Original Articles
Single-nucleotide polymorphism analyses on ABCC11, EDAR, FGFR2, and ABO genotypes of mummified people of the Joseon Dynasty, South Korea
2018 年 126 巻 2 号 p. 67-73

詳細
2018 年 126 巻 2 号 p. 67-73
Although previous studies have demonstrated successful single-nucleotide polymorphism (SNP) genotyping of modern samples, the potential applicability of this methodology to ancient human specimens has not been confirmed. With regard in particular to the SNPs in the ABCC11, EDAR, FGFR2, and ABO genes, all of which are commonly analyzed in biomedical research, only a relatively limited number of papers on ancient specimens are currently available. We thus studied the SNP genotypes in the ABCC11, EDAR, FGFR2, and ABO genes of mummies from the Joseon Dynasty of Korea. Those SNP genotypes in brain samples (n = 5) were determined using multiplex single-base extension (SBE) primers in polymerase chain reaction (PCR) analyses of each gene locus. SNP analysis revealed the mummies’ ABCC11 genotype was revealed to be 538AA (dry-type earwax and low risk for axillary osmidrosis). In the EDAR and FGFR2 genes, the variant alleles rs3827760-CC (EDAR) and rs4752566-TT (FGFR2), indicative of thick and straight hair, were identified. In addition, the ABO genotypes BO02 (SN1-2), O01O02 (Sapgyo), AO01 (Hadong2), BB (Yongin), and O02O02 (SN PK) were identified. Our SNP genotyping of Korean mummies provided us with specific insight into the potential of this methodology for application to the analysis of ancient human specimens. This study fills a gap in our knowledge of the use of SNP genotyping in forensic medicine by proving that it can help to reveal the physical traits of ancient individuals.
Single-nucleotide polymorphism (SNP), the most common type of sequence variation in the human genome, is known to be highly associated with susceptibility to several diseases as well as an individual’s response to a specific medicine (Shastry, 2002). Although sequence variation alone does not adequately predict disease risk, the identification of the SNP pattern in a specific disease can provide researchers with a good starting point from which to uncover its genetic background (Shastry, 2002).
SNP analysis is useful not only for revealing the pathogenetic aspects of health and disease. As many SNP variations diverge by human population group, forensic scientists around the world have identified relevant SNP information from crime-scene samples to assist law-enforcement authorities with their identification of suspects in criminal investigations (Fujimoto et al., 2008b; MacLean and Lamparello, 2014). Indeed, recent progress in SNP analysis has enabled the prediction of physical traits by which individuals differ from each other. Underlying such traits are, among others, the ABCC11, EDAR, FGFR2 and ABO genotypes.
The SNP located in the ABCC11 gene is revealed to be genetically related to earwax type (Yoshiura et al., 2006). Recently, light has also been shed on the novel function of the ATP-binding cassette (ABC) transporter in the apocrine glands. The ABCC11 gene is thus associated with axillary osmidrosis, colostrum secretion, as well as the potential risk of mastopathy (Ishikawa et al., 2012). Hair morphology, a highly divergent phenotype among human populations, is closely correlated with the SNPs in the ectodysplasin A receptor (EDAR) and fibroblast growth factor receptor 2 (FGFR2) genes, which are the genetic determinants of hair thickness among Asian populations (Fujimoto et al., 2008a, b, 2009). The shovel shape of the upper incisors, a common characteristic in Asian and Amerindian populations, is also known to be genetically determined by the EDAR gene (Kimura et al., 2009). FGFR2, meanwhile, is one of the major causative genes of genetic disorders implicated in craniosynostosis (Ko, 2016) and breast cancer (André and Cortés, 2015).
The ABO blood type is another trait widely analyzed in human genetics. The ABO blood group comprises three different alleles: the co-dominant A and B alleles, and one recessive O allele. Singly or in combination, they form the four different phenotypes A, B, AB, and O (Mourant, 1954; Lalueza-Fox et al., 2008). Advancements in ABO genotyping enable the prediction of an individual’s blood type based on the outcome of genetic analysis. ABO genotypes are especially useful in forensic or archaeological cases, because they make it possible to determine blood type without the need for serological analysis (Watanabe et al., 2013).
