Original Article
Relationship Between Distribution and Habitat of Theligonum japonica in Sado Island,Japan
2024 Volume 76 Issue 3 Pages 93-105
Details
2024 Volume 76 Issue 3 Pages 93-105
ヤマトグサはアカネ科の多年生草本で,日本では秋田県から熊本県にかけての広域に隔離分布している。谷沿いの湿った林内に生育することが知られているが,どのような環境に隔離分布するかはわかっていない。本研究では生育地の環境条件の特徴を見出すことを目的とし,佐渡島大佐渡山地と小佐渡山地での分布調査と気象観測および栽培試験を行った。佐渡島を約1km四方メッシュに区切り,森林率が高い381メッシュを踏査した結果,102メッシュでヤマトグサの生育を確認した。分類樹木分析の結果,夏季の日照時間が少なく,夏季の降水量が少ないメッシュに,ヤマトグサの生育が多いことが示された。気象観測では,観測地点のほとんどで夏季を通じて空中湿度が高く,実際に霧が観測された。栽培試験では,空中湿度を高く保った条件で盛んな発根が見られた。ヤマトグサの個体群の存続には,霧によって空中湿度が高く保たれる生育の核となるエリアが必要で,その周辺へはシュート伸長による栄養繁殖で広がると推測された。日本全国でみると,夏季に上昇霧が発生し,空中湿度が高く維持される立地が限られることにより,ヤマトグサは隔離分布していると考えられる。
Theligonum japonica Okubo et Makino is a perennial herb that belongs to the family Rubiaceae. It is distributed in scattered patches from Akita to Kumamoto Prefectures in Japan. It is known to grow in moist forests, but few studies have revealed what environmental preferences are responsible for its disjunct distribution. This study conducted distribution surveys, meteorological observations on Sado Island, and cultivation tests to clarify the environmental conditions under which this species grows. The study area on Sado Island was divided into approximately 1 km square meshes, and 381 meshes were surveyed. We found T. japonica within 102 meshes. The result of classification tree analysis showed that T. japonica grows mostly in meshes with shorter daylight hours in summer and lower precipitation in summer influence. Meteorological observations showed that air humidity was high throughout the summer, and fog was observed more