Abstract
Branched nonylphenol (BNP), a degradation product of nonylphenol polyethoxylates, exerts estrogenic effects on various organisms. The genes underlying BNP degradation by Sphingobium amiense DSM 16289T were analyzed by complete genome sequencing and compared with those of the versatile BNP-degrading Sphingobium cloacae JCM 10874T. An opdA homolog (opdADSM16289) encoding BNP degradation activity was identified in DSM 16289T, in contrast with JCM 10874T, possessing both the opdA homolog and nmoA. The degradation profile of different BNP isomers was examined by Escherichia coli transformants harboring opdADSM16289, opdAJCM10874, and nmoAJCM10874 to characterize and compare the expression activities of these genes.
Branched nonylphenol (BNP) is a persistent compound known to exert estrogenic effects in aquatic organisms, mammals, and birds (23, 30, 37). The environmental occurrence of BNP is mainly due to the biodegradation of nonylphenol polyethoxylates (NPEOx), which are non-ionic surfactants used widely in industrial and agricultural applications (16, 24). This estrogenic activity is found in the shorter homologs of NPEOx, as well as BNP, but not in the higher homologs. Bacteria that degrade NPEOx are known to be ubiquitous in various environments (2, 16, 24). Commercial NPEOx is produced by the ethoxylation of technical-grade BNP, which is composed of at least 22 para-isomers possessing different degrees of branching in their alkyl side chain (36), and, thus, environmentally occurring BNP is recalcitrant to biodegradation. Previous studies demonstrated that BNP isomers have different biodegradabilities and estrogenicities in the environment (for a review, see ref. 23). The isolation of a bacterial strain capable of utilizing BNP as the sole carbon and energy source was pioneered by Tanghe et al. (32) in 1999, and the strain was designated as Sphingomonas sp. TTNP3. Following that study, several BNP-degrading bacteria were isolated, most of which were characterized phylogenetically as members of the Sphingomonas/Sphingobium group: Sphingobium cloacae S-3T (=JCM 10874T) (7), Sphingobium amiense YTT (=DSM 16289T) (6, 34), Sphingobium xenophagum Bayram (8), and Sphingomonas sp. NP5 (31). Among these BNP-degrading bacteria, the strains TTNP3 and Bayram were studied in-depth for their BNP degradation pathway, and the initial step of degradation was found to be a type II ipso substitution mechanism (5, 9, 10, 18). In this mechanism, BNP is oxidized at the aromatic carbon atom, followed by substitution with an alkyl side chain, resulting in separation to a hydroquinone and carbocation with nine carbon atoms. The enzymes involved in the type II ipso substitution of BNP and BNP-related octylphenol (OP) were identified as the flavin-dependent monooxygenase (14) octylphenol 4-monooxygenase (OpdA), which was initially identified in OP-degrading Sphingomonas sp. PWE1 (27). This enzyme was subsequently found in the BNP-degrading strains TTNP3 and Bayram (28), suggesting that OpdA is also responsible for BNP degradation. Thereafter, it was revealed that BNP-degrading Sphingomonas sp. NP5 had two identical nonylphenol monooxygenase genes (nmoA) and their predicted protein sequence showed 83% identity to the OpdA of Sphingomonas sp. PWE1 (31).
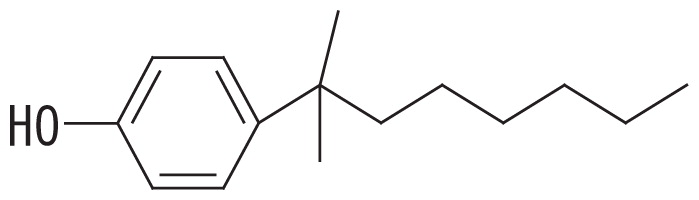
A high-resolution gas chromatographic analysis of BNP achieved the resolution of 18 para-isomers, which were classified into five distinct groups (36). By using this method, we previously demonstrated that BNP isomers with the α-dimethyl, α-methyl and α-ethyl structures (groups 1 and 2, respectively, in the left side columns of Table 1) were rapidly degraded by strain DSM 16289T (initial degradation rate, approx. 0.43 h−1 for 10 mg L−1 BNP), while the breakdown of isomers with highly branched chains (groups 3–5 in Table 1) was markedly less efficient (15). In contrast, strain JCM 10874T showed almost no substrate preference and degraded all the BNP isomers at similar rates (initial degradation rate, approx. 1.04 h−1 for 10 mg L−1 BNP). Prior to this study, we identified an opdA homolog and the nmoA gene in the two separate circular plasmids of strain JCM 10874T using a genome analysis (26). In the present study, to clarify the genetic background of BNP degradation by the strain DSM 16289T, we elucidated its complete genome sequence and identified an opdA homolog in a circular plasmid. Furthermore, to clarify differences in their preferences for BNP isomers, we compared the degradation profile of BNP isomers by three Escherichia coli transformants harboring the opdA homologs of the two strains and the nmoA gene of strain JCM 10874T.