Certainly, previous studies have efficiently demonstrated the scientific applicability of SNP genotyping to the analysis of modern individual samples. However, insufficient reports are available to determine the potential of this methodology for the analysis of ancient human specimens. In fact, with respect to the SNPs in the ABCC11, EDAR, FGFR2, and ABO genes, only a very limited number of papers on ancient specimens obtained at archaeological sites are currently available (Lalueza-Fox et al., 2008; Kim et al., 2011; Keller et al., 2012; Ambers et al., 2016).
In this regard, we should emphasize the importance of Joseon Dynasty (1392–1910 CE) mummies as an essential resource for scientific research on biological traits of ancient people. The relatively high state of preservation of most Joseon mummies after centuries-long interments, especially when considering the lack of any formalized embalming techniques at that time, is astounding (Song and Shin, 2014). For the past decade, we have been actively engaged in maintaining Joseon mummy specimens for use in investigations into the physical traits of pre-20th-century Korean populations. In the current study, we analyzed the SNPs in the ABCC11, EDAR, FGFR2, and ABO genes of Korean mummies. The results provide specific information on the effectiveness and potential of SNP analyses for revealing anthropological information on ancient populations.
The archaeological and anthropological information on the Korean mummies is summarized in Table 1. The mummies were recovered from Joseon Dynasty tombs (Figure 1) estimated to date from the 16th to 18th centuries based on archaeological and historical evidence, family pedigrees, and tree-ring test results. Brain specimens of the mummies (n = 5) were the specific materials analyzed in this study. All were collected in the course of dissection and stored in a freezer until use. This study was exempted from review by the Institutional Review Board (IRB) of Seoul National University Hospital (IRB No. 2017-001).
| Cases | Estimated date | Location discovered | Sex | Year of discovery |
|---|---|---|---|---|
| SN1-2 | 1605–1733b | Seoul | Male | 2007 |
| Sapgyo | 16Ca | Chungcheongnam-do | Male | 2011 |
| Hadong2 | 16C–early 17Cc | Gyeongsangnam-do | Female | 2009 |
| Yongin | 15C–16Ca | Gyeonggi-do | Female | 2005 |
| SN PK | 16C–18Ca | Seoul | Male | 2007 |

An example of a Joseon mummy (Sapgyo) used in this study as a subject.
After obtaining the brain-tissue samples, their surfaces were scraped using a sterilized knife and then exposed to 20 min UV irradiation. The samples (0.1–0.2 g) and extraction negative controls (ENCs) were subsequently incubated in 1 mL of lysis buffer (50 mM EDTA, pH 8.0; 1 mg/mL of proteinase K; 1% SDS; 0.1 M DTT) at 56°C for 24 h. Total DNA was extracted with an equal volume of phenol/chloroform/isoamyl alcohol (25:24:1) and then treated with chloroform/isoamyl alcohol (24:1). DNA isolation and purification were performed using a QIAmp PCR purification kit (Qiagen, Hilden, Germany). The purified DNA was then eluted in 50 μl of EB buffer (Qiagen) (Oh et al., 2015a, b).
SNP genotypingTen microliters of extracted DNA was treated with 1 unit of uracil-DNA-glycosylase (New England Biolabs, Hitchin, UK) for 30 min at 37°C. We performed SNP genotyping for the ABCC11, EDAR, FGFR2, and ABO genes. The information on the primer sets used in this study is summarized in Table 2. First, PCRs were performed to detect an SNP (rs17822931; nucleotide position 538) in ABCC11 (Yoshiura et al., 2006; Nakano et al., 2009), an SNP (rs3827760; nucleotide position 1540) in EDAR (Fujimoto et al., 2008b), an SNP (rs4752566) in intron 9 of FGFR2 (Fujimoto et al., 2009), and five SNPs (nucleotide positions 261, 771, 796, 802, and 803) in exons 6 and 7 of the ABO locus (Yamamoto, 2004; Kim et al., 2011).