frequently at most observation sites. Cultivation tests showed vigorous rooting under conditions of higher air humidity. It is suggested that the population of T. japonica requires core areas where the air humidity is kept high by fog and that this species spreads to the surrounding area through vegetative propagation. In the Japanese archipelago, there are few places where aerial humidity is kept higher by fog in summer; therefore, T. japonica is likely distributed disjunctly.
ヤマトグサTheligonum japonica Okubo et Makinoは,日本と中国に分布するアカネ科ヤマトグサ属の多年生草本である。ヤマトグサ属は北アフリカ,地中海地方,アナトリア,中国西南部から日本に4 種が分布し,日本ではヤマトグサ1種がみられる(内貴, 2017)。日本のヤマトグサは秋田県から熊本県にかけての24府県の日本海側,太平洋側の山地に点在しており(ANONYMOUS, 2021)(第1図),著しい隔離分布をしている。隔離分布する植物としては,高山植物(藤井, 2008)や,超塩基性岩にだけ生育する植物(北村, 1950)などが有名であるが,ヤマトグサのような地理的分布をする植物は他にみあたらず,どのような環境に分布が規定されているのかわからない。
ヤマトグサは,南限の熊本県では暖温帯上部に,北限の秋田県では冷温帯に生育しており,本種の気温に対する適応範囲は比較的広いと考えられる。また,ヤマトグサの生育地の地質を概観すると,堆積岩類が多い傾向はあるものの,火山岩,花崗岩,変成岩などにも生育しており,石灰岩や蛇紋岩のような特殊な地質に生育地が限定されていることはない。分布が稀なことから17の府県でレッドデータブックに掲載されており(野生動物調査協会・Envision環境保全事務所, 2021),そこには本種が森林の湿った林床に生育していることが記載されている(岡山県, 2003; 茨城県生活環境部環境政策課, 2013; 秋田県生活環境部自然保護課, 2014; 岐阜県環境生活政策課, 2015; 三重県農林水産部みどり共生推進課, 2015; 山梨県森林環境部みどり自然課, 2018; 熊本県希少野生動植物検討委員会, 2019; 和歌山県環境生活部環境政策局環境生活総務課自然環境室, 2022)。また,鈴木・田崎(1993)は筑波山において,林緑の湿った立地に生育していることを報告している。Flora of China(Chen and Funston, 2011)には,ヤマトグサの生育地は,谷間の湿地,渓流沿いの標高900~1,200 mと記載されている。つまり,ヤマトグサは,温帯域に広く生育し,特殊な地質に分布が偏ることはなく,湿潤な林床や渓流沿いという,日本中にいくらでもある立地環境に生育するという報告のみで,隔離分布の原因はわからない。