Table 1
Degradation profiles of branched nonylphenol isomer groups by
Escherichia coli JM109 strains harboring pSA
opdA, pSC
opdA, and pSC
nmoA, constructed from
Sphingobium amiense DSM 16289
T (pSA
opdA) and
Sphingobium cloacae JCM 10874
T (pSC
opdA and pSC
nmoA).
| No. |
Representative structure of BNP isomer group |
Degradation ratio/E. coli transformant with |
|
| pSAopdA |
pSCopdA |
pSCnmoA |
| 1 |
 |
1.00 (0.10)a |
1.00 (0.04)a |
1.00 (0.04)a |
| 2 |
 |
0.88 (0.14)a |
0.73 (0.06)b |
0.86 (0.04)ab |
| 3 |
 |
0.33 (0.08)b |
0.65 (0.07)b |
1.02 (0.08)a |
| 4 |
 |
0.24 (0.03)b |
0.44 (0.02)c |
0.77 (0.19)ab |
| 5 |
 |
0.20 (0.14)b |
0.46 (0.06)c |
0.62 (0.07)b |
Fourteen BNP isomers were separated into five groups by the GC-MS analysis: group 1, isomers with the α-dimethyl structure; group 2, isomers with the α-methyl, α-ethyl structure; group 3, isomers with the α-methyl, β-methyl structure; group 4, isomers with the α-methyl, β-ethyl structure; and group 5, isomers with the α-methyl, α-propyl structure (15). The degradation ratio is defined as the normalized ratio of the degradation rate for each BNP isomer group to that for isomer group 1 after a 24-h incubation. These values are the means of triplicate runs and standard deviation values are shown in the brackets. Mean values in the same column with common letters are not significantly different from each other according to a one-way ANOVA with Tukey’s HSD test, p<0.05. The percent degradation rates of isomer group 1 by the transformants harboring pSAopdA, pSCopdA, and pSCnmoA were 34, 62, and 48%, respectively. The commercial BNP product used in this study, in groups 1 to 5, contained 56.2, 28.0, 7.2, 3.0, and 5.6% BNP, respectively.
The strains DSM 16289T and JCM 10874T were obtained from the DSMZ culture collection in Germany and RIKEN BRC-JCM in Japan, respectively. These strains were grown at 30°C in two-fold (strain DSM 16289T) and five-fold (strain JCM 10874T) diluted nutrient broth (pH 7.2). Nutrient broth comprised 1.0% (w/v) Bonito extract (Wako Pure Chemical Industry, Osaka, Japan), 1.0% (w/v) Bacto Peptone (Beckton, Dickinson and Company, Sparks, MD, USA), and 0.5% (w/v) NaCl. Genomic DNA was extracted from the cultured cells as described previously (25), and the sequencing of genomic DNA from the strain DSM 16289T was performed using PacBio RS II and the HiSeq 2500 instrument at GeneBay, Yokohama, Japan (http://genebay.co.jp). The hybrid assembly with PacBio RSII and HiSeq 2500 was performed by SPAdes v.3.10.1 (4), Unicycler v.0.4.0 (38), and Canu v.1.5 (20). Genome sequences were annotated using the Microbial Genome Annotation Pipeline (MiGAP: https://www.migap.org), followed by manual annotation with the NCBI-nr databases using the BLASTP program (3). tRNA was predicted using tRNAscan-SE 2.0 (22), and GC contents were calculated using GC-Profil (11). Multiple sequence alignments and a phylogenetic analysis using the UPGMA method were performed with Clustal W (33) and MEGA v.7.0.26 (21), respectively.
