REGULAR ARTICLE
Characteristics of the Localization of Connexin 43 in Satellite Cells during Skeletal Muscle Regeneration In Vivo
2015 Volume 48 Issue 2 Pages 53-60

Details
2015 Volume 48 Issue 2 Pages 53-60
For myogenesis, new myotubes are formed by the fusion of differentiated myoblasts. In the sequence of events for myotube formation, intercellular communication through gap junctions composed of connexin 43 (Cx43) plays critical roles in regulating the alignment and fusion of myoblasts in advances of myotube formation in vitro. On the other hand, the relationship between the expression patterns of Cx43 and the process of myotube formation in satellite cells during muscle regeneration in vivo remains poorly understood. The present study investigated the relationship between Cx43 and satellite cells in muscle regeneration in vivo. The expression of Cx43 was detected in skeletal muscles on day 1 post-muscle injury, but not in control muscles. Interestingly, the expression of Cx43 was not localized on the inside of the basement membrane of myofibers in the regenerating muscles. Moreover, although the clusters of differentiated satellite cells, which represent a more advanced stage of myotube formation, were observed on the inside of the basement membrane of myofibers in regenerating muscles, the expression of Cx43 was not localized in the clusters of these satellite cells. Therefore, in the present study, it was suggested that Cx43 may not directly contribute to muscle regeneration via satellite cells.
In mammalian adult skeletal muscles, myogenic stem cells, which are called muscle satellite cells, are located between the basement membrane and the plasma membrane in myofibers [20]. When skeletal muscles are damaged by crushing, injection of myotoxic agents and disease, satellite cells are sequentially activated, proliferate and differentiate and then the differentiated satellite cells align and fuse with each other and existing myofibers to repair the damaged myofibers [1, 9, 29, 31]. In this repair process due to satellite cells, the aggregation and fusion of differentiated satellite cells play important roles in promoting subsequent myotube formation. Although the aggregation and fusion of differentiated satellite cells are regulated by diverse factors (e.g. M-cadherin, nuclear cell adhesion molecular, calcium, beta1-integrin, calpain, etc.) [10, 16, 18], connexin 43 (Cx43) is included as one of these factors.
Cx43 is one of the factors composing gap junctions, which are the membrane channels formed in adjacent cells that allow the intercellular passage of small-molecular-mass molecules, secondary messengers and ionic signals [8, 26]. It was indicated that Cx43-containing gap junctions appear in cultured myoblasts at the time of differentiation, and subsequently disappear in the myotubes in vitro [2, 4, 12]. Conversely, the blockade of Cx43-containing gap junctional channels inhibited the formation of myotubes in vitro [25]. Moreover, in Cx43-deficient myoblasts, the aggregation and subsequent fusion of differentiated myoblasts were inhibited, and consequent delay of the formation of myotubes was induced in vitro [3, 14]. In addition, it was reported that the deletion of Cx43 induced decreases of the expression of MyoD, myogenin and creatine kinase activity in cultured myoblasts [3]. Therefore, it was conjectured that Cx43-containing gap junctional intercellular communication is closely related to the regulation of the sequence of the myotube formation process, including differentiation, aggregation and fusion, in myoblasts for myogenesis in vitro [13, 24].
In general, Cx43-containing gap junctions are mainly localized in adult heart but not in adult skeletal muscle in vivo [7, 11]. However, it was indicated that the expression of Cx43 mRNA and protein is distributed throughout regenerating muscles [3, 12, 30]. Moreover, it was indicated that, in regenerating muscles, the expression of Cx43 reached peak levels in the phase in which both the down-regulation of Ki-67 (a marker molecule for proliferation) and the up-regulation of p21 (a marker molecule for cell cycle arrest/differentiation) were concomitantly induced [12]. On the other hand, it was reported that, in Cx43-deficient mice, muscle regeneration was significantly delayed after muscle trauma [3]. These previous studies suggest that Cx43 may be functionally involved in the regulation of muscle regeneration. However, because previous studies did not identify satellite cells definitely using specific molecular markers (e.g. M-cadherin and Pax7), the relationship between Cx43 and satellite cell remain poorly understood in muscle regeneration in vivo. Therefore, the present study investigated the relationship between Cx43 and satellite cells in regenerating skeletal muscles in vivo, and demonstrated the possibility that Cx43 may be not directly involved in regulation of muscle regeneration derived from satellite cells.
Female Fischer 344 rats (8 weeks of age, n=5/group) were used in the present study. They were housed in individual cages at 22°C under a 12:12-hr light-dark cycle and were provided with food and water ad libitum. All experiments were carried out with the approval of Aichi University of Education Animal Ethics Committee.