ヤマトグサ以外の植物で,生育地の水分条件の影響で隔離分布する植物としては,河川沿いに生育するユビソヤナギSalix hukaoana Kimura(指村ほか, 2010),湿地に生育するハナノキAcer pycnanthum K. Koch(植田, 1989),雲霧帯に生育するレッドウッドSequoia sempervirens D. Don (Ewing et al., 2009),海霧が発生する場所に分布が偏るエゾヒョウタンボクLonicera alpigena L. subsp. glehnii (F. Schmidt) H. Hara(指村, 2020)などがあり,水分条件といっても,土壌水分だけでなく,霧など空中湿度が影響している例も散見される。
そこで,本研究は,ヤマトグサの生育が多くみられる佐渡島で(石沢, 1982)ヤマトグサの分布調査を行い,水分条件に注目して,生育地の環境条件の特徴を見出すことを目的とした。また,ヤマトグサの生育に霧が関係している可能性を検討するため,生育地で気象観測を行った。さらに,ヤマトグサが,高い空中湿度にどのように適応しているかを明らかにするため,空中湿度を変えた容器の中で栽培試験を行った。以上から,ヤマトグサの生育を可能にする環境要因について考察し,日本国内での隔離分布の原因について推察した。

(ANONYMOUS, 2021)に基づくヤマトグサの日本における分布(●)
佐渡島は北緯約38度に位置し,本州から約32 kmの日本海上に位置する面積855 km2の離島である(第2図A)。佐渡島の地形は,北西側の大佐渡山地と南東側の小佐渡山地,およびそれらに挟まれた国中平野に区分される。主山稜は北東から南西方向に延びており,大佐渡山地は金北山(標高1,172 m)を主峰とし,標高700~1,000 m 級の険しい山脈が縦走するのに対して,小佐渡山地は全般的に標高が低く,大地山(おおちやま)646 mなど標高300~600 mのなだらかな小~中起伏山地となっている(瀬沼, 1981)。佐渡島の年平均気温は,低地では12.7~13.7℃の範囲内にあり,島内で最も標高の高い金北山山頂では5.4℃となっている。年降水量は約1,600~1,900 mmで,新潟県において最も少ない地域である。国中平野における年最深積雪量は約20~40 cmであるが,山間地においては増加し,小佐渡山地で約1 m,大佐渡山地では2~3 mである。また大佐渡山地の北西の海沿いは冬季季節風に直面し,風速がきわめて強い(瀬沼, 1981)。大佐渡山地はスギ原生林が広がる地域があり,暖候期のうち多くは霧でおおわれている(崎尾, 2021)。

Aには佐渡島の地形および気象観測地点(●)を示し,Bには各気象観測地点の地形図を示した。
佐渡島を全国標準地域メッシュ(3次メッシュ,1辺約1 km)に区切り,各メッシュで以下の調査を行った。まず,ヤマトグサの生育位置を記録するため,各メッシュを横切る道路および登山道沿いでヤマトグサを探した。踏査ルートの距離は,メッシュによりまちまちとなった。ヤマトグサは山地の森林内に生育することが既往研究より明らかであったので,平野部などで森林率が50%未満のメッシュは踏査対象外とした。ヤマトグサを発見した場合,生育地点の緯度経度をスマートフォン用アプリGeographica (iOS版 ver. 1.3.23)またはスーパー地形(iOS版 ver. 4.9.4)を用いて記録した。使用するスマートフォンはいずれも衛星みちびきに対応している機種を用いたため,位置精度は安定的に約10 m程度であった。この分布情報はArcGIS pro ver. 2.9で整理した。これら調査は,なるべく多くのメッシュで行うように努力した。
ヤマトグサの生育地点の気象を観測するため,佐渡島のヤマトグサ生育地4 か所(アオネバ,北五十里,ひょうたん池,新穂)(第2図B)に,温湿度計とインターバルカメラを設置して,冬季を除く約1年間に相当する期間(2021年8月10日~10月21日,2022年5月14日~8月22日)の観測を行った。アオネバとひょうたん池は山頂凹地,北五十里は谷底,新穂は緩斜面に位置する。インターバルカメラで撮影した写真は目視により,霧,霧不詳(霧かどうか判断がつかない),霧なし,レンズ曇り,白とびの5 カテゴリーに分けて集計した。
