Abstract
The responses of bacterial communities to halocarbon were examined using a 28-d incubation of bromoform- and methanol-enriched subarctic surface seawater. Significant increases were observed in dibromomethane concentrations and bacterial 16S rRNA gene copy numbers in the treated substrates incubated for 13 d. The accumulated bacterial community was investigated by denaturing gradient gel electrophoresis and amplicon analyses. The dominant genotypes corresponded to the genera Roseobacter, Lentibacter, and Amylibacter; the family Flavobacteriaceae; and the phylum Planctomycetes, including methylotrophs of the genus Methylophaga and the family Methylophilaceae. Therefore, various phylotypes responded along with the dehalogenation processes in subarctic seawater.
Brominated, very short-lived halocarbons, such as bromoform (CHBr3) and dibromomethane (CH2Br2), with atmospheric lifetimes of 24 and 123 d, respectively (25), are potentially significant contributors to catalytic ozone loss in the troposphere and lower stratosphere (12). Although these two halocarbons are heterogeneously distributed along the depth of oceanic environments (26), their production, degradation, and transformation processes remain unknown. Abiotically, anoxic hydrolysis may result in the transformation of CHBr3 to CH2Br2 under limited conditions, such as with the activity of reduction catalysts or strong sunlight (1). The sources of these two halocarbons are considered to be associated with phytoplankton and macroalgae present in the subsurface of the ocean (20). Bacterial production and the loss of halocarbons have been proposed, with the dehalogenation of CHBr3 and subsequent production of CH2Br2 through microbial activity representing a potential mechanism (18). CH2Br2 concentrations correlate with the abundance of Synechococcus, picoeukaryotes, and heterotrophic bacteria as well as with the total chlorophyll a concentration in the Atlantic Ocean (11). These findings suggest that CH2Br2 production is attributable to biogeochemical processes related to the phytoplankton biomass, such as degradation by heterotrophic bacteria and grazing by animals. To date, only three strains of marine class Alphaproteobacteria (Phaeobacter gallaeciensis, Roseobacter denitrificans, and Rhodobacter vinaykumarii) are known to decompose CHBr3 to CH2Br2 (6). The bacterial production of CH2Br2 occurs from a relatively early exponential phase of bacterial growth. In contrast to CHBr3, CH2Br2 has not been shown to be decomposed by these three strains. Thus, aquatic bacteria may act as new sinks for CHBr3 and new sources for CH2Br2 in marine environments. The purposes of the present study were to evaluate the responses of bacterial communities to the addition of CHBr3 to subarctic surface seawater and to identify the bacteria phylotypes involved in CH2Br2 production as a response to this addition using a 28-d incubation.
Seawater samples were collected from the eastern coast of Hokkaido, the northern area of Japan, located in the subarctic gyre of the western North Pacific (42.3°N, 140.6°E) by T/V Ushio-Maru on December 12th, 2016. Seawater was taken from a depth of approximately 5 m, at which temperature and salinity were +8.4°C and 33.9, respectively, and then filtered through a Nuclepore filter with a pore size of 10 μm (47 mm in diameter; Whatman) to remove large-sized zooplankton and phytoplankton. Four 1.2-L amber glass bottles were filled with filtered water and closed using a Teflon-coated inter-lid (GL45 and A17200-001, respectively; Shibata Scientific Technology) without head space to avoid gas exchange between the inside and outside of the bottle. Glass bottles were stored in a refrigerator (+4°C) onboard until further processing. To imitate the natural environment in which the CHBr3 concentration, which reached 1.2 nmol L−1, matched the phytoplankton biomass distribution (17), 8 nmol of CHBr3 (027-10142; Wako Pure Chemical Industries) was diluted in 99 μmol of methanol (CH3OH) (139-08821; Wako Pure Chemical Industries) and added to the glass bottle (labeled as “treatment”) in the laboratory on land within 24 h of sampling. Since it was not possible to predict the production rate of CH2Br2 from CHBr3, a sufficient amount of CHBr3 (8 nmol) was added to the treatment samples (a final concentration of 6.67 nmol L−1). Methanol was used as a solvent because CHBr3 is hardly soluble in water and is produced by phytoplankton in natural seawater (5). A bottle sample without the added reagents was also prepared (labeled as the “control”). No abiotic and only CH3OH controls were also prepared to save water and incubation space. Duplicate measurements for organic gas concentrations and bacterial parameters were conducted for each sample.