The opdA homologs of the strains DSM 16289T and JCM 10874T were amplified by PCR using the opdA-forward primer (5′-TTC ATC CTG AAA GAC ACT GCC GGA-3′) and opdA-reverse primer (5′-ACG CGC TTC CAG ACC AAC CTA TTT-3′) (27). The primers for nmoA gene amplification were originally designed from the opdA sequence (27) as forward: 5′-ATC CAA CGC GAA CAA CTT CGT GCA-3′ (nmoA-F) and reverse: 5′-TCA TTA TGG CTA GCG CGC CTA CTT-3′ (nmoA-R). PCR was performed using KOD-Plus-Neo (TOYOBO, Osaka, Japan) under the following conditions: 94°C for 2 min and 25 cycles of 98°C for 10 s, 72°C for 30 s, and 68°C for 50 s. Amplicon products were subcloned into pGEM-T Easy plasmid vectors (Promega, Madison, WI, USA), and the resulting plasmids were transformed into E. coli JM 109 (Takara Bio, Shiga, Japan). The constructed plasmids were designated pSAopdA, pSCopdA, and pSCnmoA for the opdA homologs of strains DSM 16289T and JCM 10874T and the nmoA gene of strain JCM 10874T, respectively. The transformants were grown on agar plates of LB (29) medium containing 100 μg mL−1 ampicillin (LB/Amp) and 200 mg L−1 BNP (Kanto Chemical, Tokyo, Japan). Aliquots of 100 mM IPTG aqueous solution (100 μL) and 40 mg mL−1 X-gal (5-bromo-4-chloro-3-indolyl-d-galactoside; 25 μL) were spread on the plate surface for blue/white colony selection. In addition to this selection, colonies accumulating reddish brown compounds were selected as BNP degraders according to the results of BNP-degrading Sphingomonas sp. NP5 (31). Selected transformants were examined to confirm the presence of the cloned sequence in the plasmid. In the assay of the BNP isomer degradation profile, E. coli JM 109 transformants were grown in LB/Amp liquid medium at 37°C for 24 h. E. coli cells were collected by centrifugation, suspended in fresh LB/Amp at OD600 of 0.5, and distributed into a series of sterile, glass test tubes (Φ13 mm by 100 mm) in 2-mL portions. After 10 μL of BNP stock solution (5 g L−1) in acetone was added to each of the tubes (at a final concentration of 25 mg L−1), the cultures were incubated at 37°C with gentle shaking. Triplicate cultures were assayed for the residual BNP before and after 24 h of incubation. A 2-mL culture was mixed with 1 mL of salting out solution (0.5 M HCl saturated with NaCl) (6) and 2 mL of hexane containing 10 mg L−1 4-tert-OP (Sigma-Aldrich, MO, USA) as the internal standard. After vigorous shaking, the resulting hexane layer was collected and used for BNP determination by gas chromatography–mass spectrometry (GC-MS). E. coli JM109 transformed with a plasmid containing no insert was grown as the reference to estimate the recovery of BNP from the grown culture. The recovery of each BNP group (mean±SD, n=3) relative to before the incubation ranged between 74±3% (BNP group 5) and 83±2% (BNP group 1) for the cultures grown for 24 h. The remaining BNP did not appear to be extractable by hexane, possibly due to its lipophilic property and tight binding to bacterial cells (1), and, thus, the calculation of the degradation rate was corrected by subtracting this non-specific disappearance of each BNP group. The GC-MS analysis was performed as previously described (15) on a GCMS-QP2010 system (Shimadzu, Kyoto, Japan) equipped with a fused silica capillary column (PetrocolTM DH, 100 m×0.25 mm×0.5 μm film thickness; Supelco/Sigma-Aldrich, Bellefonte, PA, USA). The oven temperature program was as follows: 3 min at 60°C, 15°C min−1 to 190°C, hold for 50 min, 15°C min−1 to 300°C, and hold for 10 min isothermally. A one-way analysis of variance (ANOVA) followed by Tukey’s honestly significant difference (HSD) test were used to test for significant differences in the degradation ratios of BNP isomer groups for each transformant (13).