Induction of skeletal muscle regenerationTibialis anterior (TA) muscles were surgically exposed under pentobarbital sodium anesthesia (60 mg/kg, i.p.). A narrow gauge (27G×3/4", 0.4 mm in diameter and 19 mm in length) needle was inserted into both the distal and proximal ends of TA. Then, in right TA muscles, 0.2 ml of 0.5% bupivacaine hydrochloride (BPVC; Marcain) in 0.9% saline was injected into each end (0.4 ml in total). The contralateral TA muscle of each animal was injected with 0.4 ml of 0.9% saline in the same manner, as a control. The incision area was sutured and disinfected. The rats were sacrificed on day 1, 3, 5 and 7 post-BPVC injection, and then the TA muscles were obtained. The samples were quickly frozen in liquid nitrogen and stored at −80°C until use.
Histological evaluation of muscle degeneration/regeneration induced by BPVC injectionFor the histological evaluation of muscle degeneration/regeneration induced by BPVC injection, serial cross sections (10 μm thickness) of mid-belly muscles were cut using a cryostat (CM1850, Leica, Germany) at −20°C and thawed on 3-amino propylethoxysilane-coated slides. Then, these sections were visualized by hematoxylin and eosin (HE) staining.
ImmunostainingThe primary antibodies used in the present study were as follows: rabbit polyclonal anti-connexin 43 (1:400, Zymed Laboratories, South San Francisco, CA, USA) [5, 12, 13], rabbit polyclonal anti-Ki67 (a marker molecule for proliferation) [27] (1:400, Novacastra Laboratories, Newcastle, UK), rabbit polyclonal anti-myogenin (a marker molecule for differentiation of satellite cells) [28] (1:100, Santa Cruz Biotech, Santa Cruz, CA, USA), rabbit polyclonal anti-laminin (a marker molecule for basement membrane) (1:3000, Sigma, St. Louis, MO, USA), mouse monoclonal anti-M-cadherin (a marker molecule for satellite cells) [17] (1:400, BD PharMingen, San Diego, CA, USA) and mouse monoclonal anti-laminin (1:200, Chemicon, Temecula, CA, USA).
For immunostaining, serial cross sections (10 μm thickness) of mid-belly muscles were cut using a cryostat at −20°C and thawed on 3-amino propylethoxysilane-coated slides. The sections were fixed with 4% paraformaldehyde in 0.1 M phosphate-buffered (pH 7.4) for 15 min and washed with 0.1 M phosphate buffer saline (PBS, pH 7.4). Then, to block any nonspecific immunoreactivity, the sections were incubated with 0.1 M PBS containing 10% normal serum and 1% Triton X-100 for 1 hr. The primary antibodies were diluted with 0.1 M PBS containing 5% normal serum and 0.3% Triton X-100, and the secondary antibodies were diluted with 0.1 M PBS containing 5% normal serum and 0.1% Triton X-100.
For multicolor immunofluorescence staining, the sections were simultaneously incubated for 16–48 hr at 4°C with the primary antibodies. Then, they were washed in 0.1 M PBS, and simultaneously incubated overnight at 4°C with secondary antibodies. The secondary antibodies used in the present study were fluorescein-labeled horse anti-mouse IgG (1:300, Vector Laboratories, Burlingame, CA, USA) and rhodamine-labeled goat anti-rabbit IgG (1:300, Chemicon). Then, the sections were washed in 0.1 M PBS and mounted in Vectashield mounting medium with DAPI (Vector Laboratories) to visualize the nuclei. All image analyses were performed using a fluorescence microscope (BX51; Olympus, Tokyo, Japan) and SR image (Cool SNAPcf, Roper Scientific, Chiba, Japan).
To evaluate whether BPVC properly induced skeletal muscle damage and subsequent muscle regeneration, in the present study, morphological observation using HE stain was performed (Fig. 1). By day 7 after BPVC injection, a sequence of muscle degeneration/regeneration events, such as hypercontraction, emptying, necrosis, phagocytosis of myofibers, myotube formation and the development of myofibers with central nuclei, were observed. These morphological findings were consistent with previous reports [9, 22, 23], and indicate that the BPVC injection performed in the present study was sufficient to induce muscle degeneration/regeneration.

Histological changes in TA muscles in control (A), 1 (B), 3 (C), 5 (D) and 7 (E) day post-BPVC injection. These findings indicate that muscle degeneration/regeneration was induced by BPVC injection. Cross sections were stained with hematoxylin and eosin. Bar=50 μm.