In the seawater incubation, glass bottles were stored at 4°C for 28 d in the dark to avoid photodegradation of the substrates (CHBr3 and CH2Br2) (1) and the growth of phytoplankton. In the analysis of organic gas concentrations, 30 mL of a subsample was carefully transferred into a 30-mL amber glass vial (Nichiden-Rika Glass) on days 0, 13, and 28. A mercuric chloride solution (saturated HgCl2; 200 μL) was added to the glass vials to stop biological activity. Vials were sealed with a Teflon-lined septum and aluminum cap (Nichiden-Rika Glass) and stored in the refrigerator (+4°C) until further analyses. In analyses of bacterial abundance and composition, a 200-mL subsample was filtered (pore size of 0.2 μm and diameter of 47 mm) and rapidly frozen at −30°C. CH2Br2 and CHBr3 concentrations were assessed using purge-and-trap and gas chromatography mass spectrometry (GC-MS) methods (16) (details in supporting information).
In molecular analyses, genomic DNA was extracted from the whole filter using a FastDNA Spin kit (Nippon gene) and quantified using a Qubit 3.0 fluorometer (Thermo Fisher Scientific). Real-time quantitative PCR (qPCR) was performed to quantify bacterial 16S rRNA gene copy numbers using the primer set of 1369F and 1492R (Table S1) as previously described (9) with some slight modifications (supporting information). Bacterial communities were investigated by denaturing gradient gel electrophoresis (DGGE) using the primer set of 907R and 341F attaching a GC-clamp to the 5′-end of the forward primer under conditions (shown in Table S1) described previously (7). To semi-quantitatively compare the genotype composition of the 16S rRNA gene, amplicon sequencing of the primer set of 341F and 805R was conducted (supporting