The genome of the strain DSM 16289T consisted of one circular chromosome (SAMIE1; 4,221,764 bp, 64.8% G+C content, 3,614 coding sequences [CDSs]) with a coverage of 212-fold, and six circular plasmids, pSAMIE2 (sequence coverage, 195-fold; 212,396 bp, 63.3% G+C, 180 CDSs), pSAMIE3 (sequence coverage, 219-fold; 212,121 bp, 63.9% G+C, 178 CDSs), pSAMIE4 (sequence coverage, 261-fold; 55,670 bp, 61.4% G+C, 51 CDSs), pSAMIE5 (sequence coverage, 116-fold; 39,623 bp, 62.5% G+C, 35 CDSs), pSAMIE6 (sequence coverage, 551-fold; 22,371 bp, 62.9% G+C, 21 CDSs), and pSAMIE7 (sequence coverage, 1,016-fold; 9,977 bp, 60.9% G+C, 10 CDSs). The chromosome carried six rRNA operons and 54 tRNA genes. An opdA homolog (opdADSM16289) was present in pSAMIE2, while no nmoA homolog was found in the chromosome or any of the plasmids. This was in contrast to strain JCM 10874T, which possessed not only an opdA homolog, but also the nmoA gene, identical to that of strain NP5 (26). The predicted amino acid sequence (535 amino acids) from opdADSM16289 showed 99.4% amino acid sequence identity to that from the opdA of strain PWE1 (opdAPWE1) and 99.8, 99.4, and 99.1% identities to those from the opdA homologs of strain JCM 10874T (opdAJCM10874), strain Bayram (opdABayram), and strain TTNP 3 (opdATTNP3), respectively. As described by Takeo et al. (31), the opdADSM16289-coded protein also showed lower identity (83.6%) to the nmoA-coded protein (Supplementary Fig. S1). A phylogenetic tree constructed from the predicted protein sequences of the known opdAs and nmoAs is shown in Supplementary Fig. S2. A gene cluster (7,939 bp; hqdR, hqdA, hqdB, orf1, orf2, hqdC, hqdD, hqdE, and hqdF) encoding a hydroquinone catabolic pathway was found in pSAMIE4. The amino acid sequences predicted from the contiguous genes excluding orf1 and orf2 showed complete identity to those predicted from the gene cluster found in strain TTNP3 (17, 19).
To examine the effects of sequence differences among opdADSM16289, opdAJCM10874, and nmoAJCM10874 on substrate specificity, E. coli JM109 strains harboring pSAopdA, pSCopdA, and pSCnmoA were grown in the presence of 25 mg L−1 BNP, and the degradation profile was analyzed by GC-MS. All the transformed strains degraded BNP group 1 isomers more efficiently than with any other BNP group: the percent degradation rates of the BNP group 1 by the transformants harboring pSAopdA, pSCopdA, and pSCnmoA were 34, 62, and 48%, respectively. To compare the degradation degrees of BNP isomer groups, we calculated the normalized ratio (called the degradation ratio) of the degradation rate for each BNP isomer group to that for isomer group 1. E. coli JM109 harboring pSAopdA showed lower degradation ratios against BNP isomer groups 3, 4, and 5 (degradation ratios, 0.33, 0.24, and 0.20, respectively), which was consistent with our previous findings obtained using the original Sphingobium strains (15). In the case of E. coli JM109 harboring pSCopdA, the degradation ratios for BNP groups 3, 4, and 5 were also lower: 0.65, 0.44, and 0.46, respectively. In contrast, E. coli JM109 harboring pSCnmoA showed less preference to any BNP isomer group: degradation ratios, 1.0, 0.77, and 0.62 for BNP isomer groups 3, 4, and 5, respectively.
The predicted amino acid sequence of opdADSM16289 differed from that of opdAJCM10874 by only one amino acid (F241V), while it differed from those of opdAPWE1 and opdABayram by three amino acids (M119L, K205Q, and F241A) and that of opdATTNP3 by five amino acids (G58A, L87S, M119L, K205E, and F241C) (Table 2). Based on this result, the amino acid residue at position 241 appears to be important for substrate specificity. Porter et al. (28) proposed a homology model of the predicted structure of OpdA and suggested that the amino acid residues at positions 58 and 241 were located in the vicinity of the substrate-binding pocket, which was also suggested by our protein structure prediction using SWISS-MODEL with the OpdA-related flavoprotein p-hydroxybenzoate hydroxylase (EC 1.14.13.2) (12, 35). This was also the case for the nmoA-coded proteins (531 amino acids) of strains NP5 and JCM 10874T: G54 and A237, respectively. As summarized in Table 2, the change at residue 58 was only found in OpdATTNP3 and was a conservative change from glycine to alanine. Therefore, the change at residue 241 near the substrate-binding pocket may be assumed to partly influence substrate specificity. This appears to be supported by our results showing that strain DSM 16289T had the opdA homolog with narrow range substrate degradation activity (degradation ratio; 0.20 to 0.33 for BNP groups 3–5) and strain JCM 10874T had nmoA (degradation ratio; 0.62 to 1.02 for BNP groups 3–5) and the opdA homolog (degradation ratio; 0.44 to 0.65 for BNP groups 3–5) with more versatile substrate specificity.