ヤマトグサの空中湿度に対する成長反応を観察するため,2021年8月17日に採集したヤマトグサを3 種類の条件(蓋を開けた容器,容器に穴をあけた容器,蓋を閉じた容器)に5 個体ずつ移植して,常温の室内に置いた。容器は光の差し込む窓辺に置いたが,処理により光条件が異ならないよう,窓から等距離に並べた。培地には湿らせた腐葉土と水苔を用いた。その後,2021年9月10日までおよそ4日おきに根の数と長さを測定した。
3. 解析方法佐渡島内の両山地におけるヤマトグサの分布と環境条件の関係を解析するため,ヤマトグサの生育を確認した地点を在地点,生育が確認できなかったメッシュでは,その踏査ルートから,メッシュの重心に最も近い点を不在地点として,分類樹木分析を行った。なお,メッシュ内の踏査ルートが1,000 m未満の不在メッシュは解析から除外した。分類樹木分析で用いる環境要因には気候と地形に関する要因を用いた。気候要因として,国土数値情報の平年値メッシュデータ(国土交通省, 2021)から,各月の日照時間,降水量および平均気温を引用し,要因ごとに主成分分析で第一主成分(pc 1)と第二主成分(pc 2)に集約した。地形要因として,数値標高モデル(10 mメッシュ)(国土地理院, 2021)を50 mのメッシュ内で平均し,水分条件の指標としてよく用いられる曲率(凸地形か凹地形かの指標),SPI(河川の流れの強さの指標)および TWI (水のたまりやすさの指標)をArcGIS pro ver. 2.9 で算出した。すなわち,分類樹木分析に用いた環境要因は月日照時間pc 1, 月日照時間pc 2, 月降水量pc 1, 月降水量pc 2, 月平均気温pc 1, 月平均気温pc 2, 曲率, SPI, TWI の9 要因である。これらの要因については,VIF(Variance Inflation Factor)が10以下であり,多重共線性が存在する可能性は低いことを確認した。分類樹木分析の結果で分岐に使われた要因について,各Nodeに振り分けられたデータセットを用い,在不在に対する箱ひげ図を作成した。分析には統計解析ソフトR ver. 4.0.3 (R Core Team, 2020)のライブラリrpart (Therneau and Atkinson, 2019)を用いた。
気象観測については,1週間ごとに各カテゴリーの割合をグラフにして傾向を検討した。栽培試験に関しては,観察日時点の根の本数と長さの平均をグラフにして,条件ごとに比較した。
調査地である佐渡島全域を標準地域メッシュ(第3次メッシュ)(1辺約1 km)で区切ると959 メッシュとなり,森林が50%以上の面積を占めるメッシュは665メッシュであった。そのうちの381メッシュを踏査し,102メッシュ(約27%)でヤマトグサの生育を確認した(第3図)。生育地の森林は,スギ(Cryptomeria japonica (L.f.) D. Don),シラキ(Neoshirakia japonica (Siebold et Zucc.) Esser),ナナカマド(Sorbus commixta Hedl.),オニグルミ(Juglans mandshurica Maxim. var. sachalinensis (Komatsu) Kitam.),ミズキ (Cornus controversa Hemsl. var. controversa),ミズナラ (Quercus crispula Blume var. crispula)など,様々な樹木が優占していた。