| Primer | Primer sequence (5′→3′) | Product size (bp) | Annealing temperature (°C) | Primer reference | ||
|---|---|---|---|---|---|---|
| ABO | PCR | Exon 6F | CTC CAT GTG CAG TAG GAA GGA | 72 | 63 | Kim et al., 2011 |
| Exon 6R | AAT GTG CCC TCC CAG ACA A | |||||
| Exon 7F | CCA GTC CCA GGC CTA CAT C | 84 | ||||
| Exon 7R | TGC AYC TCT TGC ACC GAC | |||||
| SBE | ABO261 | AGG AAG GAT GTC CTC GTG GT | 21 | 50 | ||
| ABO771 | (T)13-GTC CCA GGC CTA CAT CCC | 32 | ||||
| ABO796 | (T)16-GGA CGA GGG CGA TTT CTA CTA C | 39 | ||||
| ABO802 | (T)24-GGC GAT TTC TAC TAC ATG GGG | 46 | ||||
| (T)25-GGC GAT TTC TAC TAC CTG GGG | 47 | |||||
| ABO803 | (T)36-CAC CGA CCC CCC GAA GAA C | 56 | ||||
| ABCC11 | PCR | EW-ampF | CTT CTG GGC ATC TGC TTC TG | 68 | 56 | Nakano et al., 2009 |
| EW-ampR | CAA ACC TCA CCA AGT CTG CCA | |||||
| SBE | SBE-EW | (T)14-CAT TGC CAG TGT ACT C | 31 | 50 | This study | |
| EDAR | PCR | EDAR-F | CCA CGG AGC TGC CAT TTG AT | 127 | 56 | |
| EDAR-R | TCA TCC CTC TTC AGG CCG AA | |||||
| SBE | SBE-EDAR | (T)15-GCG CCA CGT TTT CAC A | 32 | 50 | ||
| FGFR2 | PCR | FGFR2-F | TAA AGG TCT TGG GCA TCC AC | 115 | 56 | |
| FGFR2-R | GGG TTA TCT TCC TGG TGG CAC | |||||
| SBE | SBE-FGFR2 | (T)15-ACA ACC CCG AAT ATA CCA AAA T | 38 | 50 |
Briefly, PCR amplification was performed in a 20 μl reaction volume containing 40 ng of template DNA and ENCs, AmpliTaq Gold 360 Master Mix (Life Technologies, Carlsbad, CA), and appropriate concentrations of each primer. Thermal cycling was conducted on a SimpliAmp Thermal Cycler (Applied Biosystems, Waltham, MA): pre-denaturation at 94°C for 10 min; 40 cycles of denaturation at 94°C for 30 s; annealing at 56 or 63°C for 1 min; extension at 72°C for 30 s; and a final extension at 72°C for 10 min. To purify the PCR products, 5 μl was treated with 1 μl of ExoSAP-IT (catalogue no. 78201; USB Corporation, Cleveland, OH) at 37°C for 45 min. After that, the enzyme was inactivated by incubation at 80°C for 15 min.
Single-base extension (SBE) reactions subsequently were carried out using nine SBE primers and the SNaPshot kit (Applied Biosystems) according to the manufacturer’s instructions. The thermal cycling conditions for the SBE were as follows: 20 cycles of denaturation at 96°C for 10 s; annealing at 50°C for 5 s; and extension at 60°C for 30 s. The SBE was performed using the SimpliAmp Thermal Cycler (Applied Biosystems). For post-SBE treatment, the reaction mixtures were mixed with 1.0 unit of shrimp alkaline phosphatase (SAP), incubated at 37°C for 45 min, and subjected to heat inactivation at 80°C for 15 min. The reactants were analyzed using an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, Waltham, USA) and GeneMapper ID software, v. 3.2.1 (Applied Biosystems). The peak-height threshold for the SNaPshot analysis was set at 100 relative fluorescence units (RFUs). If two or more peaks were observed, the peak height observed below 15% of the main peak was excluded as non-allelic. For authentication of the results, two separate reactions were repeated for each mummified brain-tissue sample.