Table 2
Differences in amino acid residues predicted from
opdA and
nmoA, encoding enzymes for branched nonylphenol degradation.
| Protein/Strain (Accession number) |
Amino acid residue at the position of OpdA (NmoA) |
|
| 58 (54) |
87 (83) |
119 (115) |
205 (201) |
241 (237) |
| OpdA |
| Sphingobium amiense DSM 16289T (BBE00139) |
G |
L |
M |
K |
F |
| Sphingobium cloacae JCM 10874T (BAV66737) |
G |
L |
M |
K |
V |
| Sphingomonas sp. PWE1 (ABU98341) |
G |
L |
L |
Q |
A |
| Sphingobium xenophagum Bayram (ACI12953) |
G |
L |
L |
Q |
A |
| Sphingomonas sp. TTNP3 (ACI12952) |
A |
S |
L |
E |
C |
| NmoA |
| Sphingobium cloacae JCM 10874T (BAV66828) |
G |
L |
L |
E |
A |
| Sphingomonas sp. NP5 (BAI43691/BAI43694) |
G |
L |
L |
E |
A |
In conclusion, the present results suggest that strains JCM 10874T (26) and DSM 16289T degrade BNP by the concerted effects of different plasmids separately harboring the initial oxygenation gene (opdA) and hydroquinone catabolic pathway genes. The different preference of the two strains for BNP isomers may partly result from nmoA in strain DSM 10874T and the different structure of the substrate-binding pockets in their OpdAs. To further confirm this, structure-activity analyses of OpdAs from the other BNP-degrading Sphingomonas/Sphingobium strains are needed.
Accession number
The complete genome sequences of S. amiense DSM 16289T were deposited into the DDBJ/EMBL/GenBank databases under the accession numbers AP018664 to AP018670 and were linked to the BioProject with the accession number PRJDB6948.
Acknowledgements
The authors thank Drs. Yong Guo and Akito Nishizawa for their help with the genetic analysis. This work was supported partially by a KAKENHI grant from JSPS to HO (13480151).
References
- 1. Ahel, M., J. McEvoy, and W. Giger. 1993. Bioaccumulation of the lipophilic metabolites of nonionic surfactants in freshwater organisms. Environ Pollut. 79:243-248.
- 2. Ahel, M., D. Hršak, and W. Giger. 1994. Aerobic transformation of short-chain alkylphenol polyethoxylates by mixed bacterial cultures. Arch Environ Contam Toxicol. 26:540-548.10.1007/BF00214159
- 3. Altschul, S.F., T.L. Madden, A.A. Schäffer, J. Zhang, Z. Zhang, W. Miller, and D.J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389-3402.10.1093/nar/25.17.3389
- 4. Bankevich, A., S. Nurk, D. Antipov, et al. 2012. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 19:455-477.10.1089/cmb.2012.0021
- 5. Corvini, P.F., R.J. Meesters, A. Schaffer, H.F. Schroder, R. Vinken, and J. Hollender. 2004. Degradation of a nonylphenol single isomer by Sphingomonas sp. strain TTNP3 leads to a hydroxylation-induced migration product. Appl Environ Microbiol. 70:6897-6900.10.1128/AEM.70.11.6897-6900.2004
- 6. de Vries, Y.P., Y. Takahara, Y. Ikunaga, Y. Ushiba, M. Hasegawa, Y. Kasahara, H. Shimomura, S. Hayashi, Y. Hirai, and H. Ohta. 2001. Organic nutrient-dependent degradation of branched nonylphenol by Sphingomonas sp. YT isolated from a river sediment sample. Microbes Environ. 16:240-249.10.1264/jsme2.2001.240
- 7. Fujii, K., N. Urano, H. Ushio, M. Satomi, and S. Kimura. 2001. Sphingomonas cloacae sp. nov., a nonylphenol-degrading bacterium isolated from wastewater of a sewage-treatment plant in Tokyo. Int J Syst Evol Microbiol. 51:603-610.10.1099/00207713-51-2-603
