Abstract
Fluorescence in situ hybridization (FISH) is a widely used molecular technique in microbial ecology. However, the non-specific adsorption of fluorescent probes and resulting high intensity of background signals from mineral particles hampers the specific detection of microbial cells in grain-rich environmental samples, such as subseafloor sediments. We herein demonstrated that a new buffer composition containing EDTA efficiently reduced the adsorption of probes without compromising the properties of the FISH-based probing of microbes. The inclusion of a high concentration of EDTA in the buffer in our protocol provides a simple and effective approach for reducing the background in FISH for environmental samples.
The direct visualization of microbes by fluorescence in situ hybridization (FISH) allows for the spatial and temporal identification of the dynamics of individual active microbial populations and is widely used in microbial ecology (Amann and Fuchs, 2008; Dianou et al., 2012; Hirakata et al., 2015). Recent developments that have increased the sensitivity of FISH (Furukawa et al., 2006; Kawakami et al., 2010; Kubota, 2013) as well as its combination with techniques such as transmission electron microscopy (Halary et al., 2011) and nano-scale secondary ion mass spectrometry (Dekas and Orphan, 2011; Morono et al., 2011) have further enhanced its utility in identifying specific microbes with respective functions in an environment.
However, the application of FISH-based probing is hampered by the presence of inorganic mineral grains that adsorb DNA (Ogram et al., 1988; Bezanilla et al., 1995; Saeki et al., 2010). When a standard FISH protocol is applied to subseafloor sediment samples, bright fluorescence is frequently observed on mineral particles, speculatively due to the adsorption of fluorescent oligonucleotide probes onto mineral surfaces. Furthermore, the DNA-binding dyes used to stain all microbes in samples are adsorbed onto mineral surfaces. This non-specific adsorption negatively affects the identification of active microbial populations.
To prevent the non-specific adsorption of fluorescent probes and, thus, reduce background signals, we used a high concentration of EDTA in the hybridization buffer for FISH. EDTA is a chelating agent that is widely used to chelate divalent cations, thereby preventing the degradation of DNA by deoxyribonucleases. A previous study demonstrated that the presence of a high concentration of EDTA increased the efficiency of DNA recovery in extractions from environmental samples by preventing DNA adsorption by mineral particles in soil (Krsek and Wellington, 1999; Rai et al., 2010). If DNA adsorption is prevented by a high concentration of EDTA, the non-specific adsorption of oligonucleotide probes for FISH may also be avoided. Therefore, we examined the effectiveness of EDTA to reduce background signals caused by the non-specific binding of fluorescent oligonucleotide probes during hybridization.