information). The sequence obtained from the DGGE band and representative sequences of operational taxonomic units (OTU) from the amplicon sequencing analysis were compared with the GenBank database using BLASTn, and aligned against the SILVA alignment (SSU Ref NR 128) to obtain a NJ tree.
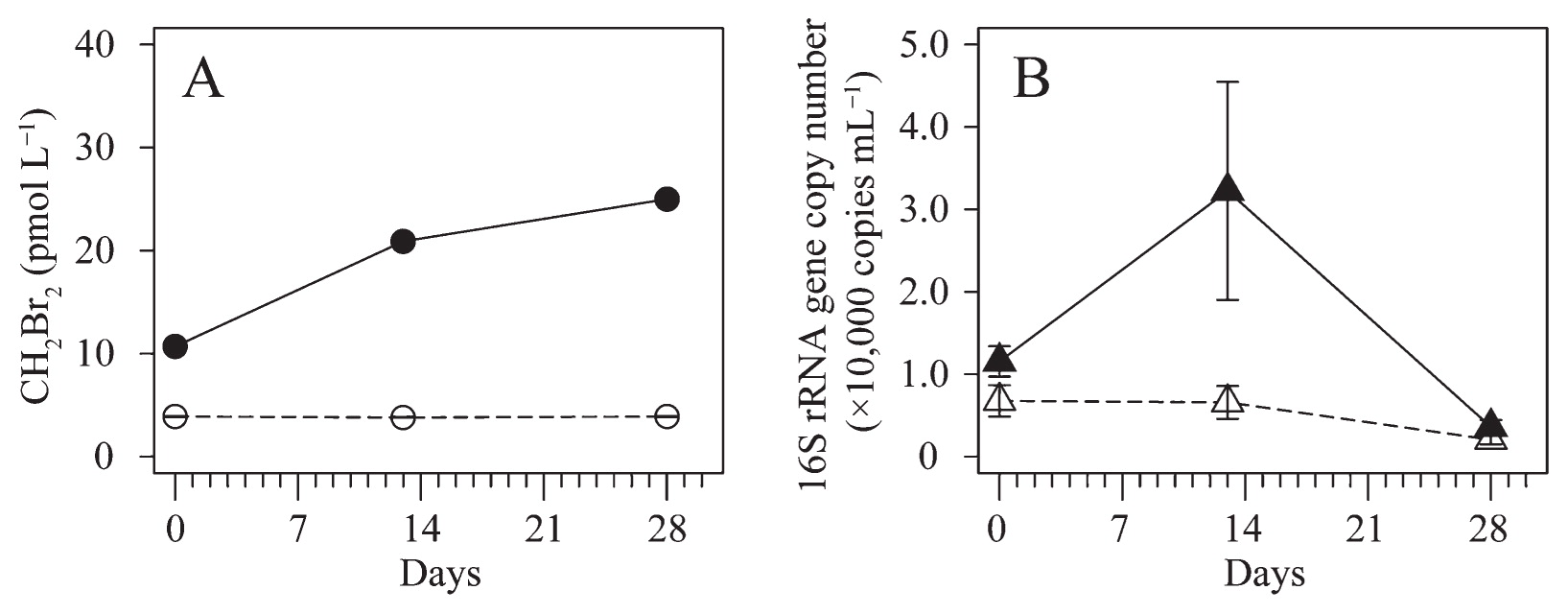
The time evolution of CH2Br2 concentrations and bacterial abundance, assessed by 16S rRNA gene copy numbers during the incubation, is shown in Fig. 1. CH2Br2 concentrations increased with time from 10.8 to 25.1 pmol L−1 in the treatment samples, but remained constant in the control samples (4.0±0.1 pmol L−1) (mean±standard deviation) (Fig. 1A). The difference observed in CH2Br2 concentrations between the control and treatment bottles on day 0 was presumably due to contamination of the CHBr3 solution (0.1% of CHBr3 solution). Bacterial abundance was greater in the treatment bottles than in the control bottles and showed a marked increase from 9.7×103 and 1.34×104 copies mL−1 on day 0 to 4.6×104 and 1.9×104 copies mL−1, respectively, on day 13 (Fig. 1B). The low copy number on day 28 indicates the death phase of the batch culture caused by a nutrient shortage after increases in some genotypes. CHBr3 consumption was not clearly detected (Fig. S1) because the initial concentration of this halocarbon was so high that its degradation into CH2Br2 was smaller than the variance due to handling errors. Although the utilization of CHBr3 and/or CH3OH, which was added as a solvent, was not measurable, the present results suggest that the sea surface bacterial community in the eastern coast of Hokkaido is involved in the production of CH2Br2.
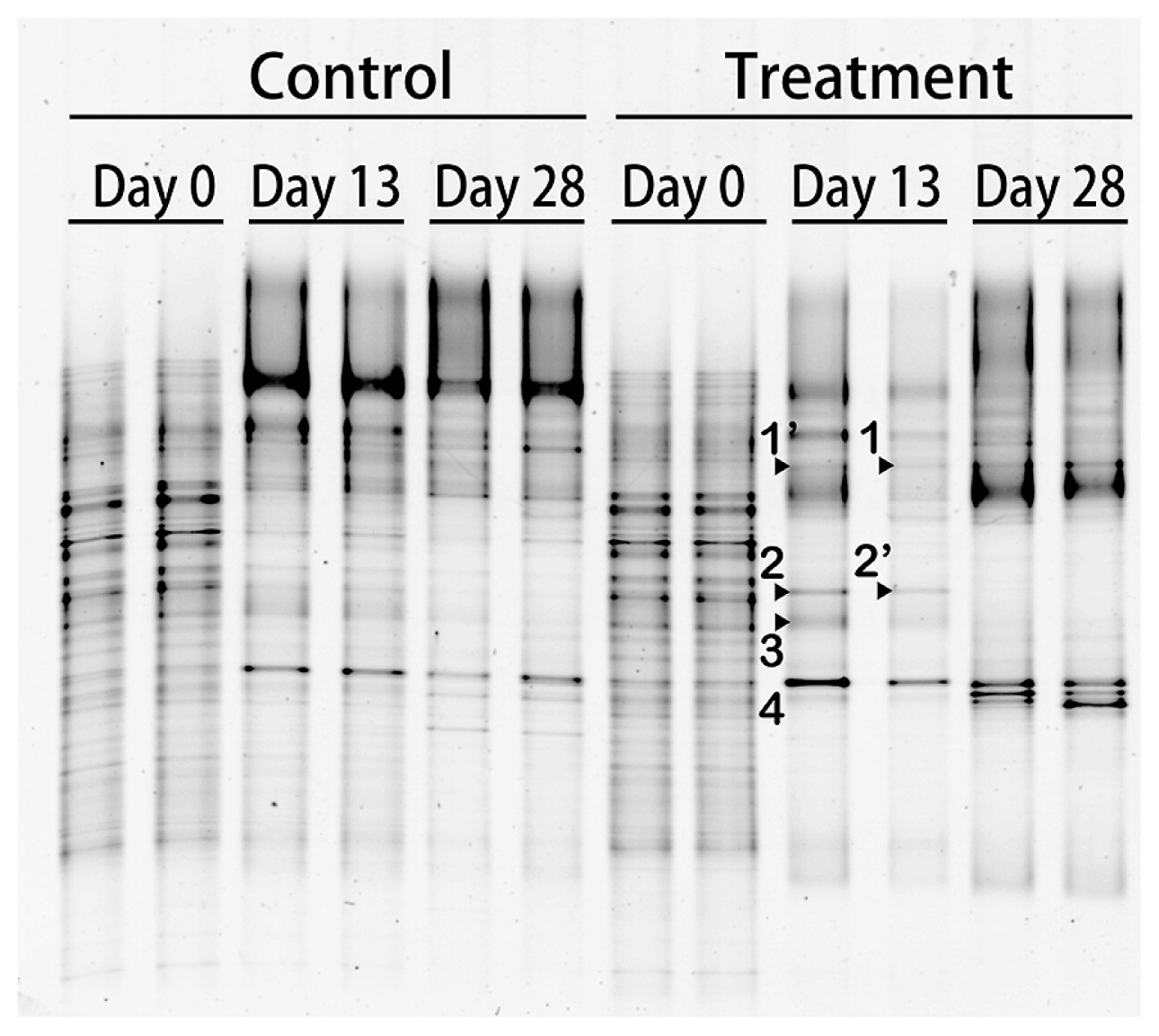