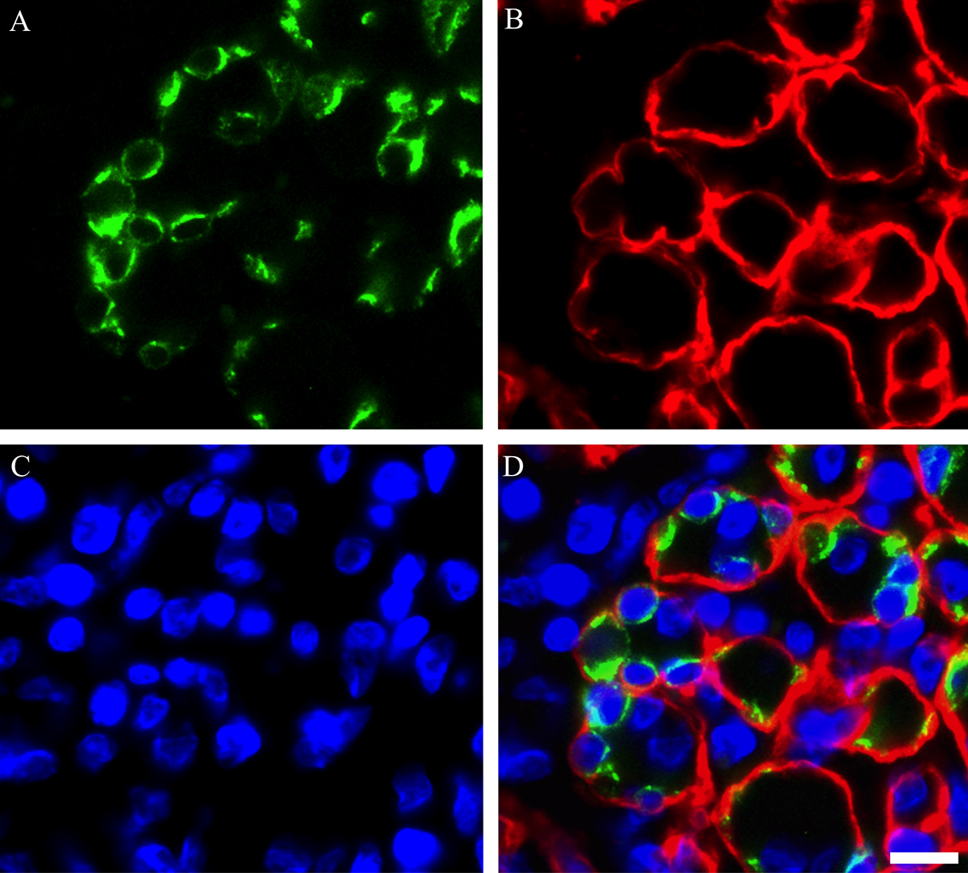
To confirm whether the expression of Cx43 was detected in adult rat skeletal muscles, we performed immunostaining by using Cx43 antibody on contralateral control, and on day 1, 3, 5 and 7 post-BPVC injection (Fig. 2A–E). Cx43-positive immunoreactivity was detected in skeletal muscles after BPVC injection, but not control muscles. Moreover, Cx43-positive immunoreactivity was observed as a punctate staining pattern throughout the regenerating skeletal muscles in the present study. These characteristics of the expression patterns of Cx43 were consistent with a previous study that investigated Cx43 expression in regenerating skeletal muscle in vivo [12]. Next, to evaluate the localization of Cx43 expressed in regenerating muscles, double fluorescence immunostaining using laminin and Cx43 antibodies was performed on day 3 post-BPVC injection (Fig. 3A–D). The Cx43 immunoreactivity was not detected on the inside of the laminin-positive basement membranes. This finding was consistently detected throughout the experimental period. These results indicate that the expression of Cx43 was induced by muscle damage, and its localization was on the outside of myofibers.
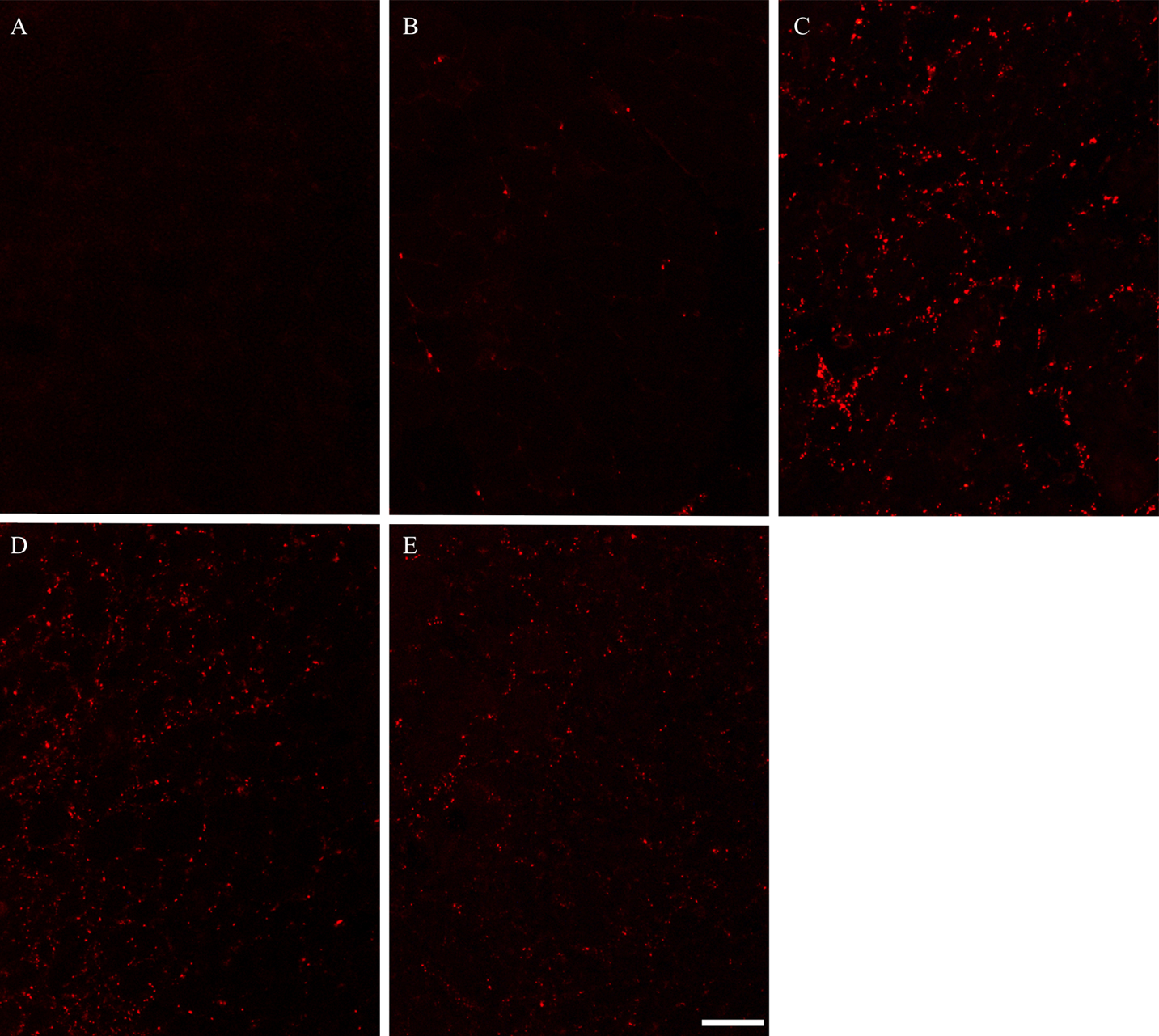