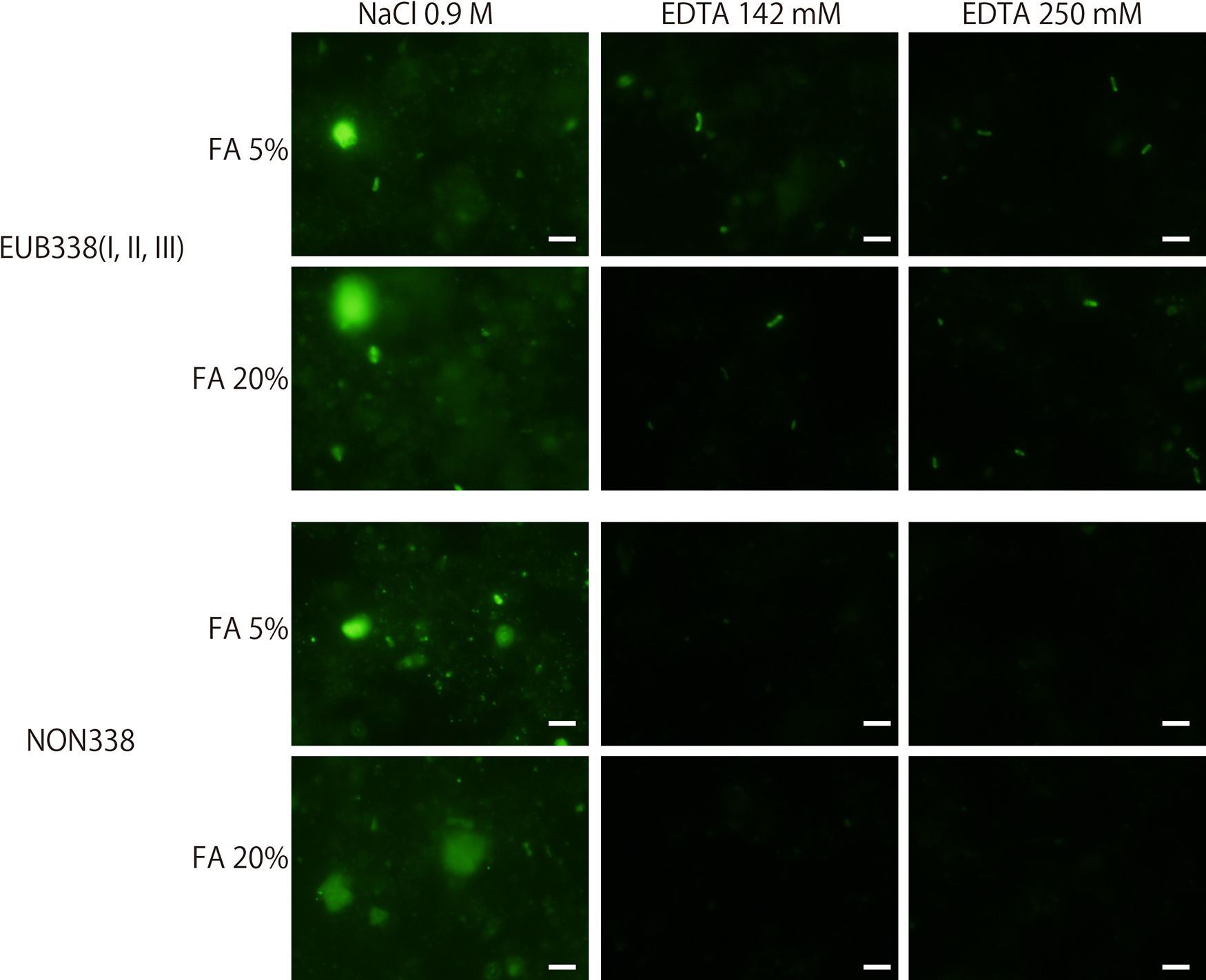
Three different buffer compositions were investigated: a standard FISH buffer composition (0.9 M NaCl, 20 mM Tris/HCl, 0.01% SDS, and 5–35% formamide) and buffers containing 142 or 250 mM EDTA instead of 0.9 M NaCl (designated as EDTA-FISH). An EDTA solution (pH 8.0) (generated using EDTA·2Na and NaOH for pH adjustments) contained approximately 750 mmols of Na+ for 250 mmol of EDTA ions. Since Na+ is the ion that affects the properties of hybridization, NaCl was omitted to avoid high Na+ concentrations in the hybridization buffer for EDTA-FISH. The marine subsurface sediment samples used in the present study were representative of typical organic-rich deep (sediment depth of 219 m) and ancient (ca. 460,000 years) sedimentary habitats in the northwestern Pacific Ocean off the Shimokita Peninsula of Japan, as described previously (Morono et al., 2011). Microbial activity was stimulated by the addition of methane to the headspace of the test tube and incubated at 10°C for 2 months. Samples were fixed with paraformaldehyde, washed with PBS, and preserved in EtOH/PBS (1:1) at –20°C until used. Sediment slurry was placed on a glass slide, embedded in low gelling point agarose (concentration 0.1%; MetaPhor, Lonza), dehydrated in an ethanol series (50, 80, and 99.5% ethanol), and then air-dried. A mixture of the EUB338 I, II, and III probes (Daims et al., 1999) labeled with Alexa Fluor 488 was used. Formamide concentrations ranged between 5 and 35%, and incubation temperatures and durations for the hybridization and washing steps were 46°C for 2 h and 48°C for 20 min, respectively. The NON338 probe (complimentary to EUB338 I) was used as a negative control. As shown in Fig. 1, fluorescence signals were obtained in hybridizations with both the EUB338 and NON338 probes after standard FISH, which clearly demonstrated that the non-specific adsorption of probes compromised the specific detection of microbes. On the other hand, EDTA-FISH effectively reduced the non-specific adsorption of the NON338 probe, while the intensity of signals from microbial cells was unchanged in the hybridization with the EUB338 probe. In EDTA-FISH, we used two different concentrations of EDTA (142 and 250 mM) to assess whether EDTA concentrations affect the adsorption of probes to sediment particles. Although the images shown in Fig. 1 did not appear to differ, a slightly lower background signal was observed with 250 mM EDTA by eye-based observations; therefore, 250 mM EDTA was subsequently used.