- 8. Gabriel, F.L., W. Giger, K. Guenther, and H.P. Kohler. 2005. Differential degradation of nonylphenol isomers by Sphingomonas xenophaga Bayram. Appl Environ Microbiol. 71:1123-1129.10.1128/AEM.71.3.1123-1129.2005
- 9. Gabriel, F.L., A. Heidlberger, D. Rentsch, W. Giger, K. Guenther, and H.P. Kohler. 2005. A novel metabolic pathway for degradation of 4-nonylphenol environmental contaminants by Sphingomonas xenophaga Bayram: ipso-hydroxylation and intramolecular rearrangement. J Biol Chem. 280:15526-15533.10.1074/jbc.M413446200
- 10. Gabriel, F.L.P., M. Cyris, N. Jonkers, W. Giger, K. Guenther, and H.-P.E. Kohler. 2007. Elucidation of the ipso-substitution mechanism for side-chain cleavage of α-quaternary 4-nonylphenols and 4-t-butoxyphenol in Sphingobium xenophagum Bayram. Appl Environ Microbiol. 73:3320-3326.10.1128/aem.02994-06
- 11. Gao, F., and C.-T. Zhang. 2006. GC-Profile: a web-based tool for visualizing and analyzing the variation of GC content in genomic sequences. Nucleic Acids Res. 34:W686-W691.10.1093/nar/gkl040
- 12. Gatti, D., B. Palfey, M. Lah, B. Entsch, V. Massey, D. Ballou, and M. Ludwig. 1994. The mobile flavin of 4-OH benzoate hydroxylase. Science. 266:110-114.10.1126/science.7939628
- 13. Guo, Y., R. Fujimura, Y. Sato, W. Suda, S.W. Kim, K. Oshima, M. Hattori, T. Kamijo, K. Narisawa, and H. Ohta. 2014. Characterization of early microbial communities on volcanic deposits along a vegetation gradient on the Island of Miyake, Japan. Microbes Environ. 29:38-49.10.1264/jsme2.ME13142
- 14. Huijbers, M.M.E., S. Montersino, A.H. Westphal, D. Tischler, and W.J.H. van Berkel. 2014. Flavin dependent monooxygenases. Arch Biochem Biophys. 544:2-17.10.1016/j.abb.2013.12.005
- 15. Ikunaga, Y., S. Miyakwa, M. Hasegawa, Y. Kasahara, O. Kodama, and H. Ohta. 2004. Degradation profiles of branched nonylphenol isomers by Sphingobium amiense and Sphingomonas cloacae. Soil Sci Plant Nutr. 50:871-875.10.1080/00380768.2004.10408548
- 16. John, D.M., and G.F. White. 1998. Mechanism for biotransformation of nonylphenol polyethoxylates to xenoestrogens in Pseudomonas putida. J Bacteriol. 180:4332-4338.
- 17. Kolvenbach, B.A., M. Lenz, D. Benndorf, E. Rapp, J. Fousek, C. Vlcek, A. Schäffer, F.L.P. Gabriel, H.-P.E. Kohler, and P.F.X. Corvini. 2011. Purification and characterization of hydroquinone dioxygenase from Sphingomonas sp. strain TTNP3. AMB Express. 1:810.1186/2191-0855-1-8
- 18. Kolvenbach, B.A., and P.F.X. Corvini. 2012. The degradation of alkylphenols by Sphingomonas sp. strain TTNP3—a review on seven years of research. New Biotechnol. 30:88-95.10.1016/j.nbt.2012.07.008
- 19. Kolvenbach, B.A., H. Dobrowinski, J. Fousek, C. Vlcek, A. Schäffer, F.L.P. Gabriel, H.-P.E. Kohler, and P.F.X. Corvini. 2012. An unexpected gene cluster for downstream degradation of alkylphenols in Sphingomonas sp. strain TTNP3. Appl Microbiol Biotechnol. 93:1315-1324.10.1007/s00253-011-3451-8
- 20. Koren, S., B.P. Walenz, K. Berlin, J.R. Miller, N.H. Bergman, and A.M. Phillippy. 2017. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. doi:10.1101/gr.215087.116
- 21. Kumar, S., G. Stecher, and K. Tamura. 2016. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 33:1870-1874.10.1093/molbev/msw054
- 22. Lowe, T.M., and S.R. Eddy. 1997. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 25:955-964.10.1093/nar/25.5.0955
- 23. Lu, Z., and J. Gan. 2014. Analysis, toxicity, occurrence and biodegradation of nonylphenol isomers: A review. Environ Int. 73:334-345.