Photomicrographs showing the expression pattern of Cx43 in control and regenerating TA muscles. Immunostaining was used to visualize the localization of Cx43 in TA muscles in control (A), 1 (B), 3 (C), 5 (D) and 7 (E) day post-BPVC injection. Cx43-positive immunoreactivity was detected in BPVC-injected muscles, but not in control muscles. Bar=50 μm.
Photomicrographs showing the localization of Cx43 in TA muscle on day 3 post-BPVC injection. Double immunostaining was used to visualize the localization of laminin (A), Cx43 (B) and DAPI (C), and these images were merged (D). These findings indicate that Cx43-positive immunoreactivity was detected on the outside of the laminin-positive basement membrane of myofibers in regenerating muscles. These findings were consistently detected throughout the experimental period. Bar=10 μm.
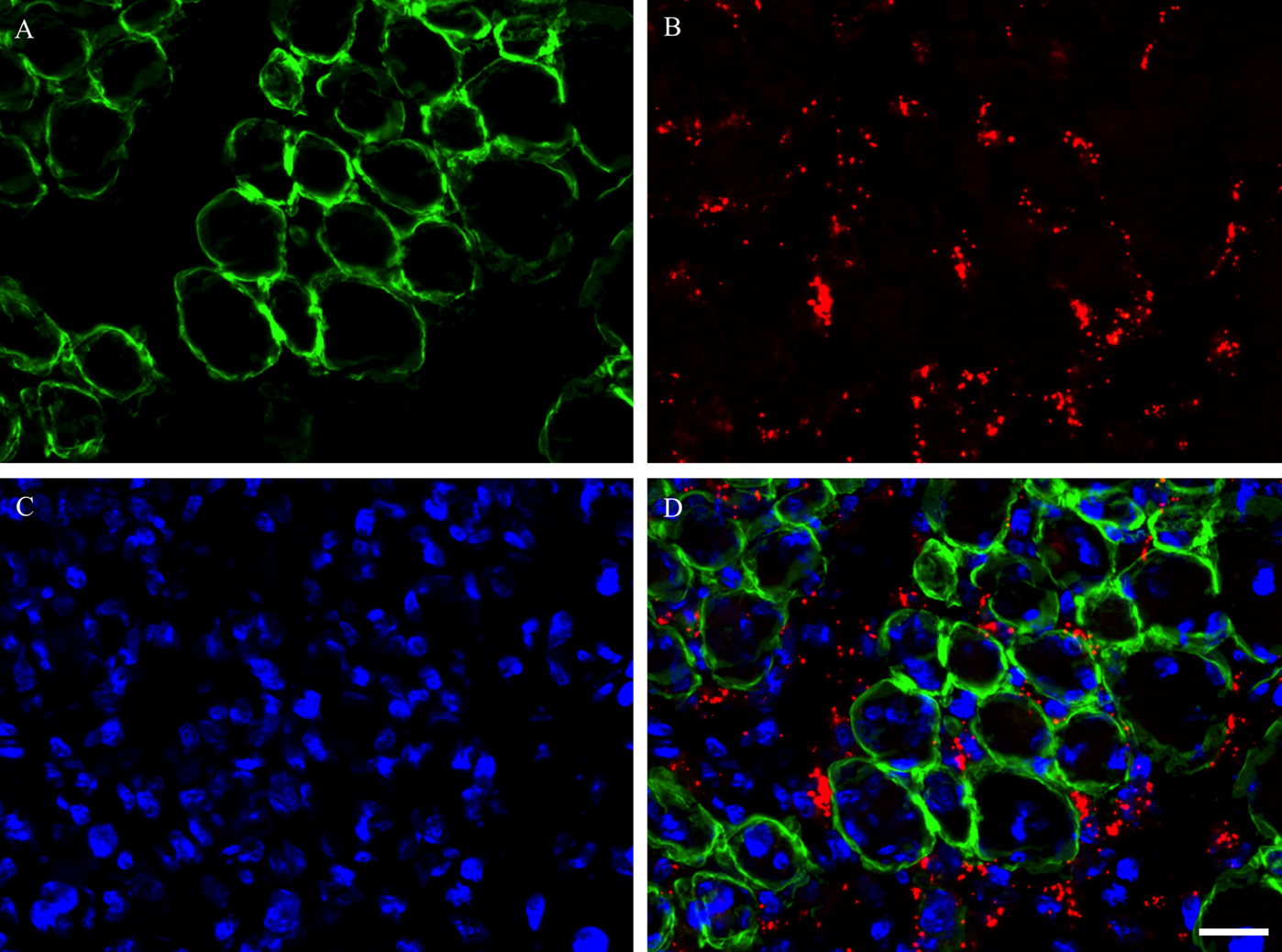
During degeneration and subsequent regeneration of skeletal muscles, satellite cells transition from the proliferation phase to the differentiation phase, and then the process of fusion with each other or existing myofibers progresses to repair myofibers completely [15]. In the present study, triple immunostaining using M-cadherin, laminin antibodies and DAPI was performed on day 5 post-BPVC injection (Fig. 4A–D). As a result, the clusters of M-cadherin-positive satellite cells were shown to align and closely appose as a ring-like structure detected on the inside of the laminin-positive basement membrane. Next, triple