A DGGE analysis was conducted to monitor the bacterial community during the experiment (Fig. 2). The DGGE band pattern in the control bottles showed a small change in the band position during the incubation period, with some bands showing marked changes in their intensities. Conversely, some intense DGGE bands in the treatment bottles specifically appeared on day 13 (bands of vg-1, -2, and -3 in Fig. 2) along with the disappearance of most of the DGGE bands that were present on day 0. A 16S rRNA gene amplicon sequencing analysis was conducted on day 13 to compare genotype richness between the control and treatment samples. The number of OTUs, which were grouped based on a similarity level of 97%, was lower in the treatment bottles than in the control bottles (Fig. S2), indicating that the bacteria community was enriched with particular genotypes that adapted to amended CHBr3 and/or CH3OH. To examine the accumulated OTUs, the ratio of the relative abundance of each OTU in the treatment bottles to that in the control bottles was calculated by the average of the replicates on day 13 (Table S2). Among the 960–961 OTUs in the treatment bottles, 23 showed >10-fold higher values for this ratio than OTUs in the control bottles or were exclusively detected in the treatment bottles with a relative abundance >0.01%. One OTU of VSN07, which was close to the genus Methylophaga of class Gammaproteobacteria (Fig. 3), was significantly dominant in the treatment bottles (65.9 and 73.5%). Methylophaga-related genotypes were also previously shown to increase their abundance in methyl bromide-amended seawater during phytoplankton blooms in the English Channel (15). Thus, the present results suggest that Methylophaga-related genotypes respond to CHBr3 and/or CH3OH in the eastern coast of Hokkaido.