- 24. Maki, H., N. Masuda, Y. Fujiwara, M. Ike, and M. Fujita. 1994. Degradation of alkylphenol ethoxylates by Pseudomonas sp. strain TR01. Appl Environ Microbiol. 60:2265-2271.
- 25. Nishizawa, T., A. Ueda, T. Nakano, A. Nishizawa, T. Miura, M. Asayama, K. Fujii, K. Harada, and M. Shirai. 2011. Characterization of the locus of genes encoding enzymes producing heptadepsipeptide micropeptin in the unicellular cyanobacterium Microcystis. J Biochem. 149:475-485.10.1093/jb/mvq150
- 26. Ootsuka, M., T. Nishizawa, and H. Ohta. 2016. Complete genome sequence of the nonylphenol-degrading bacterium Sphingobium cloacae JCM 10874T. Genome Announc. 4:e01358-1610.1128/genomeA.01358-16
- 27. Porter, A.W., and A.G. Hay. 2007. Identification of opdA, a gene involved in biodegradation of the endocrine disrupter octylphenol. Appl Environ Microbiol. 73:7373-7379.10.1128/AEM.01478-07
- 28. Porter, A.W., B.R. Campbell, B.A. Kolvenbach, P.F.-X. Corvini, D. Benndorf, G. Rivera-Cancel, and A.G. Hay. 2012. Identification of the flavin monooxygenase responsible for ipso substitution of alkyl and alkoxyphenols in Sphingomonas sp. TTNP3 and Sphingobium xenophagum Bayram. Appl Microbiol Biotechnol. 94:261-272.10.1007/s00253-011-3621-8
- 29. Sambrook, J., E.F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
- 30. Soto, A.M., H. Justicia, J.W. Wray, and C. Sonnenschein. 1991. p-Nonyl-phenol: an estrogenic xenobiotic released from “modified” polystyrene. Environ Health Perspect. 92:167-173.10.2307/3431154
- 31. Takeo, M., Y. Maeda, J. Maeda, N. Nishiyama, C. Kitamura, D. Kato, and S. Negoro. 2012. Two identical nonylphenol monooxygenase genes linked to IS6100 and some putative insertion sequence elements in Sphingomonas sp. NP5. Microbiology. 158:1796-1807.10.1099/mic.0.055335-0
- 32. Tanghe, T., W. Dhooge, and W. Verstraete. 1999. Isolation of a bacterial strain able to degrade branched nonylphenol. Appl Environ Microbiol. 65:746-751.
- 33. Thompson, J.D., D.G. Higgins, and T.J. Gibson. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22:4673-4680.10.1093/nar/22.22.4673
- 34. Ushiba, Y., Y. Takahara, and H. Ohta. 2003. Sphingobium amiense sp. nov., a novel nonylphenol-degrading bacterium isolated from a river sediment. Int J Syst Evol Microbiol. 53:2045-2048.10.1099/ijs.0.02581-0
- 35. Wang, J., M. Ortiz-Maldonado, B. Entsch, V. Massey, D. Ballou, and D.L. Gatti. 2002. Protein and ligand dynamics in 4-hydroxybenzoate hydroxylase. Proc Natl Acad Sci USA. 99:608-613.10.1073/pnas.022640199
- 36. Wheeler, T.J.R., J.R. Heim, M.R. LaTorre, and A.B. Janes. 1997. Mass spectral characterization of p-nonylphenol isomers using high-resolution capillary GC-MS. J Chromatogr Sci. 35:19-30.
- 37. White, R., S. Jobling, S.A. Hoare, J.P. Sumpter, and M.G. Parker. 1994. Environmentally persistent alkylphenolic compounds are estrogenic. Endocrinology. 135:175-182.10.1210/endo.135.1.8013351
- 38. Wick, R.R., L.M. Judd, C.L. Gorrie, and K.E. Holt. 2017. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. 13:e100559510.1371/journal.pcbi.1005595

;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/33_450_1.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)
;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/33_450_2.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)
;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/33_450_3.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)
;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/33_450_4.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)
;%0A%09%09%09newWindow.document.open();%0A%09%09%09newWindow.document.write('<img src=%22./Graphics/33_450_5.jpg%22>');%0A%09%09%09newWindow.document.close();%0A%09%09)