immunostaining using Ki-67, M-cadherin antibodies and DAPI on day 1 post-BPVC injection (Fig. 5A–D), and myogenin, M-cadherin antibodies and DAPI was performed on day 5 post-BPVC injection (Fig. 6A–D). While Ki-67-expressing satellite cells were located individually, myogenin-positive satellite cells were arranged as a ring-like cluster. These findings indicate that the arrangement of differentiating satellite cells was structurally different from that of proliferating satellite cells in regenerating muscles.
Photomicrographs showing the localization of the cluster of satellite cells in TA muscle on day 5 post-BPVC injection. Triple immunostaining was used to visualize the localization of M-cadherin (A), laminin (B) and nuclei (C), and the three images were merged (D). These findings indicate that the cluster of M-cadherin-positive satellite cells localized on the inside of the laminin-positive basement membrane of myofibers in regenerating muscles. Bar=10 μm.
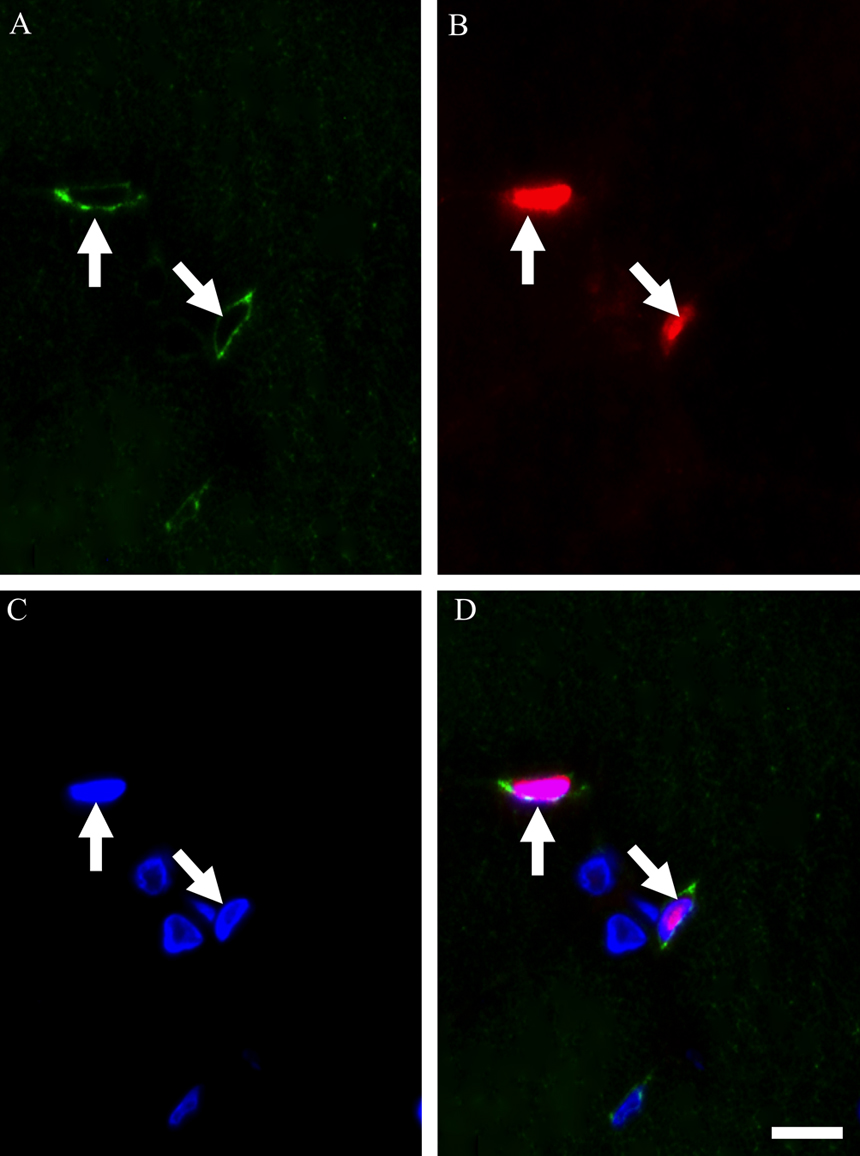
Photomicrographs showing the localization of Ki-67 in satellite cell nuclei in TA muscle on day 1 post-BPVC injection. Triple immunostaining was used to visualize the localization of M-cadherin (A), Ki-67 (B) and nuclei (C), and the three images were merged (D). Arrows (A–D) indicate a Ki-67-positive satellite cell localized in regenerating muscles. Bar=10 μm.