生育が確認できなかったメッシュは279メッシュで,これらのうち踏査ルートが1,000 m以上のメッシュは179であった。この179メッシュを不在地点とし,在地点(102メッシュ)と合わせて分類樹木分析を行った。結果として得られた樹状図と分岐に使われた要因の在不在に対する箱ひげ図を第4図に示す。樹状図の正解率(Accuracy)は0.81であり,初めの分岐(Node 1)で月日照時間の第一主成分(月日照時間pc 1)が85.88より小さいところで,ヤマトグサの生育する割合が高いことが示された。つづく分岐(Node 3)では,月降水量の第二主成分(月降水量pc 2)が−22.935 以上のところで,ヤマトグサの生育する割合がさらに高まった(Node 5)。
月日照時間pc 1は,夏季(7月)の日照時間と相関が高かった(第1表)。月降水量pc 2は,冬季(10月から1月)の降水量と正の,夏季(6月,7月)の降水量と負の相関があった(第2表)。
第5図は調査地4 地点の秋季と春季の温湿度変化の結果を表したものである。気象観測の結果,どの生育地も夏から秋にかけて80~90%を超える高い空中湿度を維持していることがほとんどであった。このほか,インターバルカメラに映った写真から,新穂では2021年9月から10月にかけて高頻度で霧が観測された(第6図)。アオネバでは2021年8月から9月にかけて3割から6割程度の写真で霧が観測された。ひょうたん池ではカメラの設置場所が光の当たる場所であったため白とびの写真が多くなってしまったものの,2021年9月から10月,2022年6月に霧が観測された。これらとは異なり,北五十里ではほとんど霧が観測されなかった。
容器内へ移植したヤマトグサの栽培試験の結果,蓋を開けた容器ではまったく発根が見られなかったのに対して,それ以外の2処理では節から根を盛んに出していた(第7図)。蓋を閉じた容器と穴をあけた容器では,最初の2週間はどちらも同程度の発根が見られたが,1カ月近く経過すると,蓋を閉じた容器の方で数多くの根が観察された。

(第3次メッシュ)およびヤマトグサ生育確認メッシュ
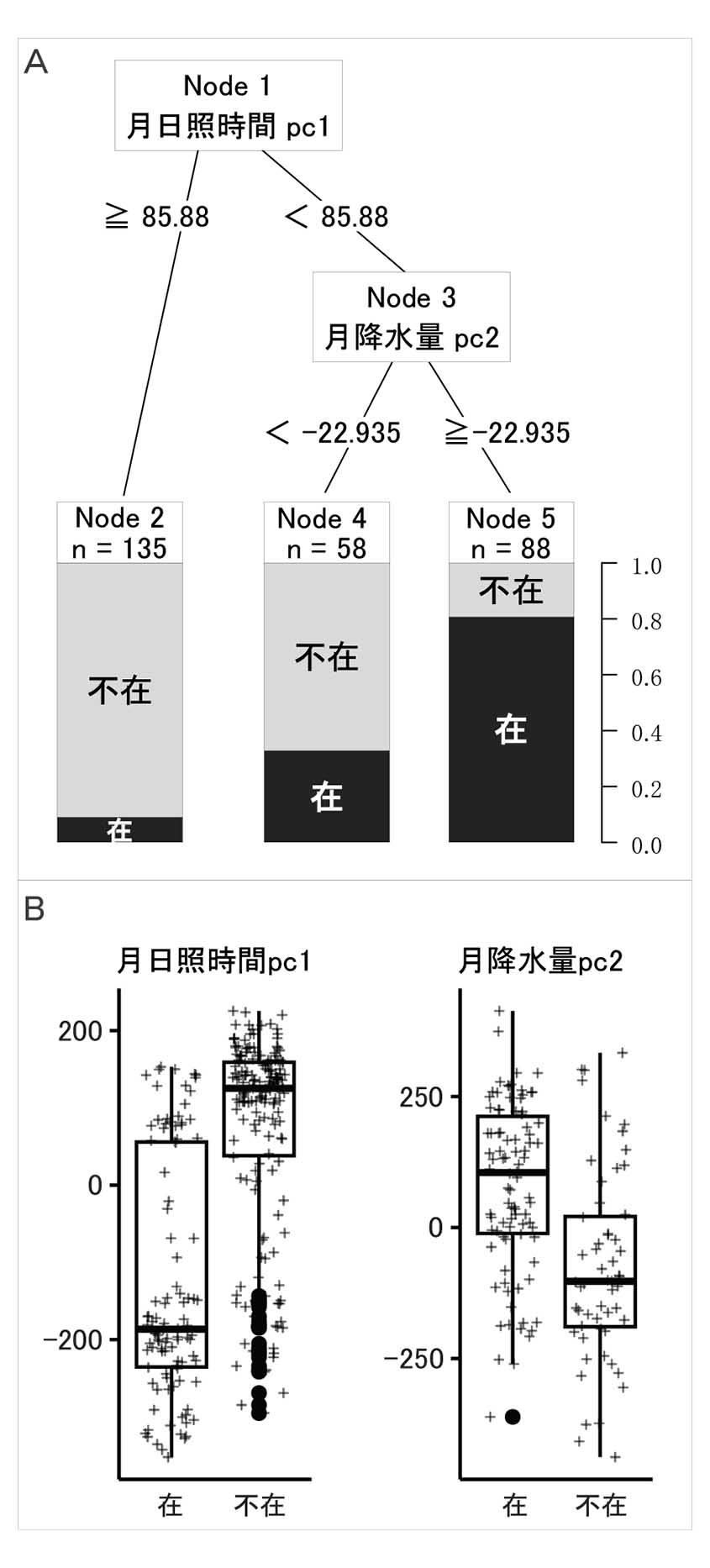
Aには樹状図を示す。下段右の0.0~1.0は在不在の割合である。Bには樹状図の分岐で使われた要因の在不在に対する箱ひげ図を示す。
| 1月 | 0.56 |
| 2月 | 0.50 |
| 3月 | 0.71 |
| 4月 | 0.82 |
| 5月 | −0.17 |
| 6月 | −0.06 |
| 7月 | 0.97 |
| 8月 | 0.08 |
| 9月 | 0.84 |
| 10月 | 0.10 |
| 11月 | −0.16 |
| 12月 | 0.16 |
| 1月 | 0.65 |
| 2月 | −0.02 |
| 3月 | −0.02 |
| 4月 | 0.10 |
| 5月 | −0.07 |
| 6月 | −0.32 |
| 7月 | −0.17 |
| 8月 | 0.24 |
| 9月 | 0.08 |
| 10月 | 0.59 |
| 11月 | 0.81 |
| 12月 | 0.83 |



各条件5個体を栽培し,観察日時点の根の本数と長さを測定した。縦線は各測点の標準誤差を表す。