Precautions against contaminationDuring the sampling and lab work, protective gloves, masks, gowns, and head caps were consistently worn. Our ancient DNA (aDNA) lab facilities had been set up in accordance with the standards outlined in previous reports (Hofreiter et al., 2001; Willerslev and Cooper, 2005). The respective aDNA extraction and PCR preparation rooms were physically separated from our main PCR lab and equipped with night UV irradiation, isolated ventilation, and a laminated flow hood. Other procedures designed for authentic aDNA analysis (Hofreiter et al., 2001; Willerslev and Cooper, 2005) also were followed by us.
The ABCC11 genotype of all the mummies (n = 5) was revealed to be 538A in this study (Figure 2). This means that their earwax was the dry type and that their risks for axillary osmidrosis were very low. The SNPs in the EDAR (rs3827760) and FGFR2 (rs4752566) genes significantly associated with hair thickness of East Asians and Amerindians (Fujimoto et al., 2008a, b, 2009) also were studied. In the SNP analysis of the EDAR gene, the variant allele rs3827760-C was obtained from four mummies (SN1-2, Hadong2, Yongin, and SN PK) (Figure 2); the analysis of Sapgyo mummy failed. The pattern was also identified in variant alleles of rs4752566 in the FGFR2 gene. As was the case for the SNP in the EDAR gene, rs4752566-T was discovered in every Korean mummy (n = 5). The variant alleles of the EDAR and FGRF2 genes revealed that every Korean mummy in this study had the physical traits of thick and straight hair.

SNP genotyping of ABCC11, EDAR, and FGFR2 genes for Joseon mummies by a SNaPshot kit. (A) SN1-2, (B) Sapgyo, (C) Hadong2, (D) Yongin, (E) SN PK, and (F) Extraction negative control. SNP genotypes could be acquired in every mummy examined for ABCC11 (538A) and FGFR2 (rs4752566-T). SNPs in EDAR gene (rs3827760-C) were only obtained from four mummies. ND, not detected. Colors of the peaks: A, green; T, red; G, blue; C, black.