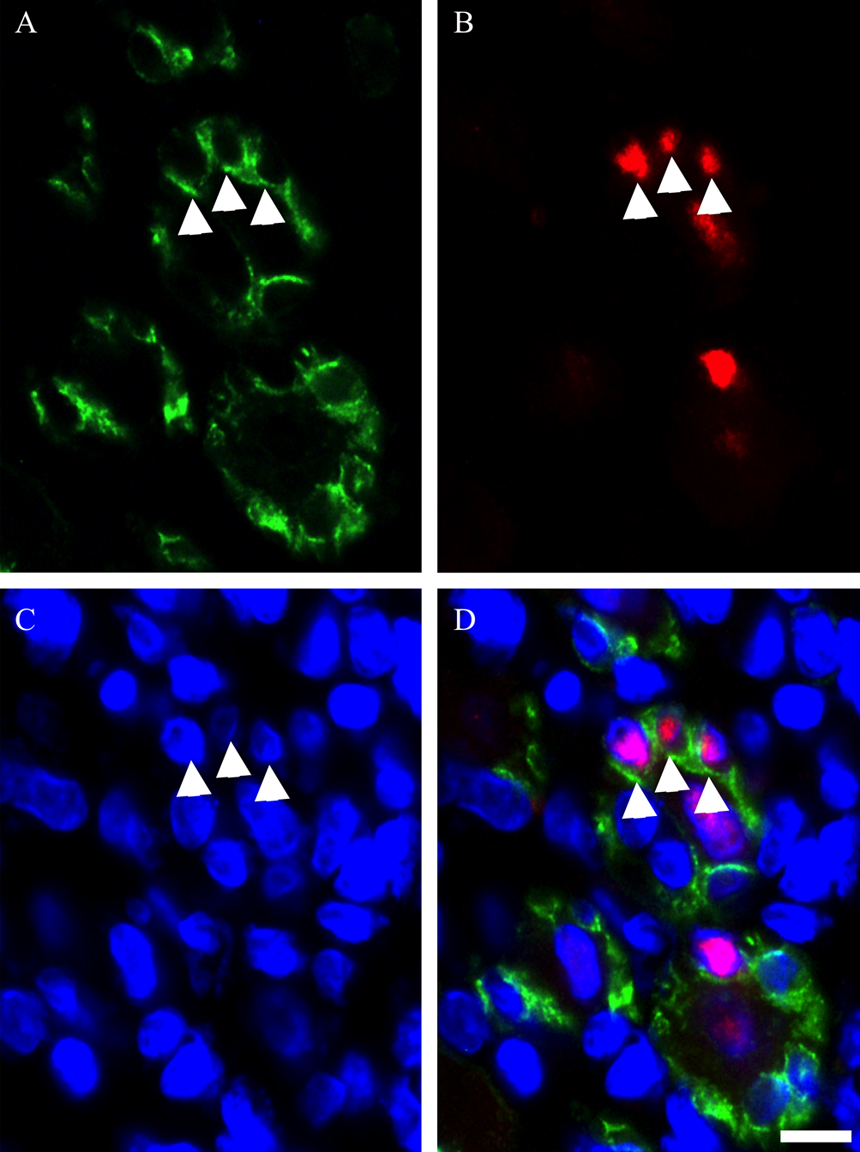
Photomicrographs showing the localization of myogenin in satellite cell nuclei in TA muscle on day 5 post-BPVC injection. Triple immunostaining was used to visualize the localization of M-cadherin (A), myogenin (B) and nuclei (C), and the three images were merged (D). Arrowheads (A–D) indicate the myogenin-positive satellite cells arranged as a ring-like structure in regenerating muscles. Bar=10 μm.