本研究により,佐渡島内でのヤマトグサの分布の偏りと,分布に影響する環境要因が明らかになった。ヤマトグサはこれまで佐渡島のいたるところで生育が見られるとされてきた(石沢, 1982)。本調査により,実際に広範囲に高頻度で生育を確認することができたが,大佐渡山地では小佐渡山地より生育地が多いなど,分布には偏りがあった(第3図)。このような分布の偏りがどのような環境条件の影響によるかを調べるために行った分類樹木分析では,夏季の日照時間と相関の高い月日照時間pc 1がより小さいメッシュに,ヤマトグサの生育する地点が多いことが示された(第4図)。分岐に使われた要因の在不在に対する箱ひげ図で見ても,月日照時間pc 1の値の小さい方に在地点が偏っていることがわかる。このことは,ヤマトグサの生育地では,夏季に晴れ間が少ないことを示唆している。さらにそれらの中でも,月降水量pc 2が高いメッシュにヤマトグサの生育する地点が多くなっていた。月降水量pc 2は冬季の降水量と正の相関が高いが,冬季の降水量は降雪量と同義であり,冬季は雪の下で休眠中のヤマトグサに影響は少ないと考えられるため,夏季の降水量との負の相関に着目すると,夏季の降水量が少ないメッシュにヤマトグサは偏って生育していたと考えることができる。これらから,ヤマトグサの生育地の日照時間の少なさは夏の降雨のためではないことがうかがえる。
佐渡島は日本海に浮かぶ日本最大の離島で,島の半分を占める大佐度山地は,暖候期の多くの時間を霧で覆われる“霧の山”とされている(河島・伊豫部, 2010)。大佐渡山地に位置する新潟大学佐渡演習林で行われた霧の観測によると,6月から9月までの4カ月間のうち7 割程度の日に上昇霧が観測されており,月別にみると7月の霧発生日数が最も多い(河島・伊豫部, 2010)。河島・伊豫部(2010)は,霧発生日の約80%が気象擾乱を伴わない天気図型であり,このとき降水量は少ないと述べていることからも,本研究でヤマトグサの分布に影響した日照時間の短さも,降雨のためではなく,霧が原因である可能性が高い。本研究でも,分類樹木分析の結果で夏季の降水量は少ない方がヤマトグサの生育が多いと考えられた(第4図)。また,河島・伊豫部(2010)は,この霧は夏に海から供給される水蒸気が山を上がることで発生する上昇霧であると述べている。大佐渡山地の標高500 mから上は雲霧帯的な気象となり,全体の過半をスギやヒノキアスナロが優占する針葉樹林および針広混交林が占める(本間, 2018)。佐渡島は,佐渡ジオパークに指定されており,その見どころのひとつとしてスギの原生林がある(相田, 2022)。これらから,佐渡島の高標高域にかかる上昇霧は,佐渡島の植物相に大きな影響を与えており,ヤマトグサもそのひとつであると考えることができる。実際,インターバルカメラでは霧が観測された地点が多かった(第2図,第6図)。また,この霧は小佐渡山地(ひょうたん池,新穂)でも観測された。小佐渡山地でも山の上の方は夏季に霧に覆われることが影響して,ヤマトグサが生育している可能性が高い。
ヤマトグサの生育地のうち,北五十里ではインターバルカメラで霧が撮影されることが少なかったが,湿度計の値は80~90%と高かった(第5図,第6図)。このことから,霧が少なくても,ヤマトグサは川沿いなどで空中湿度が高く維持される場所ならば,ある程度生育が可能と考えられた。
栽培試験において,ヤマトグサは,蓋を閉じた容器と穴をあけた容器の中で,盛んに発根していた(第7図)。これらの容器内の湿度は計測していないが,容器内側に結露した水滴が認められたことから,容器内の湿度は高く維持されていたことがわかる。生育地においても,シュートを長く伸ばして,地面に倒れ,節から根を出し,這うように生育している様子がよく見られた(第8図)。ヤマトグサは,空中湿度の高いところでは,この根からの吸水によって,長く伸ばしたシュートを維持することができ,栄養繁殖によって個体群を広げていると考えられる。また,佐渡島での観察では,ヤマトグサはどの生育地でもよく開花していたが,結実が見られる場所はとても少なかった。夏季の結実期にも踏査を行ったが,果実を確認できた生育地はアオネバと新潟大学佐渡演習林のそれぞれ1か所ずつだけであった。これらから,佐渡島のヤマトグサは種子繁殖よりも,栄養繁殖に依存して個体群維持を行っており,そのため,高い空中湿度のもとで節から発根できるような場所に分布が偏ると考えられた。
