y. Kamagata. 2014. A hidden pitfall in the preparation of agar media undermines microorganism cultivability. Appl Environ Microbiol. 80:7659-7666.
- 44. Tijssen, J.P.F., H.W. Beekes, and J. Van Steveninck. 1982. Localization of polyphosphates in Saccharomyces fragilis, as revealed by 4′6-diamidino-2-phenylindole fluorescence. Biochim Biophys Acta. 721:394-398.
- 45. Tu, Y., and A.J. Schuler. 2013. Low acetate concentrations favor polyphosphate-accumulating organisms over glycogen-accumulating organisms in enhanced biological phosphorus removal from wastewater. Environ Sci Technol. 47:3816-3824.
- 46. van Kessel, M.A., D.R. Speth, M. Albertsen, P.H. Nielsen, H.J. Op den Camp, B. Kartal, M.S. Jetten, and S. Lücker. 2015. Complete nitrification by a single microorganism. Nature. 528:555-559.
- 47. Wang, Q., G.M. Garrity, J.M. Tiedje, and J.R. Cole. 2007. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 73:5261-5267.
- 48. Wang, X., M. Hu, Y. Xia, X. Wen, and K. Ding. 2012. Pyrosequencing analysis of bacterial diversity in 14 wastewater treatment systems in China. Appl Environ Microbiol. 78:7042-7047.
- 49. Wen, Y., Y. Jin, J. Wang, and L. Cai. 2015. MiSeq sequencing analysis of bacterial community structures in wastewater treatment plants. Pol J Environ Stud. 24:1809-1815.
- 50. Wong, M.T., T. Mino, R.J. Seviour, M. Onuki, and W.T. Liu. 2005. In situ identification and characterization of the microbial community structure of full-scale enhanced biological phosphorous removal plants in Japan. Water Res. 39:2901-2914.
- 51. Yamada, T., Y. Sekiguchi, S. Hanada, H. Imachi, A. Ohashi, H. Harada, and Y. Kamagata. 2006. Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov. and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes, and description of the new classes Anaerolineae classis nov. and Caldilineae classis nov. in the bacterial phylum Chloroflexi. Int J Syst Evol Microbiol. 56:1331-1340.
- 52. Yilmaz, G., R. Lemaire, J. Keller, and Z. Yuan. 2008. Simultaneous nitrification, denitrification, and phosphorus removal from nutrient-rich industrial wastewater using granular sludge. Biotechnol Bioeng. 100:529-541.
- 53. Yuan, Z., S. Pratt, and D.J. Batstone. 2012. Phosphorus recovery from wastewater through microbial processes. Curr Opin Biotechnol. 23:878-883.
- 54. Zeng, R.J., A.M. Saunders, Z.G. Yuan, L.L. Blackall, and J. Keller. 2003. Identification and comparison of aerobic and denitrifying polyphosphate-accumulating organisms. Biotechnol Bioeng. 83:140-148.
- 55. Zeng, R.J., R. Lemaire, Z. Yuan, and J. Keller. 2003. Simultaneous nitrification, denitrification, and phosphorus removal in a lab-scale sequencing batch reactor. Biotechnol Bioeng. 84:170-178.
- 56. Zhang, T., M.F. Shao, and L. Ye. 2012. 454 pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J. 6:1137-1147.
- 57. Zilles, J.L., J. Peccia, M. Kim, C. Hung, and D.R. Noguera. 2002. Involvement of Rhodocyclus-related organisms in phosphorus removal in full-scale wastewater treatment plants. Appl Environ Microbiol. 68:2763-2769.
- 58. Zilles, J.L., C.H. Hung, and D.R. Noguera. 2002. Presence of Rhodocyclus in a full-scale wastewater treatment plant and their participation in enhanced biological phosphorus removal. Water Sci Technol. 46:123-128.