To evaluate the relationship between Cx43 expression and satellite cell clusters in regenerating muscles, double immunostaining using Cx43 and M-cadherin antibodies was performed on day 5 post-BPVC injection (Fig. 7A–D). It was observed that Cx43-positive immunoreactivity did not colocalize with M-cadherin-positive immunoreactivity. These findings indicate that the expression of Cx43 was not induced on satellite cell clusters in regenerating muscles. These findings were consistently detected throughout the experimental period.
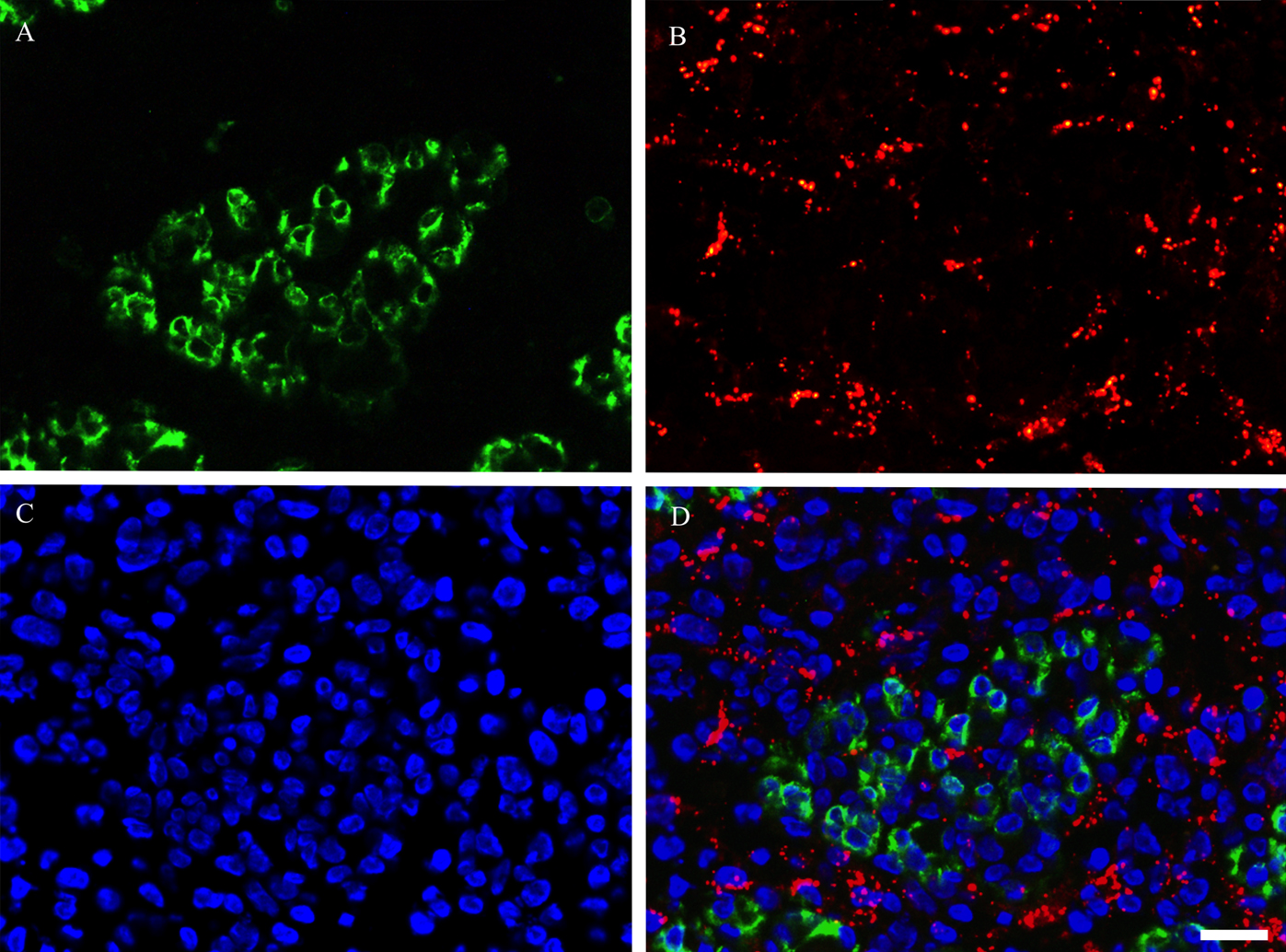
Photomicrographs showing the positional relationship between M-cadherin-positive satellite cell and Cx43 in TA muscle on day 5 post-BPVC injection. Double immunostaining was used to visualize the localization of M-cadherin (A), Cx43 (B) and DAPI (C), and the three images were merged (D). These findings indicate that Cx43-positive immunoreactivity did not colocalize with M-cadherin-positive immunoreactivity in regenerating muscles. These findings were consistently detected throughout the experimental period. Bar=20 μm.