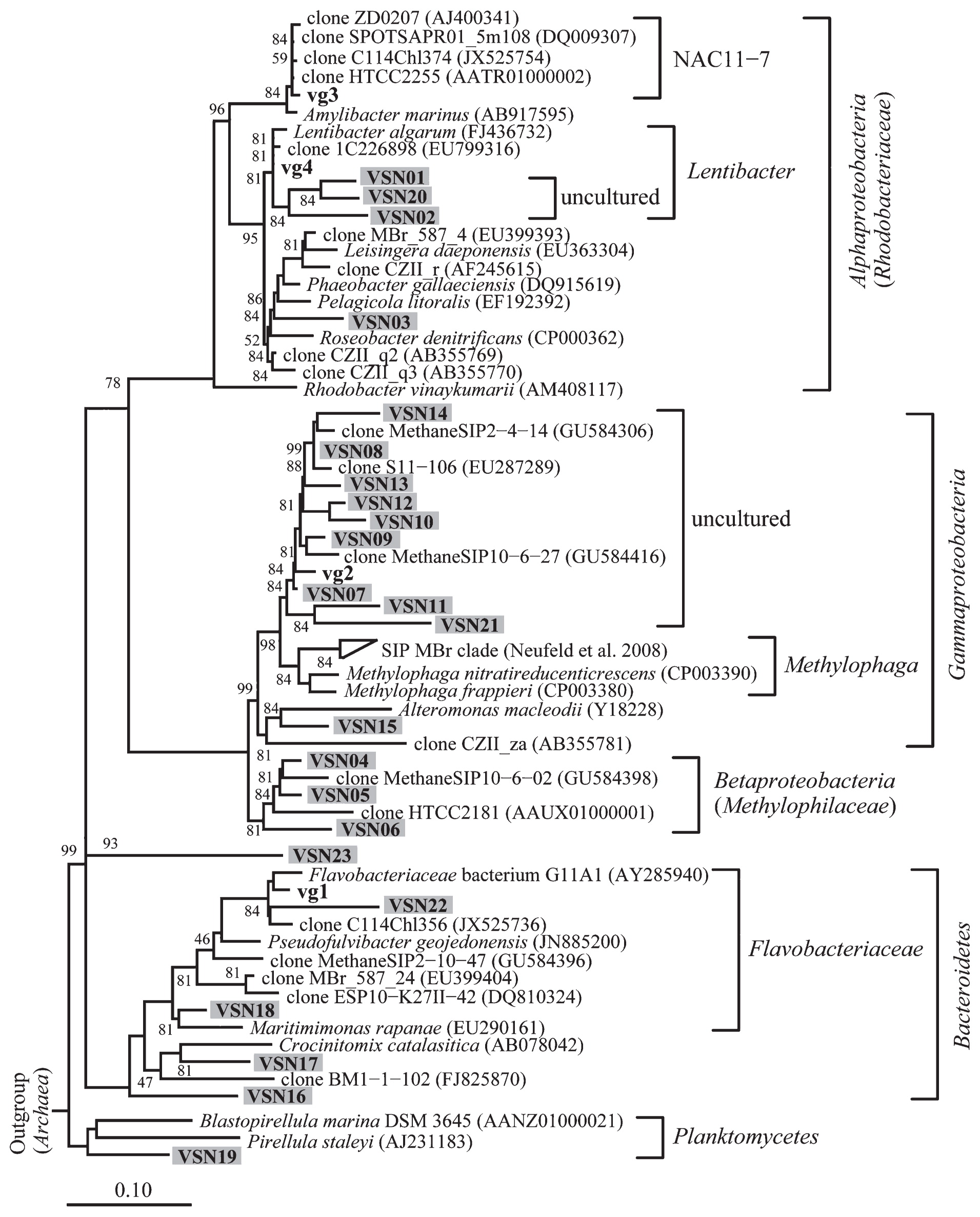
Sequence analyses revealed the phylogenetic positions of the genotypes that responded to the addition of CHBr3 and CH3OH after an incubation for 13 d (Fig. 3). Four sequences from the DGGE analysis were close to those obtained from phytoplankton blooms or were related to methanotrophic strains (8). Two sequences of vg-3 and vg-4 belonged to the family Rhodobacteraceae in the class Alphaproteobacteria. The vg-3 genotype, which was close to the recently defined taxa of Amylibacter manrinus (22), belongs to the NAC11-7 clade comprising various genotypes detected after phytoplankton blooms (2, 21, 24). The vg-4 genotype was close to Lentibacter algarum isolates obtained from massive algal blooms in China (10). The sequence of vg-1, which belongs to the family Flavobacteriaceae, is close to that of an abundant genotype in phytoplankton blooms after iron fertilization (clone C114Chl356 [19]). Thus, the major genotypes responding to CHBr3 and/or CH3OH were related to natural phytoplankton blooms. The results of the amplicon sequencing analysis corresponded to those of the DGGE analysis, except for the vg-3 genotype to which no close sequences were detected (Fig. 3 and S3). Most of the responding genotypes in the treatment bottles (Table S2) were similar to those identified from the DGGE sequences (i.e., vg-1, -2, and -4). The family Methylophilaceae in the class Betaproteobacteria is a major phylogenetic group of the OM43 clade (4, 13), which possesses the methanol dehydrogenase enzyme (XoxF) (3). Thus, the genotypes of VSN04–06 may have been involved in methanol (CH3OH) degradation even if their relative abundance was low (Table S2). The other methylotroph-related genotypes (VSN07–14 and 21) were close to the genus Methylophaga in the class Gammaproteobacteria, which uses CH3OH as a source of carbon (23). The diverse and abundant genotypes were only close to uncultured bacteria-derived sequences. Neufeld et al. (14, 15) also found diverse uncultured CH3Br-assimilating genotypes of the genus Methylophaga (SIP MBr clade); however, their sequences were different from those obtained in the present study. Although it currently remains unclear whether the methylotrophs detected in the present study contribute to the production of CH2Br2, the diverse but local genotypes that formed a clade in the phylogenetic tree (Fig. 3) increased their abundance in response to the addition of CHBr3 and/or CH3OH. Conversely, one genotype of VSN03, belonging to the class Alphaproteobacteria, was close to P. gallaeciensis, which produces CH2Br2 in CHBr3-amended media during the early growth phase (6), suggesting