To evaluate the effects of EDTA on the hybridization properties of probes and specificity of FISH, we conducted experiments to investigate probe dissociation and mismatch discrimination. Dissociation curves for EDTA-FISH and standard FISH were investigated at various concentrations of formamide in both the hybridization and washing buffers. Hybridization and washing times were 3 h and 15 min, respectively. The EUB338 and GAM42a probes were evaluated using Escherichia coli cells. Fluorescent images were obtained under a fixed exposure time, and signal intensities were measured using daime software (Daims et al., 2006). The dissociation curves obtained by EDTA-FISH shifted to a lower formamide concentration, and the degrees of these shifts differed (Fig. 2a and b). Only a slight difference was observed for the GAM42a probe, whereas the EUB338 probe showed a marked change (approximately 10% formamide difference). Differences in Na+ concentrations may partially explain the shift observed in the optimum formamide concentration. The concentration of Na+ in hybridization buffer was approximately 0.75 M when 250 mM EDTA was used (EDTA·2Na and NaOH were used for pH adjustments), which was 0.15 M lower than that in standard FISH hybridization buffer. Theoretically, the decrease of 0.15 M in Na+ reduced the melting temperature of oligonucleotides by 1.3°C (=16.6 log [0.9/0.75], [Wilkinson, 1992]), which corresponded to approximately 3% of formamide (0.5°C/% formamide for a DNA-RNA hybrid [Wilkinson, 1992]). The present results clearly demonstrated that an optimum formamide concentration needs to be selected for EDTA-FISH probe-by-probe. Mismatch experiments, for which we selected two oligonucleotide probes, GAM42a (Manz et al., 1992) and SRB385 (Amann et al., 1990), were conducted to assess the effects of EDTA on mismatch discrimination. The GAM42a probe was used to evaluate single mismatch discrimination using paraformaldehyde-fixed E. coli (Gammaproteobacteria) and Comamonas testosteroni (Betaproteobacteria) pure cultures. The SRB385 probe was for two mismatch discrimination, and fixed pure cultures of Desulfovibrio vulgaris and Deinococcus radiodurans were used. The results shown in Fig. 3 demonstrated that EDTA-FISH distinguished two mismatches (Fig. 3c), whereas the discrimination of a single mismatch was not possible (Fig. 3a). The use of a competitor probe was necessary for single mismatch discrimination (Fig. 3b). These results are consistent with standard FISH (Manz et al., 1992). Thus, no apparent effect of using EDTA on the selectivity of FISH was observed. However, it is important to note that an optimum formamide concentration needs to be selected for each probe. Differences in Na+ concentrations between the standard FISH and EDTA-FISH hybridization buffers caused a shift in the concentration of formamide, which varied depending on the probes. Although the mechanisms responsible for differences in the degree of the formamide concentration shift have not yet been elucidated and are out of the scope of the present study, a reduction in the background adsorption of oligonucleotide probes was achieved without compromising any critical property of FISH probing.

In the present study, we examined the utility of EDTA for reducing the background signals of FISH caused by the non-specific adsorption of fluorescent oligonucleotide probes. The replacement of 0.9 M NaCl with 250 mM EDTA in the hybridization buffer effectively reduced the non-specific adsorption of probes, while the hybridization of probes to target molecules in microbial cells was maintained. Although slight differences were observed in hybridization characteristics and the optimization of hybridization conditions needs to be conducted probe-by-probe, the capacity for mismatch discrimination was unchanged in EDTA-FISH. This simple EDTA-FISH procedure has potential for expansion to reducing background signals in a more advanced FISH protocol, such as hybridization chain reaction-FISH (Yamaguchi et al., 2015), or other sensitized protocols of FISH in the future.
Citation
Morono, Y., Kubota, K., Tsukagoshi, D., and Terada, T. (2020) EDTA-FISH: A Simple and Effective Approach to Reduce Non-specific Adsorption of Probes in Fluorescence in situ Hybridization (FISH) for Environmental Samples. Microbes Environ 35: ME20062.
https://doi.org/10.1264/jsme2.ME20062
Acknowledgements
The authors thank the crew, technical staff, and shipboard scientists of the DV Chikyu for their help during the shakedown cruise CK06-06 in 2006. We thank S. Tanaka and S. Fukunaga for their technical assistance with this study. The present study was partly supported by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (no. 24687004, 15H05608, and 19H00730 to Y.M. and no. 18H01564 to K.K.)
References
- Amann, R.I., Binder, B.J., Olson, R.J., Chisholm, S.W., Devereux, R., and Stahl, D.A. (1990) Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl Environ Microbiol
56: 1919–1925.
- Amann, R.I., and Fuchs, B.M. (2008) Single-cell identification in microbial communities by improved fluorescence in situ hybridization techniques. Nat Rev Microbiol
6: 339–348.