本研究の結果と佐渡島の生育地での観察から,大佐渡山地および小佐渡山地の山頂付近では,夏季に霧がかかることによって空中湿度が高く保たれ,そこにヤマトグサが栄養繁殖とまれな種子繁殖により,広く生育していると考えられる。また,佐渡島島内の下流付近の川沿いでは,霧のかかる山頂部から栄養繁殖によって分布を広げ,植物体の一部が川を流下して定着したヤマトグサが,点々と生育しているのではないだろうか。つまり,ヤマトグサの長期間にわたる個体群維持には,霧によって空中湿度が高く保たれる生育の核となるエリアが必要で,その周辺の川沿いへはシュート伸長による栄養繁殖で広がることができると推測された。これらを確かめるためには,佐渡島内で詳細なサンプリングを行い,遺伝分析による集団解析を行う必要がある。
ここで日本全国へ視野を広げてみると,川沿いで空中湿度の高い立地環境は日本中に多数あると考えられる。夏季に霧が継続的に発生する場所を第9図に示す(気象庁, 2023)。この図は全国144 の観測地点のデータから作成されたものなので,解像度が低く,ヤマトグサの分布を説明するスケールのものではないが,全国的に,夏季に霧が継続的に発生する場所は多くないことがうかがえる。全国のヤマトグサの分布を概観すると,海に近い大きな山塊の周辺に分布が集中しているように見える(第1図)。実際に生育地のひとつである高野山は,平家物語にも「花の色は林霧の底にほころび」と記述されているほどで,古くから霧がかかることで有名な山であった。佐渡島以外においても,ヤマトグサの生育に霧が影響している場合がどの程度あるのか,そのことが隔離分布と関係しているのか,調査する必要がある。
その存立に霧が大きく影響していると考えられている米カリフォルニアのレッドウッドの森林には,高い空中湿度が地史的時間スケールで維持されてきたことによって遺存している植物が多く生育しているとされる(Ewing et al., 2009)。また,エゾヒョウタンボクは,海霧が発生する場所や風穴地に分布が限定されており,東北地方での隔離分布は氷期以降の遺存であるとされる(指村, 2020)。ヤマトグサの個体群維持に霧の発生が重要である可能性を鑑みて,その隔離分布の背景を考察するには,レッドウッド林やエゾヒョウタンボクの例と同様に地史的背景も考慮に入れる必要があるだろう。今後は,日本列島の中でヤマトグサの隔離分布が生じた環境条件と地史的背景の関係について明らかにしていくことが課題となる。

a:シュートを長く伸ばす。b:這うように生育する。c:節から発根する。

気象庁(2023)より1991年~2020年の観測データから作成した。
佐渡島内でのヤマトグサの分布は,夏季に霧の出る場所に偏っていた。一部では,霧は出ていなくても川沿いの空中湿度の高い場所にもヤマトグサの生育がみられた。ヤマトグサは,高い空中湿度のもとでは節から発根し,この根からの吸水によって,長く伸ばしたシュートを維持することができると考えられる。ヤマトグサの個体群維持には,霧によって空中湿度が高く保たれる生育の核となるエリアが必要で,その周辺の川沿いへはシュート伸長による栄養繁殖で広がれることが推測された。日本全国のヤマトグサの分布についても,海に近い大きな山塊の周辺に隔離していることから,霧が影響している可能性が大きい。
本研究を実行するにあたって,日本自然環境専門学校の学生には現地調査をお手伝いいただいた。清水長正氏には本原稿作成に際して貴重なコメントをいただいた。中野剛氏には標本をお送りいただいた。なお,本研究は第19回こしじ水と緑の会・朝日酒造自然保護助成基金から助成を受けたものである。