We identified the ABO blood genotypes of the five Joseon mummies by analysis of the five SNPs (nucleotide positions 261, 771, 796, 802, and 803) in two separate PCRs on exons 6 and 7 of the ABO locus as viewed on an electropherogram (Figure 3). Heterozygotic peaks were observed at these nucleotide positions in several mummies (SN1-2: 261, 771, 796, 803; Sapgyo: 771; Hadong2: 261). The extra peaks at the positions were confirmed to be more than 25% of the main peaks, on which basis they were determined not to be stutter peaks (Figure 3).
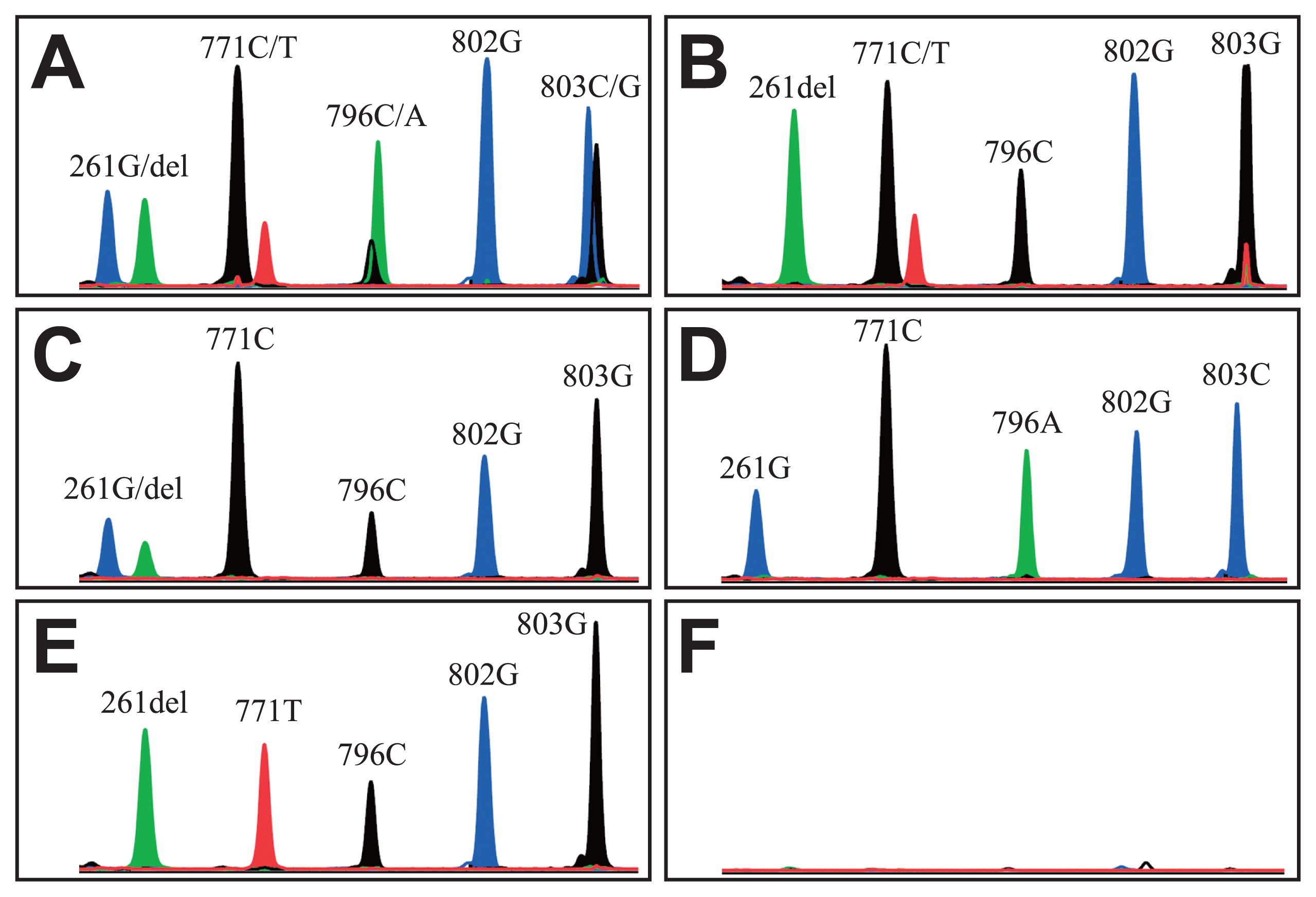
SNP analysis for ABO blood genotyping of Joseon mummies by a SNaPshot kit. Blood types obtained were: (A) SN1-2, BO02; (B) Sapgyo, O01O02; (C) Hadong2, AO01; (D) Yongin, BB; (E) SN PK, O02O02; and (F) extraction negative control. Colors of the peaks: A, green; T, red; G, blue; C, black.
The ABO blood types of the mummies were successfully obtained based on the SBE reaction results (SN1-2: BO02; Sapgyo: O01O02; Hadong2: AO01; Yongin: BB; SN PK: O02O02) (Figure 3). The ABO results were reproducible in repeated PCR amplifications, a fact that served to confirm their respective authenticities. Whereas phenotype A was found in Hadong2, B in Yongin and SN1-2, and O in Sapgyo and SN PK, no AA or AB phenotype was evident in this study. Our SNP genotype data on the ABO, ABCC11, EDAR and FGFR2 genes is summarized in Table 3.
| No. | Samples | ABO | ABCC11 (rs17822931) | EDAR (rs3827760) | FGFR2 (rs4752566) | ||||
|---|---|---|---|---|---|---|---|---|---|
| nt261 (exon 6) | nt771 (exon 7) | nt796 (exon 7) | nt802 (exon 7) | nt803 (exon 7) | |||||
| 1 | SN1-2 | G/del | C/T | C/A | G | G/C | A | C | T |
| 2 | Sapgyo | del | C/T | C | G | G | A | ND | T |
| 3 | Hadong2 | G/del | C | C | G | G | A | C | T |
| 4 | Yongin | G | C | A | G | C | A | C | T |
| 5 | SN PK | del | T | C | G | G | A | C | T |
| Alleles | G/del | C/T | C/A | G/A | G/C | G/A | T/C | G/T | |
ND, not determined.