- Bezanilla, M., Manne, S., Laney, D.E., Lyubchenko, Y.L., and Hansma, H.G. (1995) Adsorption of DNA to mica, silylated mica, and minerals: characterization by atomic force microscopy. Langmuir
11: 655–659.
- Daims, H., Bruhl, A., Amann, R., Schleifer, K.H., and Wagner, M. (1999) The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: development and evaluation of a more comprehensive probe set. Syst Appl Microbiol
22: 434–444.
- Daims, H., Lücker, S., and Wagner, M. (2006) daime, a novel image analysis program for microbial ecology and biofilm research. Environ Microbiol
8: 200–213.
- Dekas, A.E., and Orphan, V.J. (2011) Chapter twelve—Identification of diazotrophic microorganisms in marine sediment via fluorescence in situ hybridization coupled to nanoscale secondary ion mass spectrometry (FISH-NanoSIMS). In Methods in Enzymology vol. 486. Klotz, M.G. (ed.) Cambridge: Academic Press, pp. 281–305.
- Dianou, D., Ueno, C., Ogiso, T., Kimura, M., and Asakawa, S. (2012) Diversity of cultivable methane-oxidizing bacteria in microsites of a rice paddy field: Investigation by cultivation method and fluorescence in situ hybridization (FISH). Microbes Environ
27: 278–287.
- Furukawa, K., Hoshino, T., Tsuneda, S., and Inamori, Y. (2006) Comprehensive analysis of cell wall-permeabilizing conditions for highly sensitive fluorescence in situ hybridization. Microbes Environ
21: 227–234.
- Halary, S., Duperron, S., and Boudier, T. (2011) Direct image-based correlative microscopy technique for coupling identification and structural investigation of bacterial symbionts associated with metazoans. Appl Environ Microbiol
77: 4172–4179.
- Hirakata, Y., Oshiki, M., Kuroda, K., Hatamoto, M., Kubota, K., Yamaguchi, T., et al. (2015) Identification and detection of prokaryotic symbionts in the ciliate Metopus from anaerobic granular sludge. Microbes Environ
30: 335–338.
- Kawakami, S., Kubota, K., Imachi, H., Yamaguchi, T., Harada, H., and Ohashi, A. (2010) Detection of single copy genes by two-pass tyramide signal amplification fluorescence in situ hybridization (two-pass TSA-FISH) with single oligonucleotide probes. Microbes Environ
25: 15–21.
- Krsek, M., and Wellington, E.M.H. (1999) Comparison of different methods for the isolation and purification of total community DNA from soil. J Microbiol Methods
39: 1–16.
- Kubota, K. (2013) CARD-FISH for environmental microorganisms: Technical advancement and future applications. Microbes Environ
28: 3–12.
- Manz, W., Amann, R., Ludwig, W., Wagner, M., and Schleifer, K-H. (1992) Phylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria: Problems and solutions. Syst Appl Microbiol
15: 593–600.
- Morono, Y., Terada, T., Nishizawa, M., Ito, M., Hillion, F., Takahata, N., et al. (2011) Carbon and nitrogen assimilation in deep subseafloor microbial cells. Proc Natl Acad Sci U S A
108: 18295–18300.
- Ogram, A., Sayler, G.S., Gustin, D., and Lewis, R.J. (1988) DNA adsorption to soils and sediments. Environ Sci Technol
22: 982–984.
- Rai, H., Yokoyama, S., Hashimoto, S., Nakamura, S., and Hattori, H. (2010) A restriction factor of direct DNA extraction from volcanic ash soils. In Proceedings of the 19th World Congress of Soil Science; Soil Solutions for a Changing World. Gilkes, R., and Prakongkep, N. (eds). Brisbane, Australia. International Union of Soil Sciences.
- Saeki, K., Sakai, M., and Wada, S-I. (2010) DNA adsorption on synthetic and natural allophanes. Appl Clay Sci
50: 493–497.
- Wilkinson, D.G. (1992) The theory and practice of in situ hybridization. In In Situ Hybridization: A Practical Approach. Wilkinson, D.G. (ed.) Oxford: IRL Press at Oxford University Press, pp. 1–13.
- Yamaguchi, T., Fuchs, B.M., Amann, R., Kawakami, S., Kubota, K., Hatamoto, M., et al. (2015) Rapid and sensitive identification of marine bacteria by an improved in situ DNA hybridization chain reaction (quickHCR-FISH). Syst Appl Microbiol
38: 400–405.