that VSN03 was a source of CH2Br2 in our experiment. The other abundant genotypes in the treatment bottles belonged to the genus Lentibacter in the class Alphaproteobacteria and the phyla Bacteroidetes and Planctomycetes. Although their capacity to metabolize halogenated alkanes remains unknown, there is scope for assessing their metabolism because CFB and some unknown phylotypes have been shown to incorporate 13CHBr3 (15). A second diverse clade formed by VSN01, 02, and 20 within the genus Lentibacter may also metabolize C1 methyl halides.
In conclusion, the present results suggest that the eastern coast of Hokkaido is inhabited by a bacterial community that responds to CHBr3 and/or CH3OH and plays a major role in dehalogenation. Pure cultures of these genotypes will provide insights into the role of halocarbon metabolism in carbon and halocarbon cycles in the ocean.
Acknowledgements
We wish to thank the captain and crew of the T/V Ushio-Maru for their helpful support. This study was performed under the Joint Research Program of the Japan Arctic Research Network Center. We also thank A. Kataoka for support.
References
- 1. Betterton, E.A., R.G. Arnold, R.J. Kuhler, and G.A. Santo. 1995. Reductive dehalogenation of bromoform in aqueous solution. Environ Health Perspect. 103:89-91.
- 2. Buchan, A., G.R. LeCleir, C.A. Gulvik, and J.M. González. 2014. Master recyclers: features and functions of bacteria associated with phytoplankton blooms. Nat Rev Microbiol. 12:686.
- 3. Giovannoni, S.J., D.H. Hayakawa, H.J. Tripp, et al. 2008. The small genome of an abundant coastal ocean methylotroph. Environ Microbiol. 10:1771-1782.
- 4. Hanson, B.T., I. Hewson, and E.L. Madsen. 2014. Metaproteomic survey of six aquatic habitats: Discovering the identities of microbial populations active in biogeochemical cycling. Microb Ecol. 67:520-539.
- 5. Heikes, B.G., W.N. Chang, M.E.Q. Pilson, et al. 2002. Atmospheric methanol budget and ocean implication. Global Biogeochem Cycles. 16.
- 6. Ichikawa, K., M. Kurihara, H. Tamegai, and S. Hashimoto. 2015. Decomposition of brominated organic halogens by cultures of marine proteobacteria: Phaeobacter, Roseobacter, and Rhodobacter. Mar Chem. 176:133-141.
- 7. Kataoka, T., Y. Hodoki, K. Suzuki, H. Saito, and S. Higashi. 2009. Tempo-spatial patterns of bacterial community composition in the western North Pacific Ocean. J Mar Syst. 77:197-207.
- 8. Kataoka, T., K. Suzuki, M. Hayakawa, I. Kudo, S. Higashi, and A. Tsuda. 2009. Temporal changes in community composition of heterotrophic bacteria during in situ iron enrichment in the western subarctic Pacific (SEEDS-II). Deep Sea Res, Part II. 56:2779-2787.