Gap junctions consisting of Cx43 play important roles in the regulation of myoblast fusion and myotube formation during myogenesis in vitro [2, 4, 25]. The present study characterized the Cx43 expression pattern in regenerating rat adult skeletal muscles induced by BPVC injection. Our findings indicate that Cx43 was expressed on the outside of the basement membrane of myofibers in regenerating muscles. Moreover, it was indicated that Cx43 was not expressed in the clusters of satellite cells in regenerating muscles. These findings suggest that Cx43 may not be directly involved in the repair mechanisms of myofibers via satellite cells in muscle regeneration.
In mammalian adult striated muscles, gap junctions consisting of Cx43 are absent in adult skeletal muscles, although they are present in cardiac muscles constitutively [7, 11]. Non-expression of Cx43 in control muscles as indicated in the present study is consistent with these previous findings. On the other hand, in the present study, the expression of Cx43 was induced in regenerating muscles. In particular, higher intensity of Cx43-positive immunoreactivity was detected on day 3 and 5 post-BPVC injection compared with that on day 1. These expression patterns of Cx43 were consistent with the expression pattern indicated in previous studies that investigated the Cx43 expression in regenerating skeletal muscle in vivo [2, 12]. Therefore, it was indicated that muscle damage is one of the triggers that induce the expression of Cx43 in adult skeletal muscles in vivo, although there may also be other physiological stimuli that can induce its expression that remain to be clarified.
The localization of Cx43 expressed in regenerating muscles remains poorly understood. The present study examined the relationship between Cx43 and myofibers in regenerating muscles using immunostaining with Cx43 and laminin antibodies, and indicated that Cx43 expression was not localized on the inside of the laminin-positive basement membrane of myofibers. These findings suggest that the expression of Cx43 may not be regulated in the myofibers in regenerating muscles. In addition, although the present study was not able to determine whether the expression of Cx43 was induced in the Ki-67-positive proliferating satellite cells due to the technical limitation of multicolor immunostaining, because of the localization of Cx43 expression outside of myofibers, it was suggested that the expression of Cx43 was not induced in the proliferating satellite cells in muscle regeneration.
In regenerating post-muscle damage, differentiated satellite cells fuse with each other and existing myofibers to repair damaged myofibers via the formation of myotubes [10, 21, 29, 31]. In the present study, it was indicated that the clusters of M-cadherin-positive satellite cells arranged as a ring-like structure were localized on the inside of the basement membrane of myofibers, and simultaneously it was indicated that the myogenin-expressing satellite cells formed ring-like cell clusters. The present study could not determine whether the clusters of myogenin-expressing satellite cells were localized on the inside of the basement membrane of myofibers. However, it was previously reported that differentiated satellite cells arranged as a ring-like structure were localized on the inside of the basement membrane of myofibers, and that these clusters represented an earlier stage of the sequence of events of myotube formation in muscle regeneration [19]. The findings obtained in the present study were remarkably similar to the findings indicated in this previous study. Therefore, it was suggested in the present study that these clusters may be at the aggregation and fusion phase of differentiated satellite cells, representing a more advanced stage of myotube formation.
It was indicated that intracellular communication via gap junctions consisting of Cx43 plays critical roles in regulating differentiation and fusion in myoblasts during myogenesis in vitro [2–4, 14, 25]. However, in the present study, Cx43 was not expressed on the satellite cell clusters formed on the inside of the basement membrane of myofibers in the regenerating muscles. This finding is consistent with a previous report that indicated that Cx43 mRNA was not expressed in aligned myogenin-positive cell clusters in regenerating muscles [30]. Moreover, it was indicated that Cx43 mRNA was mainly expressed in macrophages in regenerating muscles [3, 30]. In addition, it was indicated that Cx39 mRNA was expressed in clusters of myogenin-positive myogenic cells in regenerating muscles in vivo, and that Cx39 may be involved in the regulation of satellite cell aggregation and fusion followed by myotube formation in the regenerating muscles [6, 30]. Therefore, in the present study, unlike myogenesis in vitro, it was suggested that Cx43 may not directly play a functional role in regulating the aggregation and fusion of satellite cell clusters in regenerating muscles in vivo. On the other hand, it was indicated that the expression of Cx43 was down-regulated when myoblasts were beginning to form myotubes [13]. Moreover, it was reported that the half-life of Cx43 was about 1.3 hours in adult rat heart [5]. Therefore, the present study cannot rule out the possibility that transient Cx43 expression occurs in the satellite cell clusters in stages preceding myotube formation based on our immunohistochemical analysis. Further studies are needed to elucidate the regulatory mechanisms of the expression and function of Cx43 in activated satellite cells in muscle regeneration.
In conclusion, the present study demonstrated that the expression of Cx43 was induced by muscle trauma, and that the expressed Cx43 was not localized on the inside of the basement membrane of myofibers in regenerating muscles. Moreover, it was indicated that Cx43 was not expressed in the clusters of satellite cells in regenerating muscles. These findings suggest that Cx43 may not be directly involved in the repair mechanisms of myofibers via satellite cells in muscle regeneration.
This work was supported by a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (B; project no. 21300236, to N. Kasuga).