Previous scientific analyses have served to demonstrate that mummy tissues and organs are ideal specimens for researchers interested in the health and disease statuses of ancient human populations (Kahila Bar-Gal et al., 2012; Santiago-Rodriguez et al., 2015, 2016). As even 100-year-old soft-tissue samples are still very rare in South Korea, the mummified specimens from Joseon tombs have therefore proved to be invaluable resources in efforts to expand our paleopathological knowledge base. Therefore, under the appropriate permits of the Institutional Research Board, collaborative investigation among medical scientists, archaeologists, and anthropologists has been carried out on Joseon mummies by CT-image-guided autopsy over the past 10 years (Hershkovitz et al., 2014; Kim et al., 2014, 2017). An impressive number of autopsy-derived samples, moreover, have been catalogued for use in more comprehensive scientific studies (Kim et al., 2017).
Meanwhile, aDNA analysis also has been established as an effective research tool for ancient-human-population genetics (Willerslev and Cooper, 2005; Oh et al., 2013). Naturally enough, among various archaeologically obtained specimens, mummy tissues have been regarded as invaluable to paleogenetics, owing specifically to their excellent preservation status. In South Korea, there have been several aDNA studies on mummified human specimens unearthed from Joseon tombs. Briefly, the mitochondrial DNA (mtDNA), Y-chromosomal short tandem repeat (Y-STR), and ABO genes of two Joseon mummies were analyzed by Kim et al. (2011). Oh et al. (2013) showed that aDNA inside mummified brain tissue was less fragmented than that in femurs, thus confirming the brain’s status as the preferred sample source for future genetic analysis of Korean mummies. aDNA analysis has also been applied to the autosomal STR of a Joseon mummy, demonstrating that genotyping can be an indispensable tool in archaeological science (Oh et al., 2015a). The above-noted studies have validated the scientific potential of Korean mummy specimens as subjects of aDNA analysis. However, paleogenetic studies on mummies from South Korea remain unsatisfactory, both qualitatively and quantitatively.
Previous SNP analyses on ancient specimensSNP genotyping analysis allows researchers to study specific alleles in an individual representative of our ancestors. For instance, in an investigation of the genetic traits of the 5300-year-old Copper Age Tyrolean ‘Iceman,’ SNP analyses revealed variants of clinical or physiological relevance in specific genomic regions (Keller et al., 2012). Those variants proved that the Iceman was in all probability lactose intolerant and that he might have had a genetic predisposition to increased risk of atherosclerosis, coronary heart disease, ischemic stroke, and sudden cardiac death. The SNPs in the genes identified as being associated with iris color proved that the Iceman had brown eyes. Furthermore, other variants in the Iceman’s genes showed the characteristics of a blood group O Rh-positive carrier (Keller et al., 2012).
The present study focused mainly on the SNPs in the ABCC11, EDAR, FGFR2, and ABO genes. Whereas there are very few studies on SNPs in the EDAR and FGFR2 genes of ancient people, SNP genotypes in the ABCC11 and ABO genes have already been the subject of previously reported aDNA research. In brief, the allele frequencies of the ABCC11 locus were studied for insight into the ancient populations of the northern Japanese island of Hokkaido, SNP genotyping having been successfully performed for the Okhotsk (originating from eastern Siberia) and Jomon/Epi-Jomon (a major ancestor of the Ainu) peoples. The frequencies of the wet-type allele were higher in the Jomon/Epi-Jomon people, whereas the dry-type allele was much more frequently seen in the Okhotsk people (Sato et al., 2009).
Some studies also have been completed on ABO genotyping analysis. Lalueza-Fox et al. (2008) found a human-specific diagnostic deletion for blood group O (O01 haplo-type) in two Neanderthal skeletons unearthed at the El Sidron site. Ambers et al. (2016) introduced a degenerate oligonucleotide-primed polymerase chain reaction (DOP-PCR)-based method for ABO genotyping of degraded and low-copy templates. As for ancient human samples obtained from archaeological sites in South Korea, Kim et al. (2011) utilized a multiplex SBE method to determine the blood types of two pre-modern Korean mummies (AO01 and AB). Although these studies have successfully validated the scientific potential of mummies as the subjects of scientific studies, most of them entailed the retrieval and analysis of aDNA from only one or two samples. The current study is therefore especially meaningful in that it demonstrated the applicability of using ancient samples for SNP genotyping of the ABCC11, EDAR, FGFR2, and ABO genes based on a larger sample of mummy specimens.