- 9. Kataoka, T., S. Mitsunobu, and N. Hamamura. 2018. Influence of the chemical form of antimony on soil microbial community structure and arsenite oxidation activity. Microbes Environ. 33:214-221.
- 10. Li, Z., Z. Qu, X.M. Zhang, and X.H. Zhang. 2012. Lentibacter algarum gen. nov., sp nov., isolated from coastal water during a massive green algae bloom. Int J Syst Evol Microbiol. 62:1042-1047.
- 11. Liu, Y., S.A. Yvon-Lewis, D.C.O. Thornton, J.H. Butler, T.S. Bianchi, L. Campbell, L. Hu, and R.W. Smith. 2013. Spatial and temporal distributions of bromoform and dibromomethane in the Atlantic Ocean and their relationship with photosynthetic biomass. J Geophys Res : Oceans. 118:3950-3965.
- 12. Montzka, S.A., and S. Reimann. 2011. Ozone-depleting substances (ODSs) and related chemicals, p.1-108. In C.A. Ennis (ed.), Global Ozone Research and Monitoring Project-Report. 52. World Meteorological Organization, Geneva.
- 13. Morris, R.M., K. Longnecker, and S.J. Giovannoni. 2006. Pirellula and OM43 are among the dominant lineages identified in an Oregon coast diatom bloom. Environ Microbiol. 8:1361-1370.
- 14. Neufeld, J.D., H. Schafer, M.J. Cox, R. Boden, I.R. McDonald, and J.C. Murrell. 2007. Stable-isotope probing implicates Methylophaga spp and novel Gammaproteobacteria in marine methanol and methylamine metabolism. ISME J. 1:480-491.
- 15. Neufeld, J.D., R. Boden, H. Moussard, H. Schäfer, and J.C. Murrell. 2008. Substrate-specific clades of active marine methylotrophs associated with a phytoplankton bloom in a temperate coastal environment. Appl Environ Microbiol. 74:7321-7328.
- 16. Ooki, A., and Y. Yokouchi. 2011. Dichloromethane in the Indian Ocean: Evidence for in-situ production in seawater. Mar Chem. 124:119-124.
- 17. Ooki, A., K. Miwa, H. Onishi, Y. Kamei, N. Kobayashi, and K. Kuma. 2013. Seasonal variations of bromoform distribution in the Funka Bay. Bunseki Kagaku. 62:1071-1078.
- 18. Quack, B., and D.W.R. Wallace. 2003. Air-sea flux of bromoform: Controls, rates, and implications. Global Biogeochem Cycles. 17:1023.
- 19. Singh, S.K., A. Kotakonda, R.K. Kapardar, et al. 2015. Response of bacterioplankton to iron fertilization of the Southern Ocean, Antarctica. Front Microbiol. 6:863.
- 20. Sturges, W.T., C.W. Sullivan, R.C. Schnell, L.E. Heidt, and W.H. Pollock. 1993. Bromoalkane production by Antarctic ice algae. Tellus B. 45:120-126.
- 21. Teeling, H., B.M. Fuchs, D. Becher, et al. 2012. Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom. Science. 336:608-611.
- 22. Teramoto, M., and M. Nishijima. 2014. Amylibacter marinus gen. nov., sp nov., isolated from surface seawater. Int J Syst Evol Microbiol. 64:4016-4020.
- 23. Villeneuve, C., C. Martineau, F. Mauffrey, and R. Villemur. 2012. Complete genome sequences of Methylophaga sp. strain JAM1 and Methylophaga sp. strain JAM7. J Bacteriol. 194:4126-4127.
- 24. Wemheuer, B., F. Wemheuer, J. Hollensteiner, F.-D. Meyer, S. Voget, and R. Daniel. 2015. The green impact: bacterioplankton response toward a phytoplankton spring bloom in the southern North Sea assessed by comparative metagenomic and metatranscriptomic approaches. Front Microbiol. 6:805.
- 25. WMO (World Meteorological Organization). 2011. Scientific Assessment of Ozone Depletion: 2010, Geneva.
- 26. Yamamoto, H., Y. Yokouchi, A. Otsuki, and H. Itoh. 2001. Depth profiles of volatile halogenated hydrocarbons in seawater in the Bay of Bengal. Chemosphere. 45:371-377.