SNP analyses on Joseon mummiesRecent evidence suggests that the risk of axillary osmidrosis can be determined by SNP in the ABCC11 gene. The 538G allele in the ABCC11 gene is associated with the high-secretory phenotypes of the apocrine gland, increased risk of axillary osmidrosis, and the wet type of earwax, with the opposite being observed for the 538A allele (Toyoda et al., 2017). In fact, ABCC11 538G→A diminishes the ABCC11 protein level in the axillary apocrine glands of subjects with the AA homozygote. The higher allele frequency of ABCC11 538G in European populations than in East Asian ones explains why axillary osmidrosis is more common in the former (Toyoda et al., 2017). In a previous report, ABCC11 538A was found in 100% of Koreans examined (Ishikawa et al., 2012). The presence of an SNP in the ABCC11 genes of the present Joseon mummies, namely the 538A allele, thus made sense to us as indicative of the dry type of earwax and a low risk of axillary osmidrosis, both traits being typical among Korean people.
Meanwhile, the SNPs in EDAR and FGFR2 are known to determine hair thickness (Fujimoto et al., 2008a, b, 2009). In an animal experiment, Mou et al. (2008) proved that elevation of EDAR activity decreases the number of kinks in the hair fiber while increasing its diameter. Therefore, the variant EDAR gene can be considered to be implicated in the straighter and coarser hair commonly seen among Chinese, Koreans, Japanese, and Amerindian peoples. This conclusion was further supported by the data of Bryk et al. (2008) and Fujimoto et al. (2008b), which indicated that EDAR gene substitution is rarely found in African, Central/South Asian, European, and Middle Eastern peoples. The thick, straight hair of East Asian peoples has also been revealed to be linked to the allele variants in the FGFR2 gene, which is, like the EDAR gene, mainly observed among such populations (Fujimoto et al., 2009). In fact, the alleles rs3827760-C (EDAR) and rs4752566-T (FGFR2) were found in 78.6% and 89.5% of the Japanese population, respectively (Fujimoto et al., 2008b, 2009). In our current study on the SNPs in the EDAR and FGFR2 genes of Joseon mummies, almost the same results were obtained from each specimen. Briefly, in the analysis of the SNP in the EDAR gene, every mummy had the rs3827760-C variant allele; and as for the SNP in the FGFR2 gene, only variant allele rs4752566-T was found in all the specimens. Based on these SNP analysis results, every Korean mummy in this study shared the physical trait of thick, straight hair that is commonly seen among most East Asian people.
Finally, the ABO genotypes of the Joseon mummies were successfully obtained based on the SBE reaction results: phenotypes A, B, and O were found in one (Hadong2), two (Yongin and SN1-2), and two (Sapygo and SN PK) mummy samples, respectively. In previous research conducted with the participation of 111842 Korean blood donors, the most common blood type was phenotype A (36.4%), followed by O (26.0%), B (25.7%), and AB (11.7%) (Cho et al., 2003). While blood type A scarcer than other blood types among Korean mummies, the small sample size in this study precludes conclusive identification of blood type prevalence among the Joseon people. Despite this limitation, the current data adequately fills the knowledge gap regarding the utility of ABO genotyping in forensic medicine for analysis of ancient samples.
The current study is significant because it demonstrates that SNP genotyping analyses on the ABCC11, EDAR, FGFR2, and ABO genes can be effectively applicable to ancient specimens from several-hundred-year-old Joseon mummies from South Korea. In fact, due to the rapid rise of DNA typing, SNP analysis nowadays is being increasingly used for genetic analysis of human remains found at archaeological sites. In this study, we also proved that SNP genotyping can help to uncover the physical traits of long-dead Korean mummified individuals. The latest analytical tools, such as microarrays, are still prohibitively expensive for routine analysis of archaeological samples (Kayser, 2015). In this light, our study can be particularly helpful for specific tasks such as tracing the applicability of SNPs among different sets of ancient remains worldwide by more conventional SNaPshot analysis.
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Ministry of Education (NRF-2017R1D1A1B03030127) and by a Seoul National University Hospital research grant (0420170490).